Effects of Kimchi Intake on the Gut Microbiota and Metabolite Profiles of High-Fat-Induced Obese Rats

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Kimchi
2.2. Animal Study
2.3. Biochemical Markers
2.4. Analysis of Gut Microbiota
2.5. Global Metabolomic Analysis
2.6. Steroid Hormone and Bile Acid Analysis Using UPLC-Q-TOF MS
2.7. Analysis of Short-Chain Fatty Acids (SCFAs) by GC–MS
2.8. Real-Time PCR
2.9. Statistical Analysis
3. Results
3.1. Biological Characteristics
3.2. Gene Expression Levels of Inflammatory Cytokines in the Colon
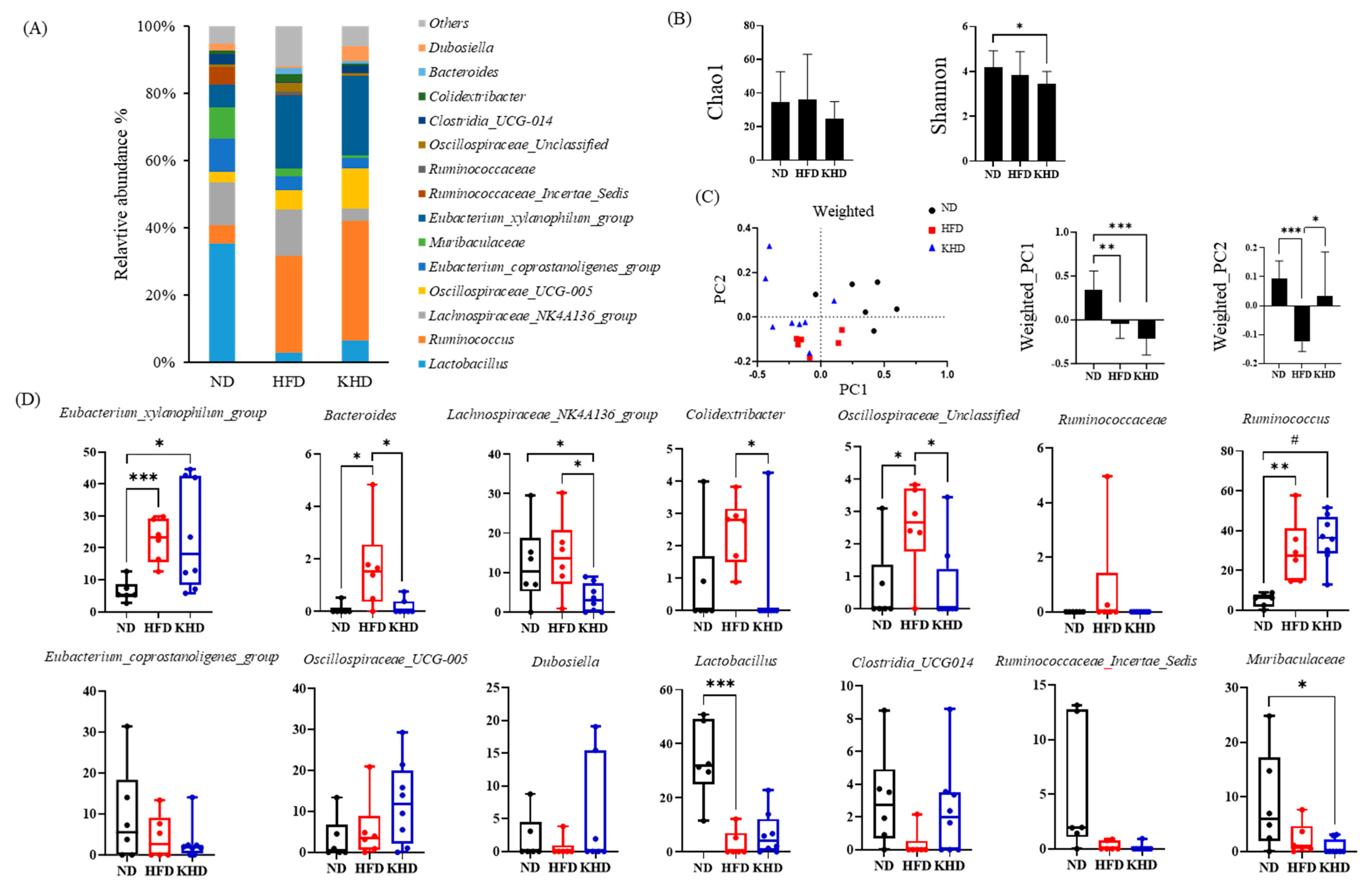
3.3. Gut Microbiota
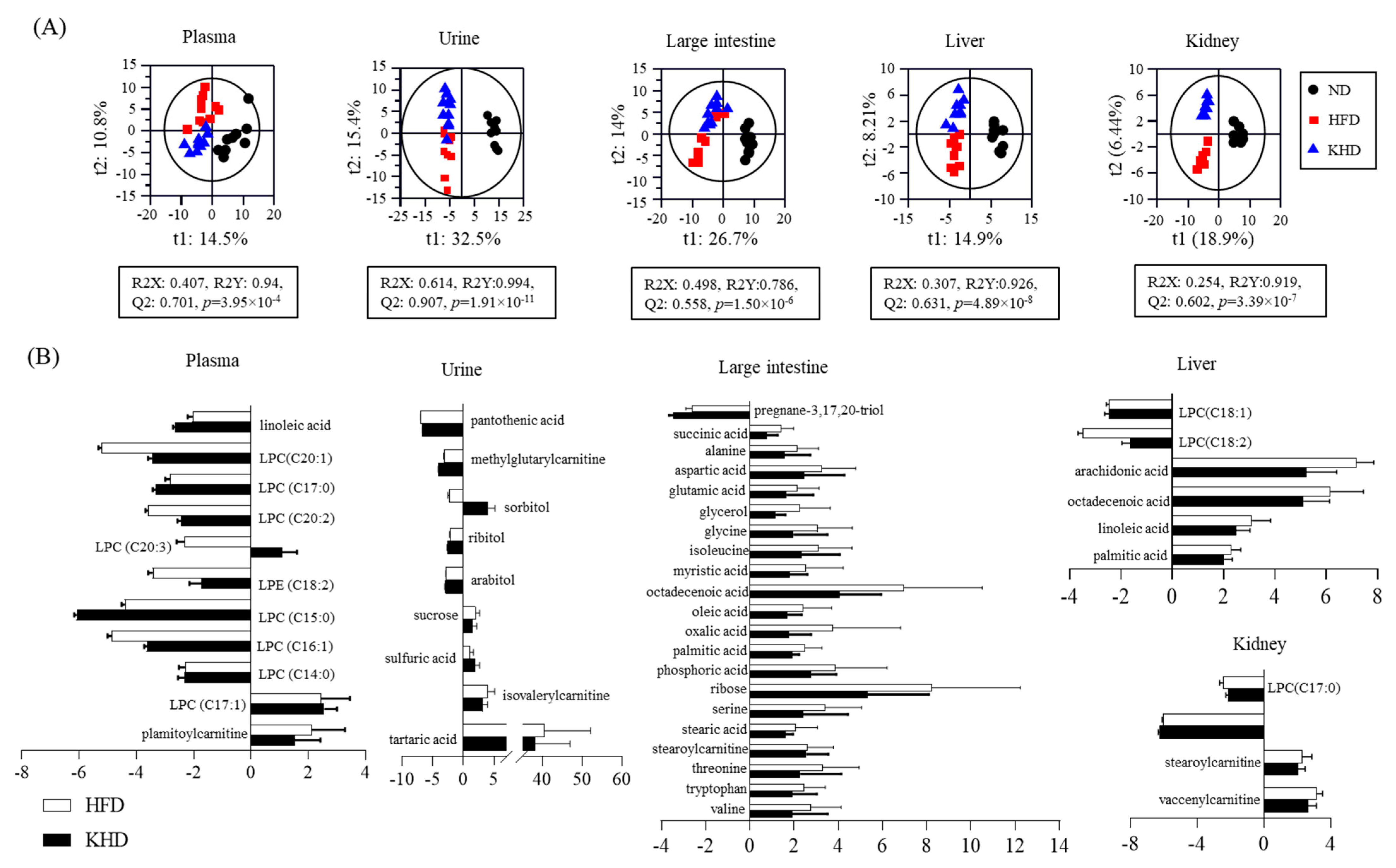
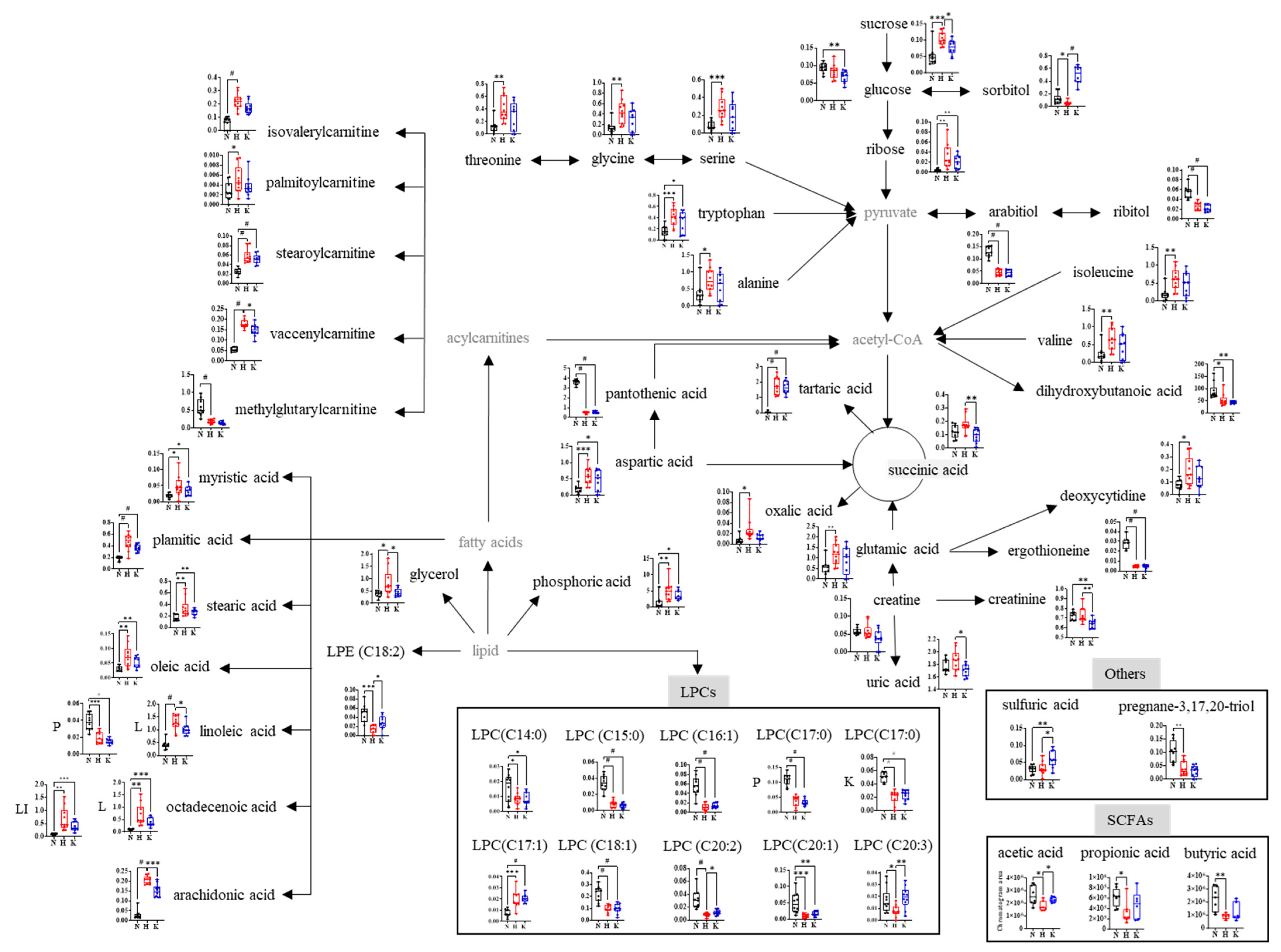
3.4. Global Metabolites and SCFAs
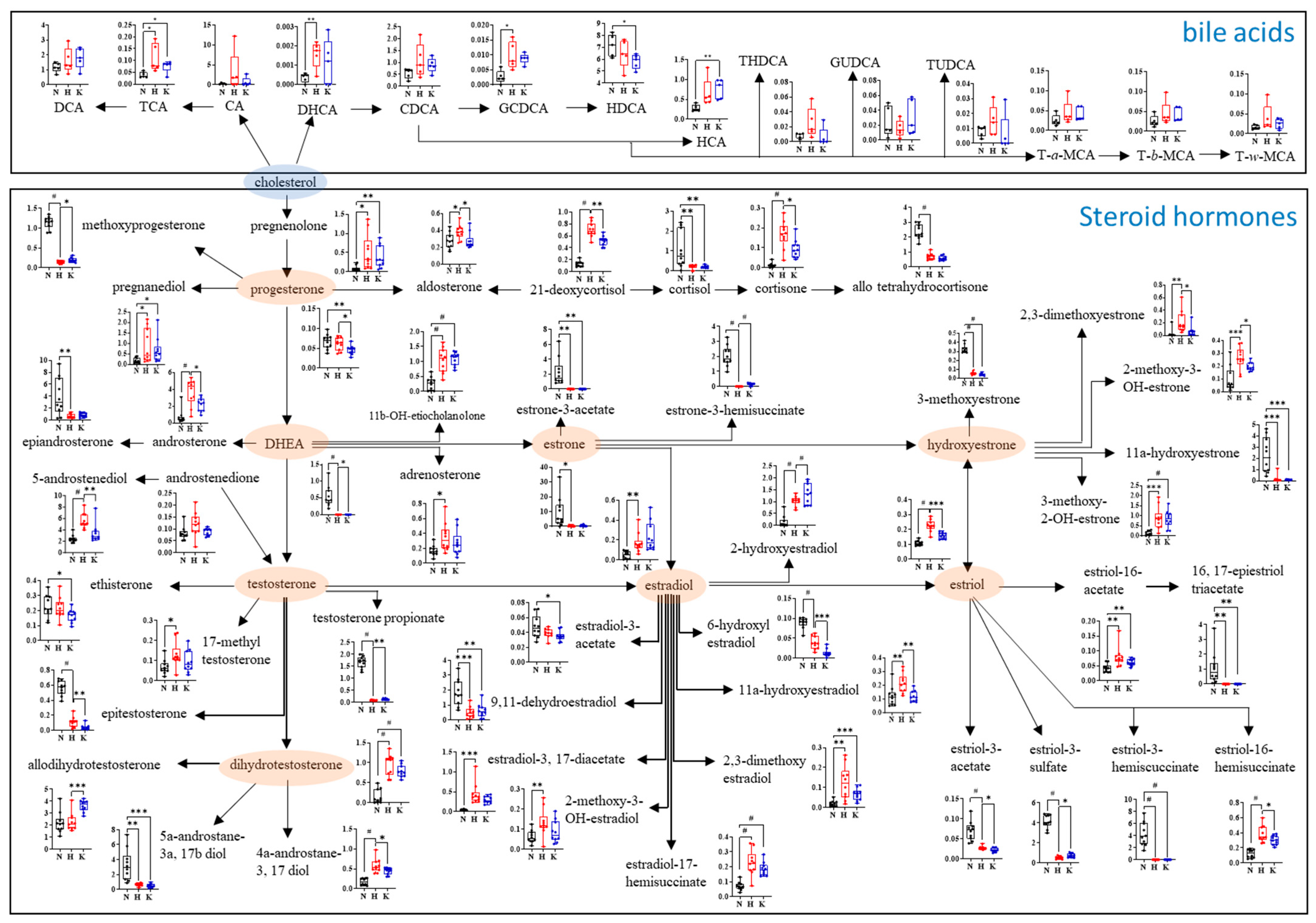
3.5. Steroid Hormone and Bile Acid Profile
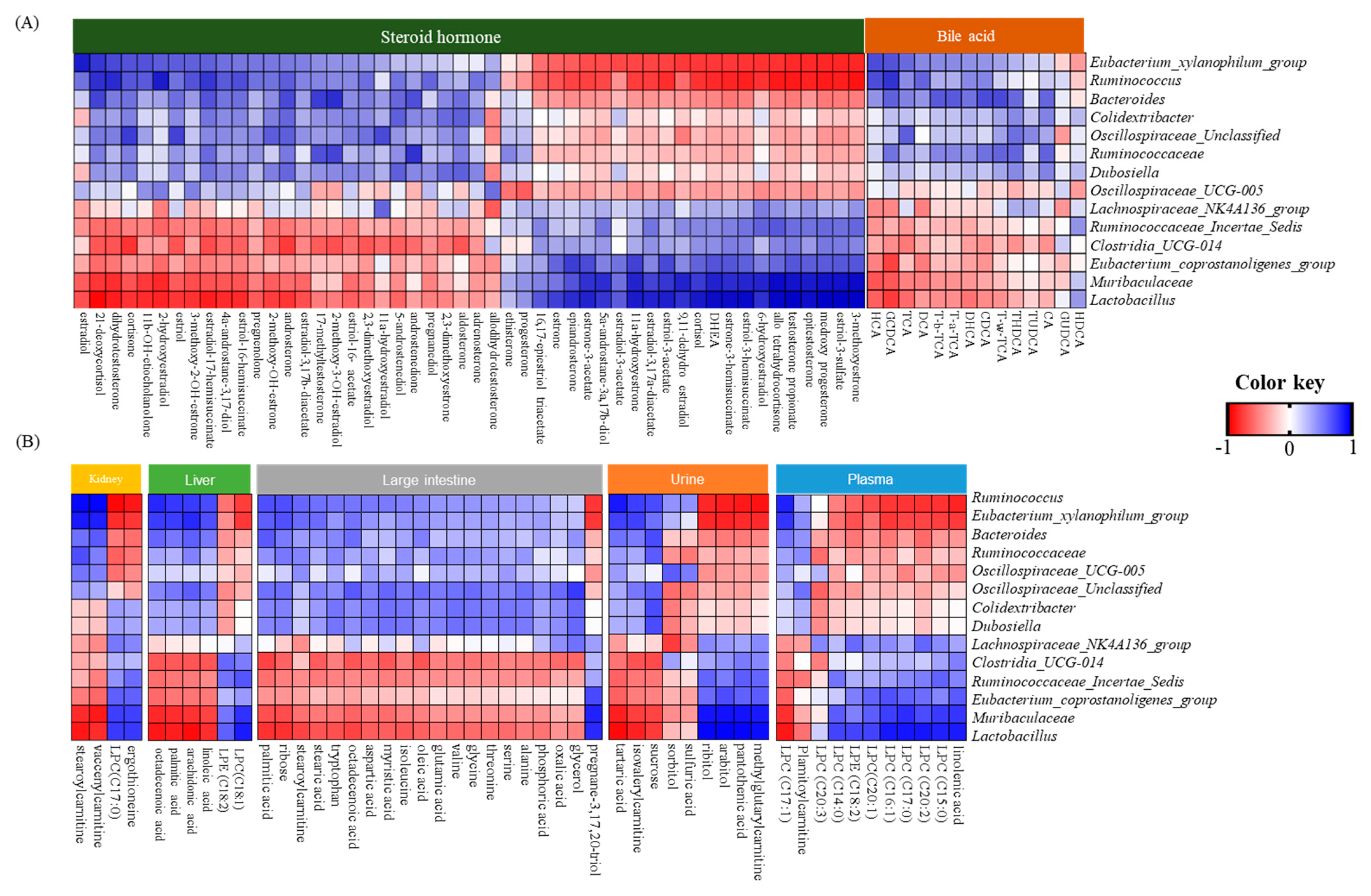
3.6. Correlations between Gut Microbiota, Bile Acids, Steroid Hormones, and Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, H.-J.; Lee, N.-K.; Park, H.-D. Health benefits of lactic acid bacteria isolated from kimchi, with respect to immunomodulatory effects. Food Sci. Biotechnol. 2015, 24, 783–789. [Google Scholar] [CrossRef]
- Choi, I.H.; Noh, J.S.; Han, J.-S.; Kim, H.J.; Han, E.-S.; Song, Y.O. Kimchi, a fermented vegetable, improves serum lipid profiles in healthy young adults: Randomized clinical trial. J. Med. Food 2013, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Bose, S.; Wang Jh Kim, B.S.; Kim, M.J.; Kim, E.J.; Kim, H. Contrasting effects of fresh and fermented kimchi consumption on gut microbiota composition and gene expression related to metabolic syndrome in obese Korean women. Mol. Nutr. Food Res. 2015, 59, 1004–1008. [Google Scholar] [CrossRef]
- Kim, E.K.; An, S.-Y.; Lee, M.-S.; Kim, T.H.; Lee, H.-K.; Hwang, W.S.; Choe, S.J.; Kim, T.-Y.; Han, S.J.; Kim, H.J. Fermented kimchi reduces body weight and improves metabolic parameters in overweight and obese patients. Nutr. Res. 2011, 31, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Sekhon, S.S.; Ko, J.H.; Kim, H.C.; Kim, S.Y.; Won, K.; Ahn, J.-Y.; Lee, K.; Kim, Y.-H. Lactic acid bacteria isolated from kimchi to evaluate anti-obesity effect in high fat diet-induced obese mice. Toxicol. Environ. Health Sci. 2018, 10, 11–16. [Google Scholar] [CrossRef]
- Shen, W.; Shen, M.; Zhao, X.; Zhu, H.; Yang, Y.; Lu, S.; Tan, Y.; Li, G.; Li, M.; Wang, J. Anti-obesity effect of capsaicin in mice fed with high-fat diet is associated with an increase in population of the gut bacterium Akkermansia muciniphila. Front. Microbiol. 2017, 8, 235091. [Google Scholar] [CrossRef]
- Hattori H, Moriyama A, Ohno T, Shibata T, Iwahashi H, Mitsunaga T: Molecular networking-based lipid profiling and multi-omics approaches reveal new contributions of functional vanilloids to gut microbiota and lipometabolism changes. Food Chem. Mol. Sci. 2022, 5, 100123. [CrossRef] [PubMed]
- Xie, C.; Teng, J.; Wang, X.; Xu, B.; Niu, Y.; Ma, L.; Yan, X. Multi-omics analysis reveals gut microbiota-induced intramuscular fat deposition via regulating expression of lipogenesis-associated genes. Anim. Nutr. 2022, 9, 84–99. [Google Scholar] [CrossRef]
- Kim, H.-J.; Kim, J.H.; Noh, S.; Hur, H.J.; Sung, M.J.; Hwang, J.-T.; Park, J.H.; Yang, H.J.; Kim, M.-S.; Kwon, D.Y. Metabolomic analysis of livers and serum from high-fat diet induced obese mice. J. Proteome Res. 2011, 10, 722–731. [Google Scholar] [CrossRef]
- Wu, J.; Wang, K.; Wang, X.; Pang, Y.; Jiang, C. The role of the gut microbiome and its metabolites in metabolic diseases. Protein Cell 2021, 12, 360–373. [Google Scholar] [CrossRef]
- Chanmuang, S.; Kim, B.-M.; Gu, S.-Y.; Son, Y.-J.; Le, H.-G.; Nam, Y.-D.; Song, E.-J.; Ham, K.-S.; Kim, H.-J. Effects of sea salt intake on metabolites, steroid hormones, and gut microbiota in rats. PLoS ONE 2022, 17, e0269014. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Lee, E.-S.; Song, E.-J.; Nam, Y.-D.; Nam, T.G.; Kim, H.-J.; Lee, B.-H.; Seo, M.-J.; Seo, D.-H. Effects of enzymatically modified chestnut starch on the gut microbiome, microbial metabolome, and transcriptome of diet-induced obese mice. Int. J. Biol. Macromol. 2020, 145, 235–243. [Google Scholar] [CrossRef]
- Park, S.-E.; Kwon, S.J.; Cho, K.-M.; Seo, S.-H.; Kim, E.-J.; Unno, T.; Bok, S.-H.; Park, D.-H.; Son, H.-S. Intervention with kimchi microbial community ameliorates obesity by regulating gut microbiota. J. Microbiol. 2020, 58, 859–867. [Google Scholar] [CrossRef]
- Kusada, H.; Kameyama, K.; Meng, X.-Y.; Kamagata, Y.; Tamaki, H. Fusimonas intestini gen. nov., sp. nov., a novel intestinal bacterium of the family Lachnospiraceae associated with diabetes in mice. Sci. Rep. 2017, 7, 18087. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Harrington, J.M.; Millar, S.R.; Perry, I.J.; O’Toole, P.W.; Phillips, C.M. Gut microbiota associations with metabolic health and obesity status in older adults. Nutrients 2020, 12, 2364. [Google Scholar] [CrossRef]
- Sorbara, M.T.; Littmann, E.R.; Fontana, E.; Moody, T.U.; Kohout, C.E.; Gjonbalaj, M.; Eaton, V.; Seok, R.; Leiner, I.M.; Pamer, E.G. Functional and genomic variation between human-derived isolates of Lachnospiraceae reveals inter-and intra-species diversity. Cell Host Microbe 2020, 28, 134–146.e134. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zheng, H.; Zhang, G.; Chen, F.; Chen, L.; Yang, Z. High Oscillospira abundance indicates constipation and low BMI in the Guangdong Gut Microbiome Project. Sci. Rep. 2020, 10, 9364. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira-a candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef] [PubMed]
- Indiani, C.M.d.S.P.; Rizzardi, K.F.; Castelo, P.M.; Ferraz, L.F.C.; Darrieux, M.; Parisotto, T.M. Childhood obesity and Firmicutes/Bacteroidetes ratio in the gut microbiota: A systematic review. Child. Obes. 2018, 14, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Song, E.-J.; Shin, N.R.; Jeon, S.; Nam, Y.-D.; Kim, H. Lorcaserin and phentermine exert anti-obesity effects with modulation of the gut microbiota. Front. Microbiol. 2023, 13, 1109651. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S.; Chiba, Y.; Wang, C.; Yamashiro, Y. The effects of the Lactobacillus casei strain on obesity in children: A pilot study. Benef. Microbes 2017, 8, 535–543. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Xia, S.; Li, L.; Li, Y.; Wang, P.; Shi, Y.; Le, G. Effects of different Lactobacillus reuteri on inflammatory and fat storage in high-fat diet-induced obesity mice model. J. Funct. Foods 2015, 14, 424–434. [Google Scholar] [CrossRef]
- Song, W.; Song, C.; Li, L.; Wang, T.; Hu, J.; Zhu, L.; Yue, T. Lactobacillus alleviated obesity induced by high-fat diet in mice. J. Food Sci. 2021, 86, 5439–5451. [Google Scholar] [CrossRef] [PubMed]
- Madan, R.; Petri, W.A., Jr. Role of obesity and adipose tissue-derived cytokine leptin during Clostridium difficile infection. Anaerobe 2015, 34, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Mulki, R.; Baumann, A.; Alnabelsi, T.; Sandhu, N.; Alhamshari, Y.; Wheeler, D.; Perloff, S.; Katz, P. Body mass index greater than 35 is associated with severe Clostridium difficile infection. Aliment. Pharmacol. Ther. 2017, 45, 75–81. [Google Scholar] [CrossRef]
- Woting, A.; Pfeiffer, N.; Loh, G.; Klaus, S.; Blaut, M. Clostridium ramosum promotes high-fat diet-induced obesity in gnotobiotic mouse models. MBio 2014, 5, 1110–1128. [Google Scholar] [CrossRef]
- Jiao, N.; Baker, S.S.; Nugent, C.A.; Tsompana, M.; Guan, L.; Wang, Y.; Buck, M.J.; Genco, R.J.; Baker, R.D.; Zhu, R. High-fat diet increases Clostridium clusters XIVa in obese rodents. FASEB J. 2017, 31, 965–969. [Google Scholar] [CrossRef]
- Park, J.E.; Oh, S.-H.; Cha, Y.-S. Lactobacillus brevis OPK-3 from kimchi prevents obesity and modulates the expression of adipogenic and pro-inflammatory genes in adipose tissue of diet-induced obese mice. Nutrients 2020, 12, 604. [Google Scholar] [CrossRef]
- Lim, S.; Moon, J.H.; Shin, C.M.; Jeong, D.; Kim, B. Effect of Lactobacillus sakei, a probiotic derived from kimchi, on body fat in Koreans with obesity: A randomized controlled study. Endocrinol. Metab. 2020, 35, 425. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.S.; Choi, J.H.; Sung, M.J.; Hur, J.Y.; Hur, H.J.; Park, J.D.; Kim, Y.C.; Gu, E.J.; Min, B.; Kim, H.J. Green tea changes serum and liver metabolomic profiles in mice with high-fat diet-induced obesity. Mol. Nutr. Food Res. 2015, 59, 784–794. [Google Scholar] [CrossRef]
- Lord, R.S.; Burdette, C.K.; Bralley, J.A. Significance of urinary tartaric acid. Clin. Chem. 2005, 51, 672–673. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-López, I.; Parilli-Moser, I.; Arancibia-Riveros, C.; Tresserra-Rimbau, A.; Martínez-González, M.A.; Ortega-Azorín, C.; Salas-Salvadó, J.; Castañer, O.; Lapetra, J.; Arós, F. Urinary tartaric acid, a biomarker of wine intake, correlates with lower total and LDL cholesterol. Nutrients 2021, 13, 2883. [Google Scholar] [CrossRef]
- Ajao, F.O.; Iyedupe, M.O. Effect of high salt intake on plasma lipid profile in pregnant wistar rats. Int. J. Physiol. Pathophysiol. Pharmacol. 2020, 12, 147–152. [Google Scholar] [PubMed]
- Yun, Y.-R.; Lee, J.J.; Lee, H.J.; Choi, Y.-J.; Lee, J.-H.; Park, S.J.; Park, S.H.; Seo, H.-Y.; Min, S.G. Comparison of quality characteristics of commercial kimchi manufactured in Korea, China, and the United States. Foods 2021, 10, 2488. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Andreu-Sánchez, S.; Kuipers, F.; Fu, J. Gut microbiome and bile acids in obesity-related diseases. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101493. [Google Scholar] [CrossRef]
- Moghetti, P.; Castello, R.; Negri, C.; Tosi, F.; Perrone, F.; Caputo, M.; Zanolin, E.; Muggeo, M. Metformin effects on clinical features, endocrine and metabolic profiles, and insulin sensitivity in polycystic ovary syndrome: A randomized, double-blind, placebo-controlled 6-month trial, followed by open, long-term clinical evaluation. J. Clin. Endocrinol. Metab. 2000, 85, 139–146. [Google Scholar]
- Derby, C.A.; Zilber, S.; Brambilla, D.; Morales, K.H.; McKinlay, J.B. Body mass index, waist circumference and waist to hip ratio and change in sex steroid hormones: The Massachusetts Male Ageing Study. Clin. Endocrinol. 2006, 65, 125–131. [Google Scholar] [CrossRef]
- Tchernof, A.; Després, J.-P. Sex steroid hormones, sex hormone-binding globulin, and obesity in men and women. Horm. Metab. Res. 2000, 32, 526–536. [Google Scholar] [CrossRef]
- Lizcano, F.; Guzmán, G. Estrogen deficiency and the origin of obesity during menopause. BioMed Res. Int. 2014, 2014, 757461. [Google Scholar] [CrossRef] [PubMed]
- Sumińska, M.; Podgórski, R.; Fichna, P.; Mazur, A.; Fichna, M. The Impact of Obesity on the Excretion of Steroid Metabolites in Boys and Girls: A Comparison with Normal-Weight Children. Nutrients 2023, 15, 1734. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Huang, F.; Zhao, L.; Zhang, Y.; Yang, W.; Wang, S.; Li, M.; Han, X.; Ge, K.; Qu, C. A dysregulated bile acid-gut microbiota axis contributes to obesity susceptibility. EBioMedicine 2020, 55, 102766. [Google Scholar] [CrossRef] [PubMed]
- Wahlström, A.; Sayin, S.I.; Marschall, H.-U.; Bäckhed, F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, Y.; Ma, F.; Sun, M.; Mu, G.; Tuo, Y. The ameliorative effect of Lactobacillus plantarum Y44 oral administration on inflammation and lipid metabolism in obese mice fed with a high fat diet. Food Funct. 2020, 11, 5024–5039. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ND | HFD | KHD | |
|---|---|---|---|
| weight gain (g) | 199.55 ± 19.22 a | 237.57 ± 19.20 b | 208.71 ± 20.49 ab |
| food intake (g/ day) | 46.8 ± 0.89 b | 30.21 ± 2.29 a | 30.6 ± 1.556 a |
| spleen (g) | 0.72 ± 0.08 a | 0.7 ± 0.11 a | 0.6 ± 0.11 a |
| liver (g) | 11.85 ± 0.85 a | 11.08 ± 1.25 a | 11.30 ± 1.38 a |
| kidney (g) | 1.23 ± 0.39 a | 1.23 ± 0.09 a | 1.20 ± 0.14 a |
| epididymal adipose tissue (g) | 5.63 ± 1.08 a | 9.30 ± 2.95 b | 9.27 ± 1.52 b |
| AST (U/L) | 57.22 ± 7.47 a | 60.29 ± 15.59 a | 67.57 ± 10.67 a |
| ALT (U/L) | 38.33 ± 4.18 b | 28.28 ± 4.60 a | 31.00 ± 7.52 a |
| TCHO (mg/dL) | 114.44 ± 19.37 a | 111.57± 12.06 a | 112.85± 12.26 a |
| TG (mg/dL) | 45.11 ± 12.02 b | 36.14 ± 11.68 a | 35.71 ± 10.89 ab |
| HDL (mg/dL) | 45.88 ± 8.96 a | 49.43 ± 5.06 a | 59.14 ± 33.28 a |
| LDH (U/L) | 97.67± 35.01 a | 105.71 ± 36.76 a | 133.57 ± 77.58 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-W.; Nguyen, Q.-A.; Chanmuang, S.; Lee, S.-B.; Kim, B.-M.; Lee, H.-J.; Jang, G.-J.; Kim, H.-J. Effects of Kimchi Intake on the Gut Microbiota and Metabolite Profiles of High-Fat-Induced Obese Rats. Nutrients 2024, 16, 3095. https://doi.org/10.3390/nu16183095
Kim D-W, Nguyen Q-A, Chanmuang S, Lee S-B, Kim B-M, Lee H-J, Jang G-J, Kim H-J. Effects of Kimchi Intake on the Gut Microbiota and Metabolite Profiles of High-Fat-Induced Obese Rats. Nutrients. 2024; 16(18):3095. https://doi.org/10.3390/nu16183095
Chicago/Turabian StyleKim, Dong-Wook, Quynh-An Nguyen, Saoraya Chanmuang, Sang-Bong Lee, Bo-Min Kim, Hyeon-Jeong Lee, Gwang-Ju Jang, and Hyun-Jin Kim. 2024. "Effects of Kimchi Intake on the Gut Microbiota and Metabolite Profiles of High-Fat-Induced Obese Rats" Nutrients 16, no. 18: 3095. https://doi.org/10.3390/nu16183095
APA StyleKim, D.-W., Nguyen, Q.-A., Chanmuang, S., Lee, S.-B., Kim, B.-M., Lee, H.-J., Jang, G.-J., & Kim, H.-J. (2024). Effects of Kimchi Intake on the Gut Microbiota and Metabolite Profiles of High-Fat-Induced Obese Rats. Nutrients, 16(18), 3095. https://doi.org/10.3390/nu16183095