Gut Microbiome—How Does Two-Month Consumption of Fiber-Enriched Rolls Change Microbiome in Patients Suffering from MASLD?

 ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
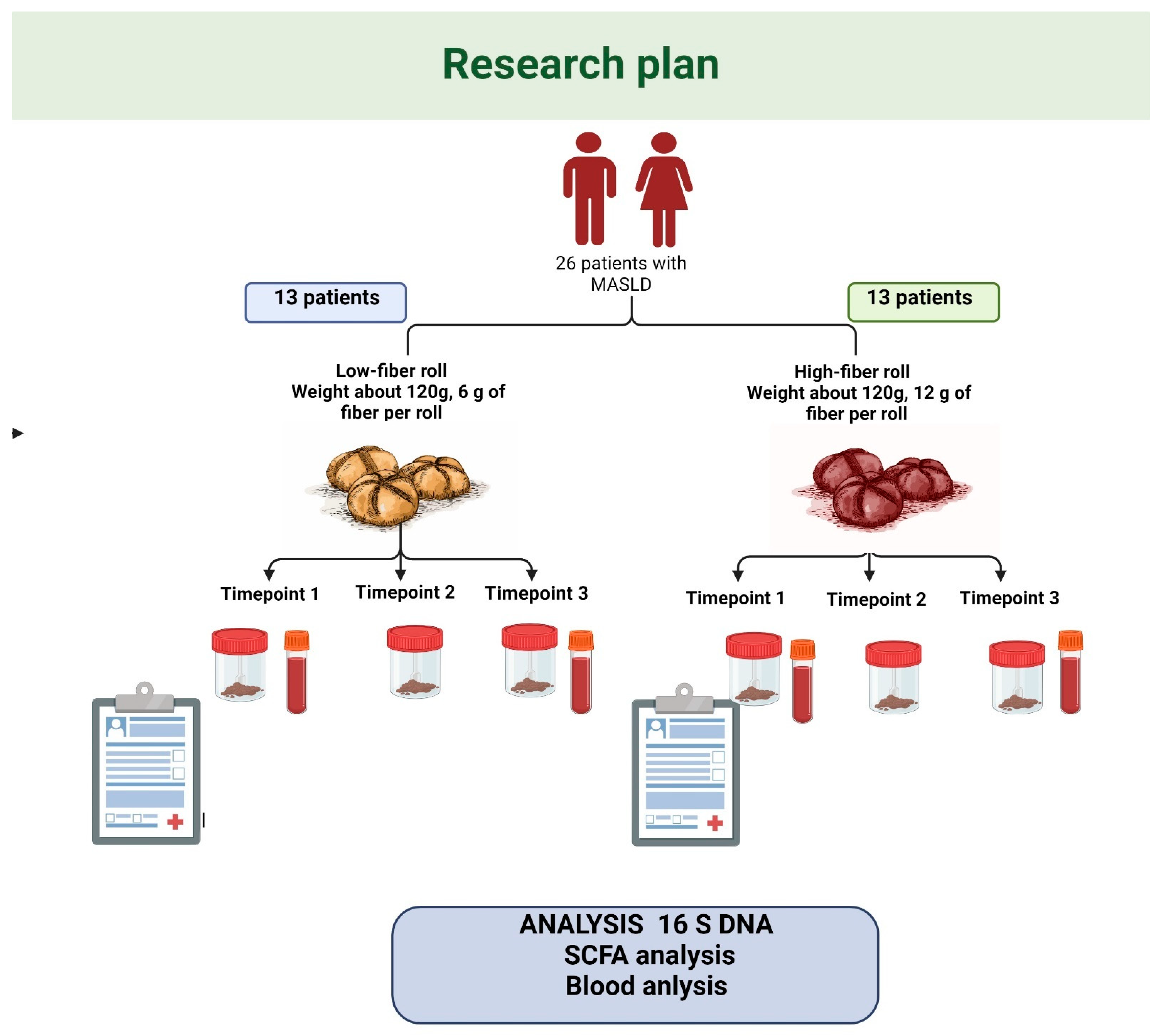
2.1. Study Design
2.2. Dietary Guidelines
2.3. Fecal Microbiota Sequenced Using 16S on Nanopore
2.4. Short-Chain Fatty Acids (SCFA) Analysis
2.5. Bioinformatic and Statistical Analysis
2.5.1. Taxonomic Assignment of 16S Reads
2.5.2. Statistical Analysis
3. Results
3.1. Patients’ Characteristics
3.2. Gut Microbiota-Derived Metabolites (SCFAs)
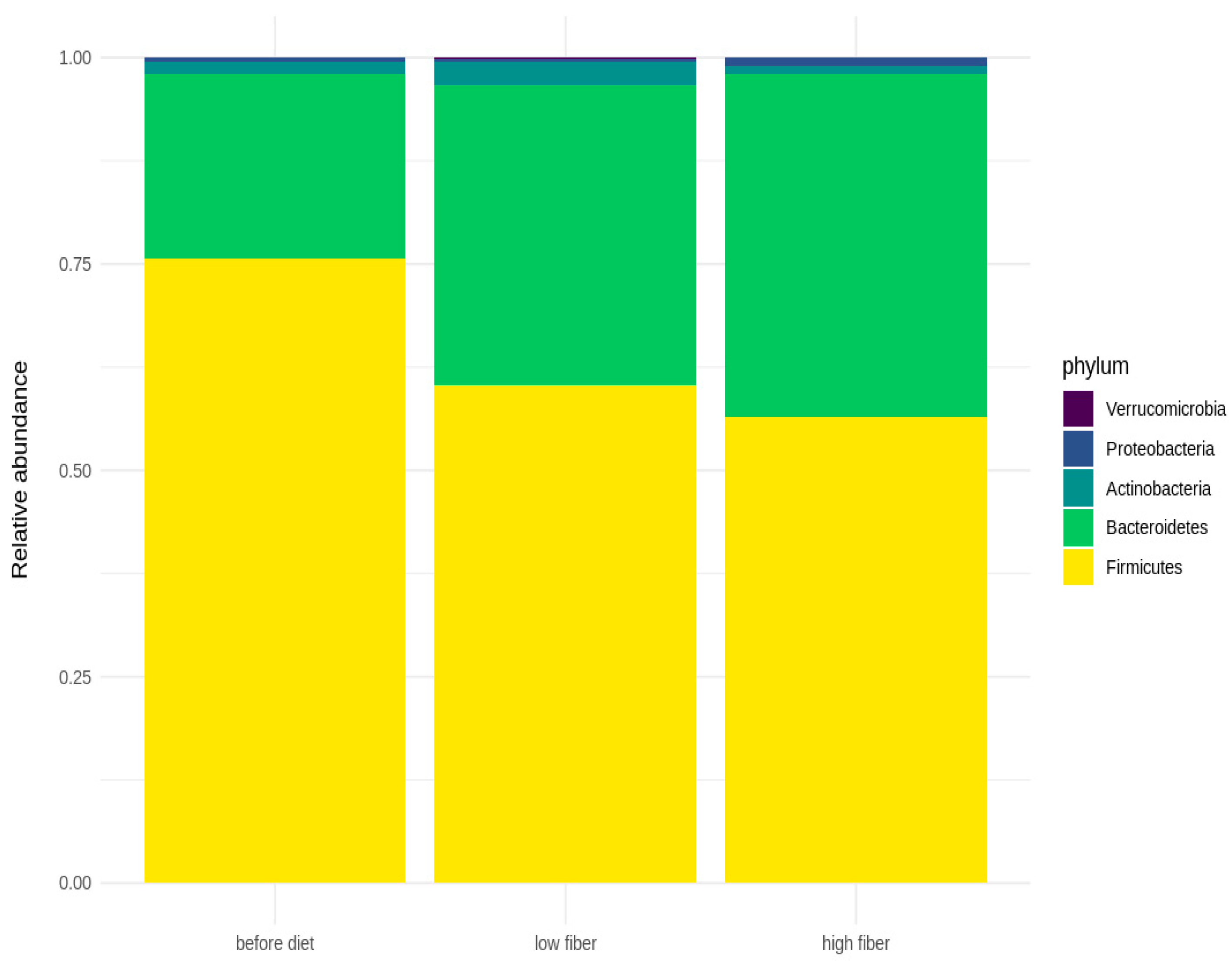
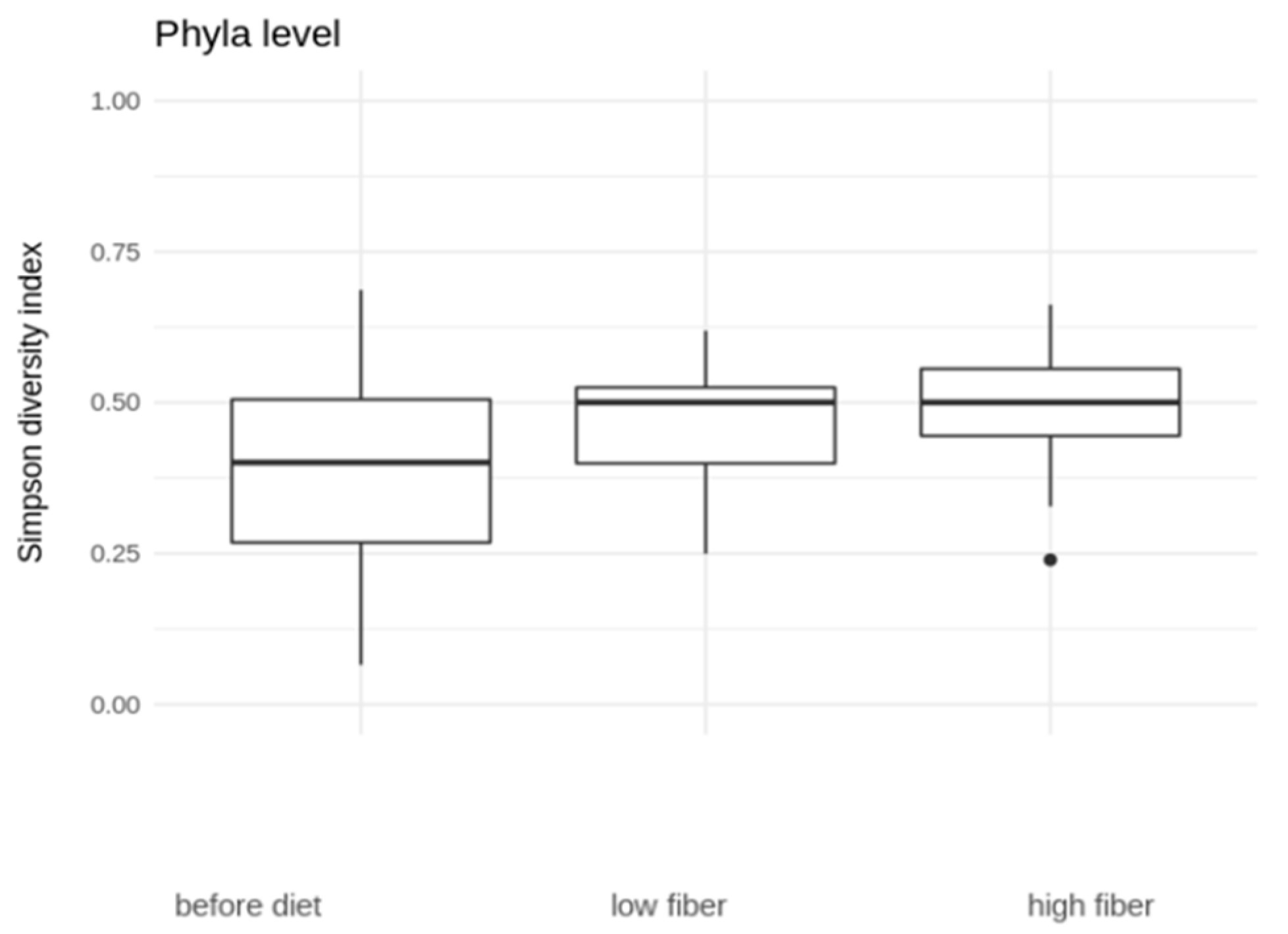
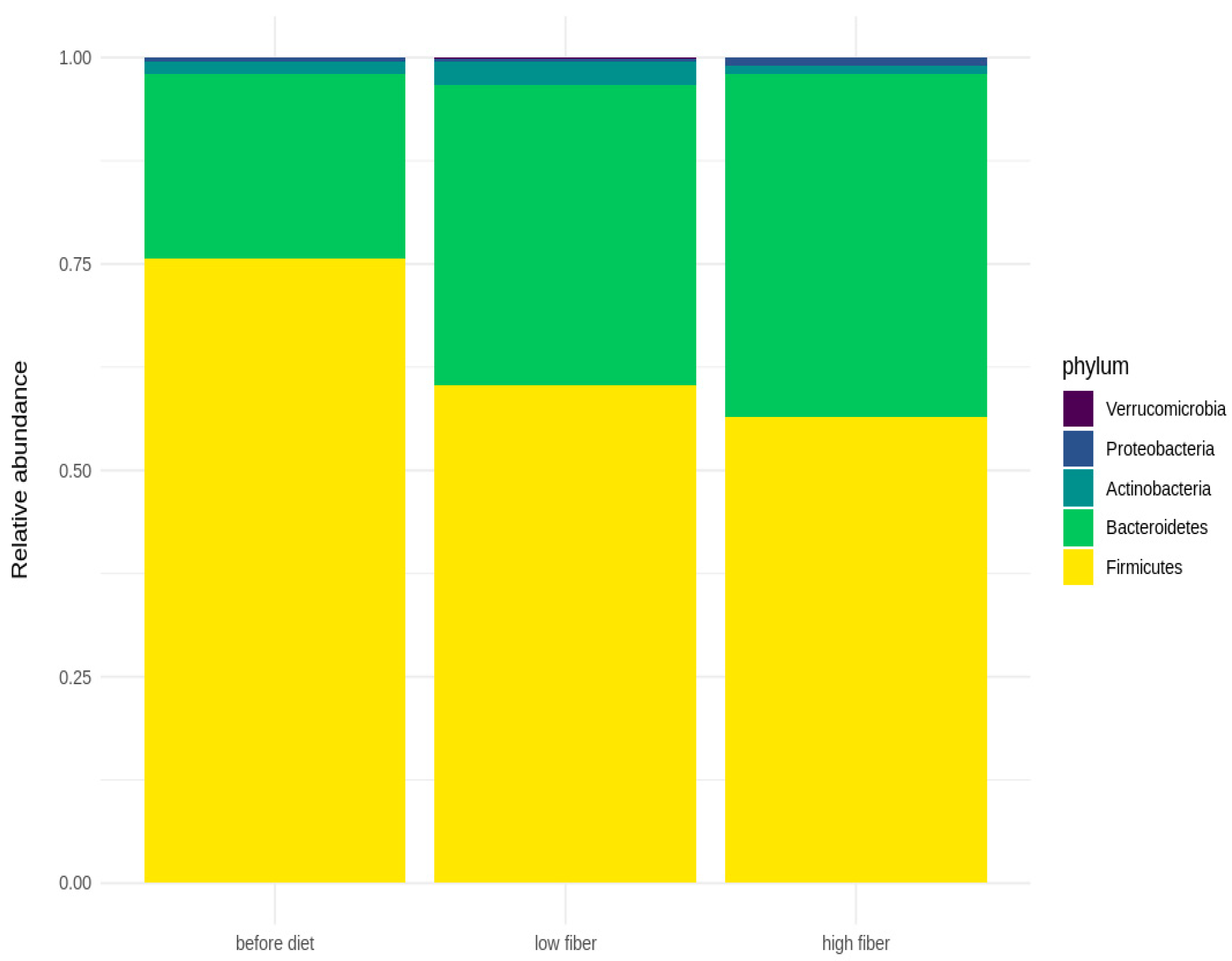
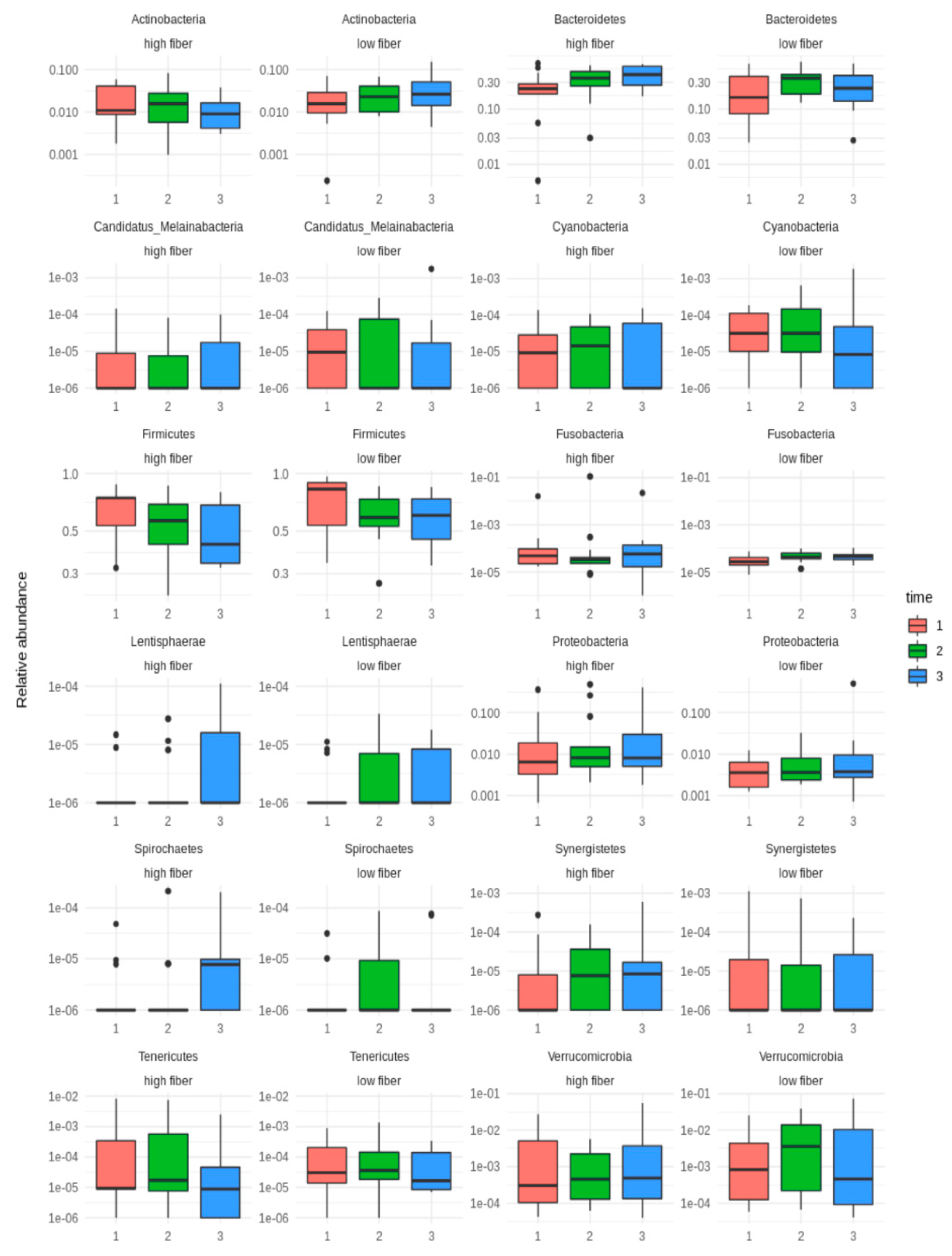
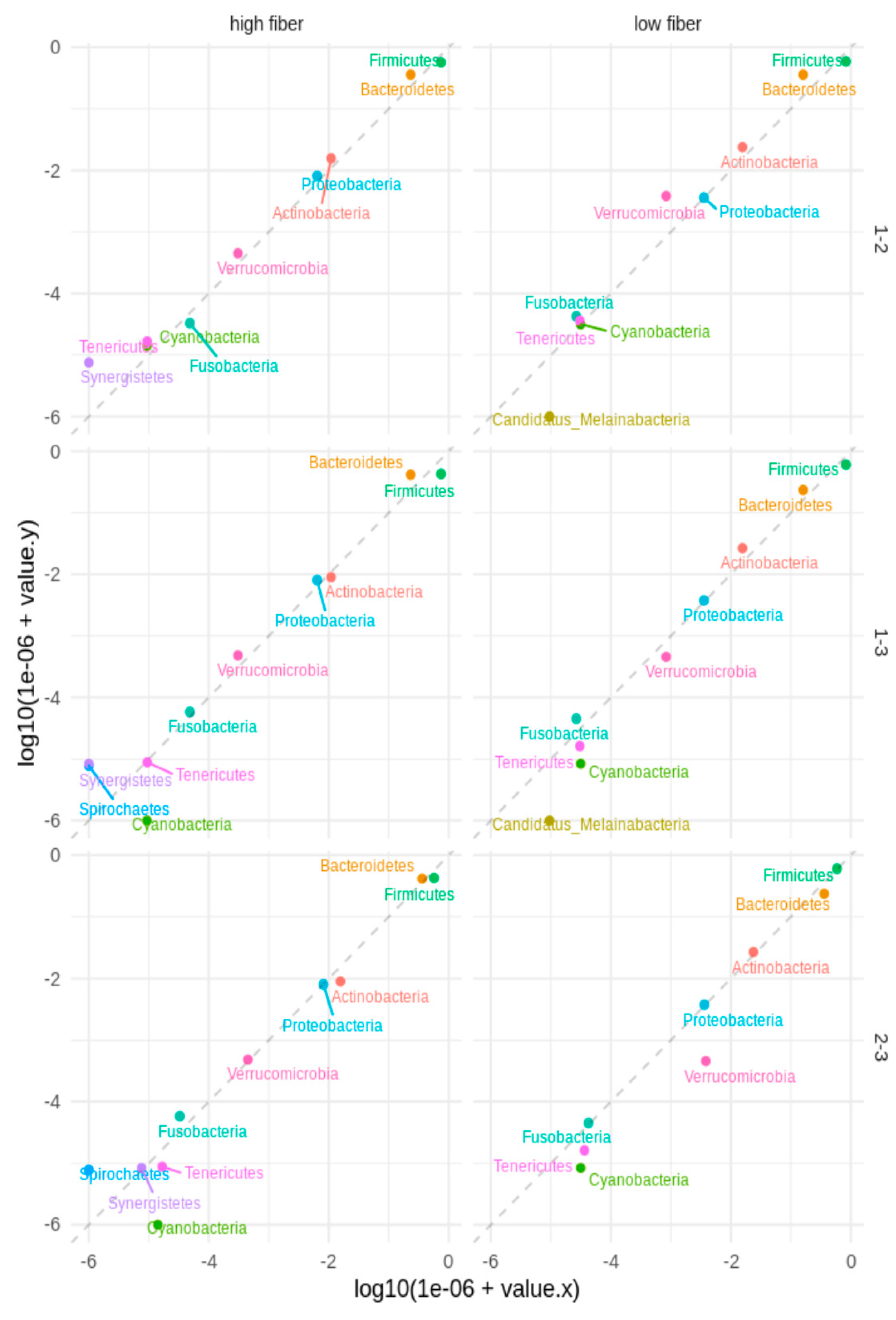
3.3. Microbial Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krag, A.; Buti, M.; Lazarus, J.V.; Allen, A.M.; Bowman, J.; Burra, P.; Donnini, G.; Duseja, A.; El-Sayed, M.H.; Gastaldelli, A.; et al. Uniting to defeat steatotic liver disease: A global mission to promote healthy livers and healthy lives. J. Hepatol. 2023, 79, 1076–1078. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Golabi, P.; Paik, J.M.; Henry, A.; Van Dongen, C.; Henry, L. The global epidemiology of nonalcoholic fatty liver disease (MASLD) and nonalcoholic steatohepatitis (NASH): A systematic review. Hepatology 2023, 77, 1335–1347. [Google Scholar] [CrossRef]
- Mitra, S.; De, A.; Chowdhury, A. Epidemiology of non-alcoholic and alcoholic fatty liver diseases. Transl. Gastroenterol. Hepatol. 2020, 5, 16. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Yaskolka Meir, A.; Rinott, E.; Tsaban, G.; Zelicha, H.; Kaplan, A.; Rosen, P.; Shelef, I.; Youngster, I.; Shalev, A.; Blüher, M.; et al. Effect of green-Mediterranean diet on intrahepatic fat: The DIRECT PLUS randomised controlled trial. Gut 2021, 70, 2085–2095. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Y.; Gong, C.; Wang, J.; Liu, B.; Shi, L.; Duan, J. Prevalence of malnutrition and analysis of related factors in elderly patients with COVID-19 in Wuhan, China. Eur. J Clin. Nutr. 2020, 74, 871–875. [Google Scholar] [CrossRef] [PubMed]
- Meroni, M.; Longo, M.; Rustichelli, A.; Dongiovanni, P. Nutrition and Genetics in NAFLD: The Perfect Binomium. Int. J Mol. Sci. 2020, 21, 2986. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Boccuto, L.; Federico, A.; Dallio, M.; Loguercio, C.; Di Renzo, L.; De Lorenzo, A. Diet and Non-Alcoholic Fatty Liver Disease: The Mediterranean Way. Int. J. Environ. Res. Public Health 2019, 16, 3011. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Nicolucci, A.C.; Virtanen, H.; Schick, A.; Meddings, J.; Reimer, R.A.; Huang, C. Effect of Prebiotic on Microbiota, Intestinal Permeability, and Glycemic Control in Children With Type 1 Diabetes. J. Clin. Endocrinol. Metab. 2019, 104, 4427–4440. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.; Ojo, O.O.; Zand, N.; Wang, X. The Effect of Dietary Fibre on Gut Microbiota, Lipid Profile, and Inflammatory Markers in Patients with Type 2 Diabetes: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2021, 13, 1805. [Google Scholar] [CrossRef] [PubMed]
- Stachowska, E.; Portincasa, P.; Jamioł-Milc, D.; Maciejewska-Markiewicz, D.; Skonieczna-Żydecka, K. The Relationship between Prebiotic Supplementation and Anthropometric and Biochemical Parameters in Patients with NAFLD-A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2020, 12, 3460. [Google Scholar] [CrossRef] [PubMed]
- Hagströmer, M.; Oja, P.; Sjöström, M. The International Physical Activity Questionnaire (IPAQ): A study of concurrent and construct validity. Public Health Nutr. 2006, 9, 755–762. [Google Scholar] [CrossRef]
- Bach-Faig, A.; Berry, E.M.; Lairon, D.; Reguant, J.; Trichopoulou, A.; Dernini, S.; Medina, F.X.; Battino, M.; Belahsen, R.; Miranda, G.; et al. Mediterranean diet pyramid today. Science and cultural up-dates. Public Health Nutr. 2011, 14, 2274–2284. [Google Scholar] [CrossRef] [PubMed]
- Karlas, T.; Petroff, D.; Garnov, N.; Böhm, S.; Tenckhoff, H.; Wittekind, C.; Wiese, M.; Schiefke, I.; Linder, N.; Schaudinn, A.; et al. Non-invasive assessment of hepatic steatosis in patients with NAFLD using con-trolled attenuation parameter and 1H-MR spectroscopy. PLoS ONE 2014, 9, e91987. [Google Scholar] [CrossRef]
- Aparicio-Ugarriza, R.; Cuenca-García, M.; Gonzalez-Gross, M.; Julián, C.; Bel-Serrat, S.; Moreno, L.A.; Breidenassel, C.; Kersting, M.; Arouca, A.B.; Michels, N.; et al. Relative validation of the adapted Mediterranean Diet Score for Adolescents by comparison with nutritional biomarkers and nutrient and food in-takes: The Healthy Lifestyle in Europe by Nutrition in Adolescence (HELENA) study. Public Health Nutr. 2019, 22, 2381–2397. [Google Scholar] [CrossRef]
- Safari, Z.; Gérard, P. The links between the gut microbiome and non-alcoholic fatty liver disease (NAFLD). Cell Mol. Life Sci. 2019, 76, 1541–1558. [Google Scholar] [CrossRef]
- Jamioł-Milc, D.; Gudan, A.; Kaźmierczak-Siedlecka, K.; Hołowko-Ziółek, J.; Maciejewska-Markiewicz, D.; Janda-Milczarek, K.; Stachowska, E. Nutritional Support for Liver Diseases. Nutrients 2023, 15, 3640. [Google Scholar] [CrossRef]
- Semmler, G.; Datz, C.; Reiberger, T.; Trauner, M. Diet and exercise in NAFLD/NASH: Beyond the obvious. Liver Int. 2021, 41, 2249–2268. [Google Scholar] [CrossRef]
- Haigh, L.; Kirk, C.; El Gendy, K.; Gallacher, J.; Errington, L.; Mathers, J.C.; Anstee, Q.M. The effectiveness and acceptability of Mediterranean diet and calorie restriction in non-alcoholic fatty liver dis-ease (NAFLD): A systematic review and meta-analysis. Clin. Nutr. 2022, 41, 1913–1931. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Montes de Oca, A.; Julián, M.T.; Ramos, A.; Puig-Domingo, M.; Alonso, N. Microbiota, Fiber, and NAFLD: Is There Any Connection? Nutrients 2020, 12, 3100. [Google Scholar] [CrossRef]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Usuda, H.; Okamoto, T.; Wada, K. Leaky Gut: Effect of Dietary Fiber and Fats on Microbiome and Intestinal Barrier. Int. J. Mol. Sci. 2021, 22, 7613. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Siedlecka, K.; Skonieczna-Żydecka, K.; Palma, J.; Sobocki, B.; Świerblewski, M.; Siedlecka-Kroplewska, K.; Kalinowski, L.; Połom, K. Microbiota-derived metabolites in colorectal cancer patients in preoperative period. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 1443–1449. [Google Scholar] [PubMed]
- Deng, M.; Qu, F.; Chen, L.; Liu, C.; Zhang, M.; Ren, F.; Guo, H.; Zhang, H.; Ge, S.; Wu, C.; et al. SCFAs alleviated steatosis and inflammation in mice with NASH induced by MCD. J. Endocrinol. 2020, 245, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Yang, X.; Wu, Q.; Gong, Y.; Pang, N.; Ge, X.; Nagaratnam, N.; Jiang, P.; Zhou, M.; Hu, T.; et al. Butyrate Attenuates Hepatic Steatosis Induced by a High-Fat and Fiber-Deficient Diet via the Hepatic GPR41/43-CaMKII/HDAC1-CREB Pathway. Mol. Nutr. Food Res. 2023, 67, e2200597. [Google Scholar] [CrossRef] [PubMed]
- Rau, M.; Rehman, A.; Dittrich, M.; Groen, A.K.; Hermanns, H.M.; Seyfried, F.; Beyersdorf, N.; Dandekar, T.; Rosenstiel, P.; Geier, A. Fecal SCFAs and SCFA-producing bacteria in gut microbiome of human NAFLD as a putative link to systemic T-cell activation and advanced disease. United Eur. Gastroenterol. J. 2018, 6, 1496–1507. [Google Scholar] [CrossRef]
- Wu, M.Y.; Fan, J.G. Gut microbiome and nonalcoholic fatty liver disease. Hepatobiliary Pancreat. Dis. Int. 2023, 22, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Vitetta, L. Gut Microbiota Metabolites in NAFLD Pathogenesis and Therapeutic Implications. Int. J. Mol. Sci. 2020, 21, 5214. [Google Scholar] [CrossRef]
- Ji, Y.; Yin, Y.; Li, Z.; Zhang, W. Gut Microbiota-Derived Components and Metabolites in the Progression of Non-Alcoholic Fatty Liver Disease (NAFLD). Nutrients 2019, 11, 1712. [Google Scholar] [CrossRef]
- Hrncir, T.; Hrncirova, L.; Kverka, M.; Hromadka, R.; Machova, V.; Trckova, E.; Kostovcikova, K.; Kral-ickova, P.; Krejsek, J.; Tlaskalova-Hogenova, H. Gut Microbiota and NAFLD: Pathogenetic Mechanisms, Microbiota Signatures, and Therapeutic Interventions. Microorganisms 2021, 9, 957. [Google Scholar] [CrossRef]
- Ohtani, N.; Hara, E. Gut-liver axis-mediated mechanism of liver cancer: A special focus on the role of gut microbiota. Cancer Sci. 2021, 112, 4433–4443. [Google Scholar] [CrossRef] [PubMed]
- Beisner, J.; Filipe Rosa, L.; Kaden-Volynets, V.; Stolzer, I.; Günther, C.; Bischoff, S.C. Prebiotic Inulin and Sodium Butyrate Attenuate Obesity-Induced Intestinal Barrier Dysfunction by Induction of Antimicrobial Peptides. Front. Immunol. 2021, 12, 678360. [Google Scholar] [CrossRef]
- Behrouz, V.; Aryaeian, N.; Zahedi, M.J.; Jazayeri, S. Effects of probiotic and prebiotic supplementation on metabolic parameters, liver aminotransferases, and systemic inflammation in nonalcoholic fatty liver disease: A randomized clinical trial. J. Food Sci. 2020, 85, 3611–3617. [Google Scholar] [CrossRef] [PubMed]
- Leung, H.; Long, X.; Ni, Y.; Qian, L.; Nychas, E.; Siliceo, S.L.; Pohl, D.; Hanhineva, K.; Liu, Y.; Xu, A.; et al. Risk assessment with gut microbiome and metabolite markers in NAFLD development. Sci. Transl. Med. 2022, 14, eabk0855. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.; Christodoulatos, G.S.; Karampela, I.; Tsilingiris, D.; Magkos, F.; Stratigou, T.; Kounatidis, D.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Non-Alcoholic Fatty Liver Disease: Current Evidence and Perspectives. Biomolecules 2021, 12, 56. [Google Scholar] [CrossRef]
- Di Ciaula, A.; Bonfrate, L.; Portincasa, P. The role of microbiota in nonalcoholic fatty liver disease. Eur. J Clin. Investig. 2022, 52, e13768. [Google Scholar] [CrossRef]
- Marques, F.Z.; Nelson, E.; Chu, P.Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef]
- Jasirwan, C.O.M.; Muradi, A.; Hasan, I.; Simadibrata, M.; Rinaldi, I. Correlation of gut Firmicutes/Bacteroidetes ratio with fibrosis and steatosis stratified by body mass index in patients with non-alcoholic fatty liver disease. Biosci. Microbiota Food Health 2021, 40, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, Y.; Xie, H.; Bai, H.; Lin, G.; Dong, Y.; Shi, D.; Wang, J.; Zhang, Q.; Zhang, Y.; et al. Impact of a low-carbohydrate and high-fiber diet on nonalcoholic fatty liver disease. Asia Pac. J. Clin. Nutr. 2020, 29, 483–490. [Google Scholar] [PubMed]
- Zhu, Y.; Yang, H.; Zhang, Y.; Rao, S.; Mo, Y.; Zhang, H.; Yang, W. Dietary fiber intake and non-alcoholic fatty liver disease: The mediating role of obesity. Front. Public Health 2022, 10, 1038435. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Values (Min.–Max.) |
|---|---|
| BMI | 29.1 (22.2–35.7) |
| Body_weight | 87.4 (60.3–115.6) |
| Body fat | 29.5 (17.1–43.8) |
| Muscle_mass | 54.6 (39.5–76.7) |
| Fibroscan_CAP | 305.5 (242–400) |
| Fibroscan_elast | 5.65 (3.9–9.4) |
| ALT | 29 (11–136) |
| AST | 22 (11–52) |
| GGTP | 27.5 (12–70) |
| Total_Cholesterol | 205.6 (110–394.4) |
| LDL | 137 (43.5–282.2) |
| HDL | 47.15 (25–71.3) |
| Intervention with 12 g (LFIB) T1 vs. T3 | Intervention with 24 g (HFIB) T1 vs. T3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Parameter Intervention | Median | IQR | Median | IQR | p | Median | IQR | Median | IQR | p |
| Fasting glucose [mg/dL] | 93 | 16.2 | 91 | 29.5 | 0.72 | 94.6 | 15.3 | 96.9 | 10.2 | 0.94 |
| Total cholesterol [mg/dL] | 191.4 | 53.2 | 179.9 | 27.1 | 0.18 | 221 | 59.6 | 197.3 | 47.7 | 0.01 |
| HDL [mg/dL] | 44 | 12.1 | 43.1 | 15.9 | 0.58 | 49.1 | 6 | 48.7 | 6.5 | 0.17 |
| LDL [mg/dL] | 125 | 54.5 | 111.9 | 42.1 | 0.21 | 148.9 | 38.9 | 127.7 | 44 | 0.05 |
| TG [mg/dL] | 153.3 | 94 | 129.3 | 92 | 0.11 | 168.2 | 154.2 | 162.3 | 111 | 0.28 |
| ALT [U/L] | 38 | 17 | 38 | 18 | 0.89 | 43 | 20 | 32 | 12 | 0.04 |
| AST [U/L] | 28 | 12 | 30 | 11 | 0.66 | 27 | 8 | 23 | 6 | 0.02 |
| GGTP [U/L] | 33 | 10 | 35 | 16 | 0.23 | 28 | 12 | 24 | 14 | 0.12 |
| Fasting insulin [uU/mL] | 19.1 | 20.3 | 16.1 | 13.9 | 0.66 | 36.8 | 87.2 | 37.6 | 31 | 0.18 |
| Age [years] | 47.5 | 12.3 | - | - | - | 47.5 | 14.5 | - | - | - |
| BMI [kg/m2] | 29.1 | 3.8 | 28.6 | 5.2 | 0.04 | 28.5 | 10.4 | 27.3 | 9.5 | 0.61 |
| Intervention with 12 g (LFIB) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| SCFA mol% | C 2:0 A,B | C 3:0 | C 4:0 n B,C | ||||||
| Visit | T1 | T2 | T3 | T1 | T2 | T3 | T1 | T2 | T3 |
| Median | 64.93 | 68.53 | 62.51 | 19.66 | 17.11 | 18.46 | 14.38 | 12.65 | 18.9 |
| IQR | 14.91 | 10.61 | 14.01 | 4.46 | 3.19 | 3.35 | 7.5 | 9.64 | 7.52 |
| Intervention with 24 g (HFIB) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| SCFA mol% | C 2:0 A,B | C 3:0 | C 4:0 n B*,C | ||||||
| Visit | T1 | T2 | T3 | T1 | T2 | T3 | T1 | T2 | T3 |
| Median | 60.52 | 63.12 | 56.1 | 20.39 | 19.08 | 20.18 | 14.79 | 12.61 | 15.76 |
| IQR | 9.62 | 10.89 | 9.84 | 4.52 | 4.51 | 2.71 | 4.69 | 7.74 | 7.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaźmierczak-Siedlecka, K.; Maciejewska-Markiewicz, D.; Sykulski, M.; Gruszczyńska, A.; Herman-Iżycka, J.; Wyleżoł, M.; Katarzyna Petriczko, K.; Palma, J.; Jakubczyk, K.; Janda-Milczarek, K.; et al. Gut Microbiome—How Does Two-Month Consumption of Fiber-Enriched Rolls Change Microbiome in Patients Suffering from MASLD? Nutrients 2024, 16, 1173. https://doi.org/10.3390/nu16081173
Kaźmierczak-Siedlecka K, Maciejewska-Markiewicz D, Sykulski M, Gruszczyńska A, Herman-Iżycka J, Wyleżoł M, Katarzyna Petriczko K, Palma J, Jakubczyk K, Janda-Milczarek K, et al. Gut Microbiome—How Does Two-Month Consumption of Fiber-Enriched Rolls Change Microbiome in Patients Suffering from MASLD? Nutrients. 2024; 16(8):1173. https://doi.org/10.3390/nu16081173
Chicago/Turabian StyleKaźmierczak-Siedlecka, Karolina, Dominika Maciejewska-Markiewicz, Maciej Sykulski, Agata Gruszczyńska, Julia Herman-Iżycka, Mariusz Wyleżoł, Karolina Katarzyna Petriczko, Joanna Palma, Karolina Jakubczyk, Katarzyna Janda-Milczarek, and et al. 2024. "Gut Microbiome—How Does Two-Month Consumption of Fiber-Enriched Rolls Change Microbiome in Patients Suffering from MASLD?" Nutrients 16, no. 8: 1173. https://doi.org/10.3390/nu16081173
APA StyleKaźmierczak-Siedlecka, K., Maciejewska-Markiewicz, D., Sykulski, M., Gruszczyńska, A., Herman-Iżycka, J., Wyleżoł, M., Katarzyna Petriczko, K., Palma, J., Jakubczyk, K., Janda-Milczarek, K., Skonieczna-Żydecka, K., & Stachowska, E. (2024). Gut Microbiome—How Does Two-Month Consumption of Fiber-Enriched Rolls Change Microbiome in Patients Suffering from MASLD? Nutrients, 16(8), 1173. https://doi.org/10.3390/nu16081173