
Vitamin D Maintains Growth and Bone Mineral Density against a Background of Severe Vitamin A Deficiency and Moderate Toxicity in a Swine Model
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
3. Results
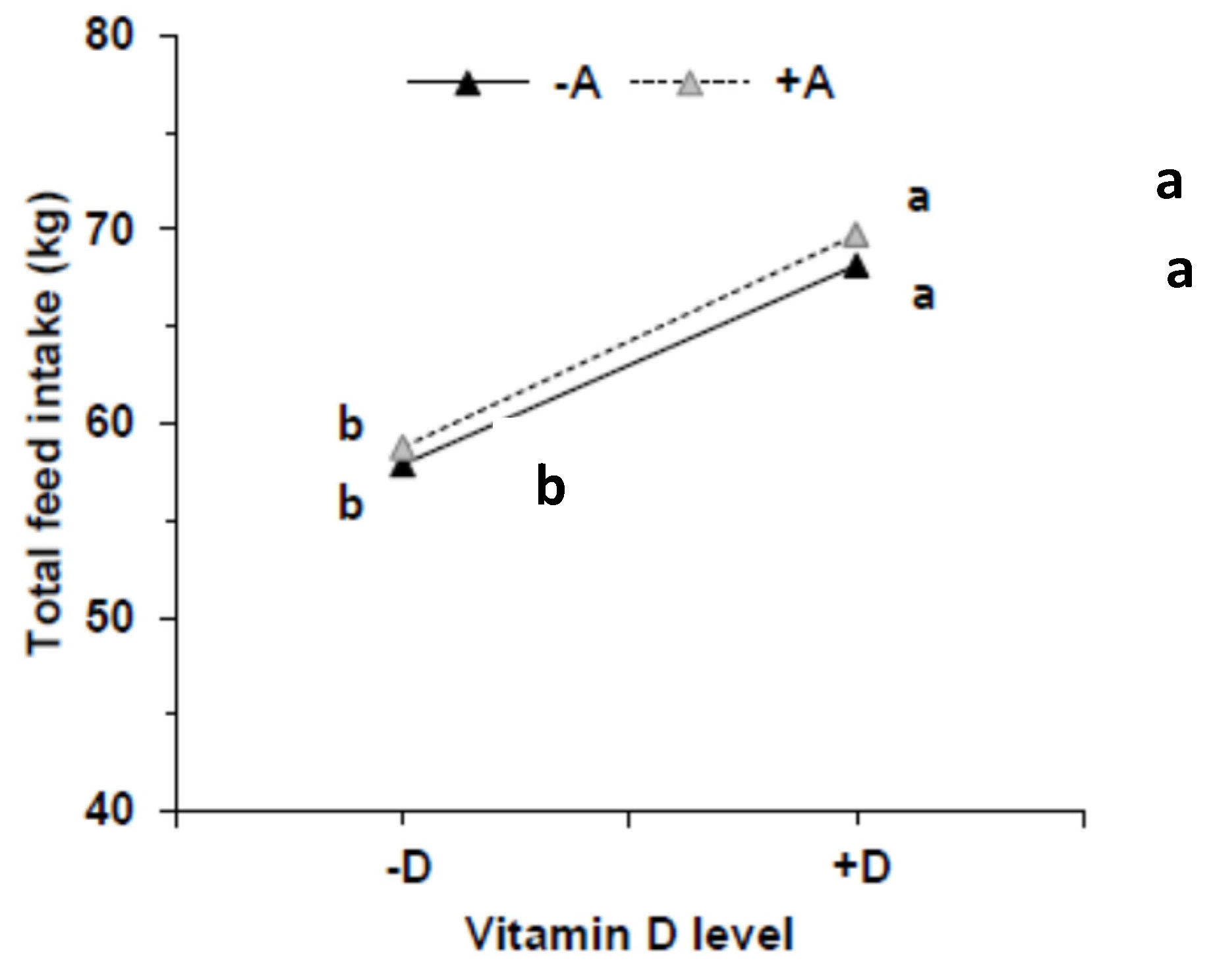
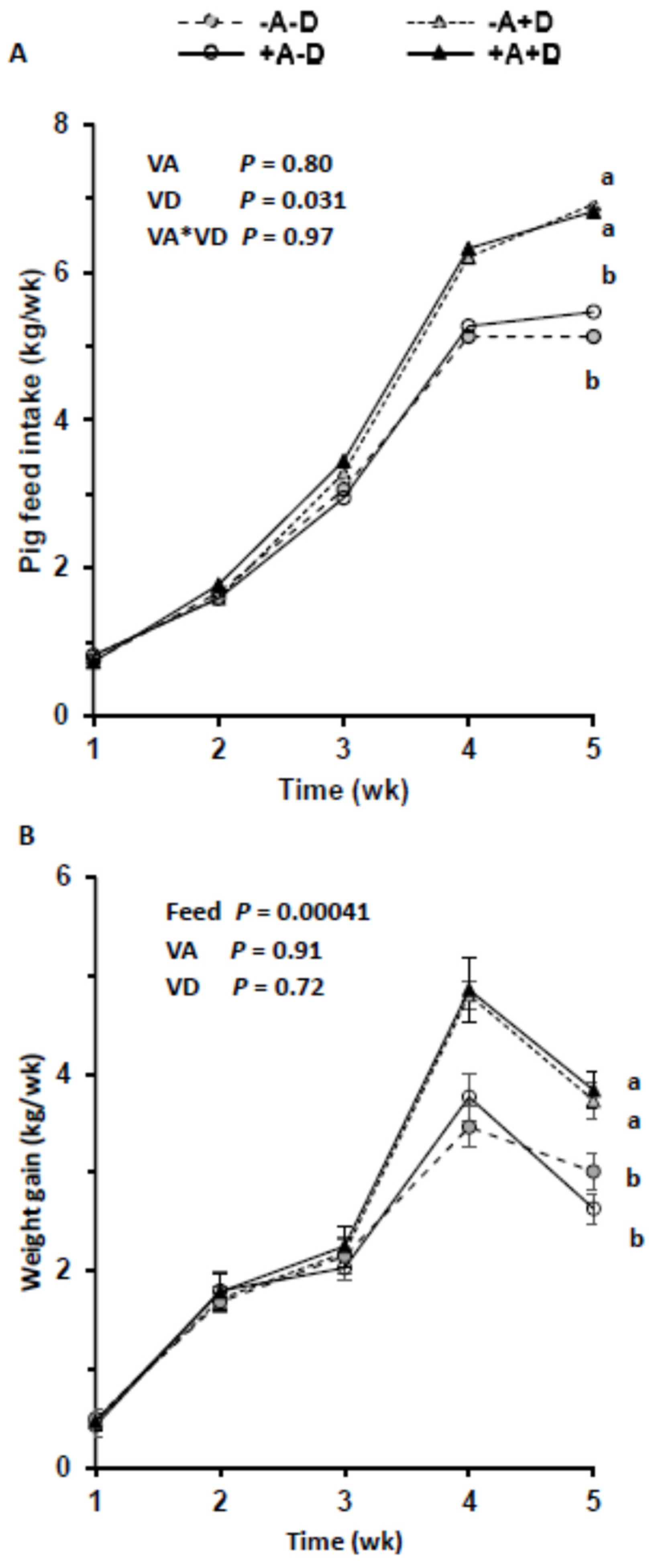
3.1. Feed Intake, Growth, and Kyphosis
3.2. Liver Vitamin A
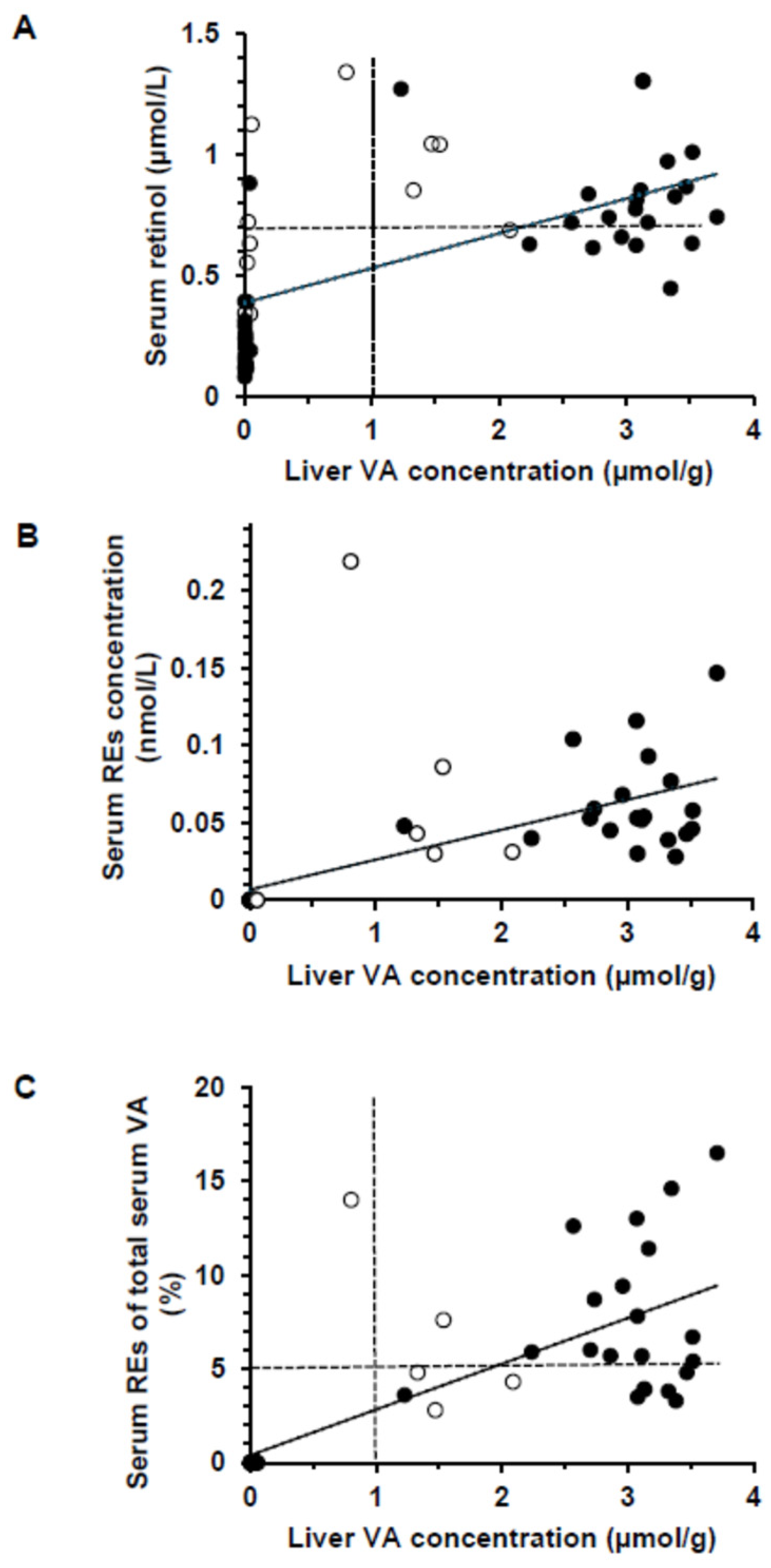
3.3. Serum Retinol, Retinyl Esters, and Vitamin D
3.4. Extrahepatic Vitamins A and D
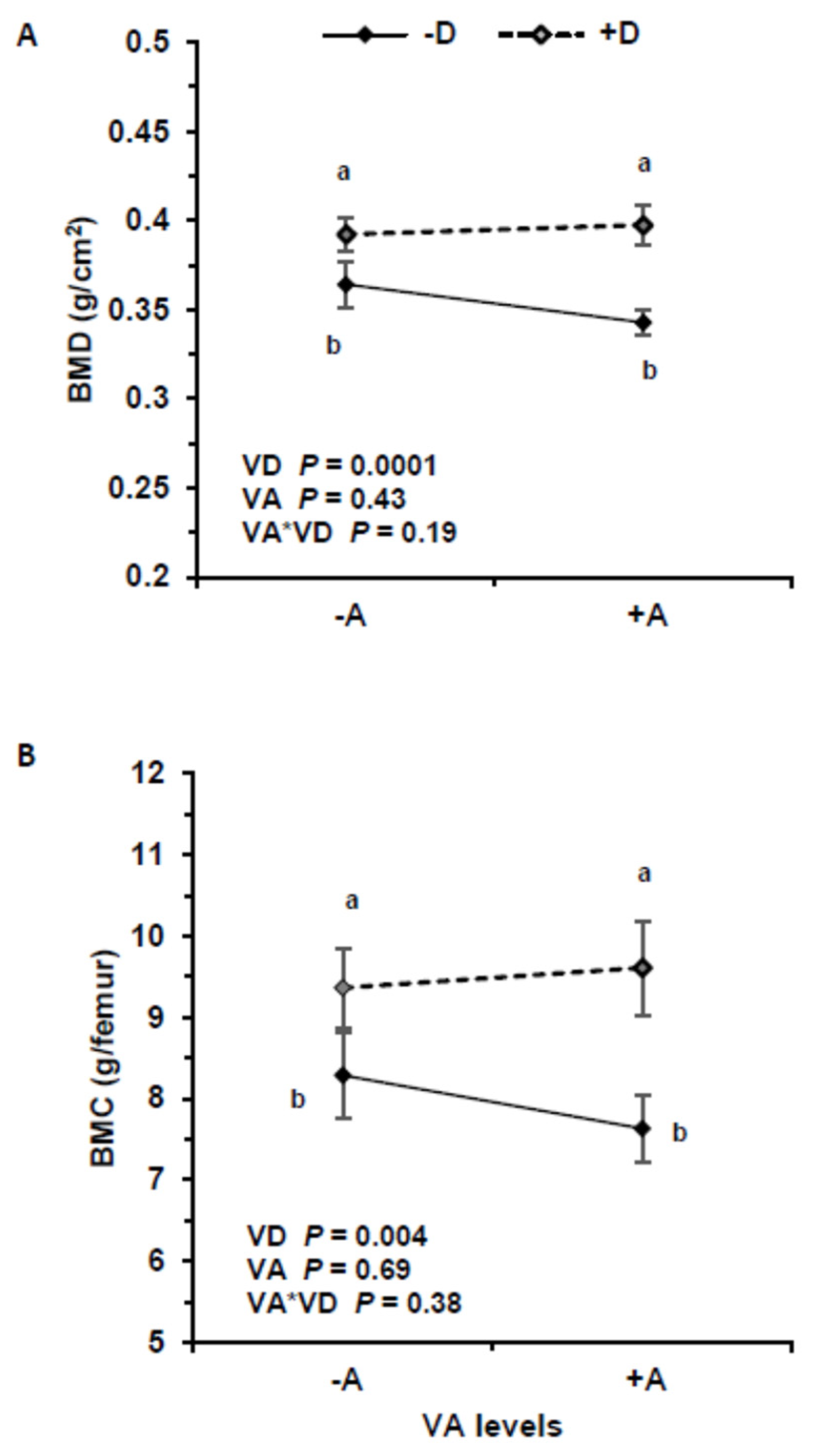
3.5. Bone Mineral Density and Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Melhus, H.; Michaelsson, K.; Kindmark, A.; Bergstrom, R.; Holmberg, L.; Mallmin, H.; Wolk, A.; Ljunghall, S. Excessive dietary intake of vitamin A is associated with reduced bone mineral density and increased risk for hip fracture. Ann. Intern. Med. 1998, 129, 770–778. [Google Scholar] [CrossRef]
- Feskanich, D.; Singh, V.; Willet, W.C.; Colditz, G.C. Vitamin A intake and hip fractures among postmenopausal women. JAMA 2002, 287, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Wright, N.C.; Looker, A.C.; Saag, K.G.; Curtis, J.R.; Delzell, E.S.; Randall, S.; Dawson-Hughes, B. The recent prevalence of osteoporosis and low bone mass in the United States based on bone mineral density at the femoral neck or lumbar spine. J. Bone Miner. Res. 2014, 29, 2520–2526. [Google Scholar] [CrossRef]
- DeLuca, H.F. Overview of general physiologic features and functions of vitamin D. Am. J. Clin. Nutr. 2004, 80, 1689S–1696S. [Google Scholar] [CrossRef]
- Holick, M.F. Vitamin D status: Measurement, interpretation and clinical application. Ann. Epidemiol. 2009, 19, 73–78. [Google Scholar] [CrossRef]
- Arnold, J.; Madison, D.M.; Ensley, S.M.; Golf, J.P. Survey of serum vitamin D status across stages of swine production and evaluation of supplemental vitamin D premixes used in swine diets. J. Swine Health Prod. 2015, 23, 23–34. Available online: https://www.aasv.org/shap/issues/v23n1/v23n1p28.html (accessed on 12 May 2023).
- Promislow, J.H.; Goodman-Gruen, D.; Slymen, D.J.; Barrett-Connor, E. Retinol intake and bone mineral density in the elderly: The Rancho Bernardo Study. J. Bone Miner. Res. 2002, 17, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Yee, M.M.F.; Chin, K.-Y.; Ima-Nirwana, S.; Wong, S.K. Vitamin A and bone health: A review on current evidence. Molecules 2021, 26, 1757. [Google Scholar] [CrossRef] [PubMed]
- Henning, P.; Conaway, H.H.; Lerner, U.H. Retinoid receptors in bone and their role in bone remodeling. Front. Endo. 2015, 6, 31. [Google Scholar] [CrossRef]
- Wu, A.M.; Huang, C.Q.; Lin, Z.K.; Tian, N.F.; Ni, W.F.; Wang, X.Y.; Xu, H.; Chi, L. The relationship between vitamin A and risk of fracture: Meta-analysis of prospective studies. J. Bone Miner. Res. 2014, 29, 2031–2039. [Google Scholar] [CrossRef]
- Frankel, T.L.; Seshadri, M.S.; McDowall, D.B.; Cornish, C.J. Hypervitaminosis A and calcium-regulated hormones in the rat. J. Nutr. 1986, 116, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Baineni, R.; Gulati, R.; Delhi, C.G.K. Vitamin A toxicity presenting as bone pain. Arch. Dis. Child. 2017, 102, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Carrigan, M.J.; Glastonbury, J.R.W.; Evers, J.V. Hypovitaminosis A in pigs. Aust. Vet. J. 1988, 65, 158–159. [Google Scholar] [CrossRef] [PubMed]
- Lakritz, J.; George, L.W.; Moore, J. Seizures and acute death attributable to hypovitaminosis A and suspected hypovitaminosis D in feeder pigs. JAVMA 1993, 202, 1276–1278. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Prevalence of Vitamin A Deficiency in Populations at Risk 1995–2005: Global Database on Vitamin A Deficiency. 2009. Available online: https://apps.who.int/iris/handle/10665/44110 (accessed on 14 February 2024).
- World Health Organization. Guideline: Vitamin A Supplementation in Infants and Children 6–59 Months of Age. 2011. Available online: https://apps.who.int/iris/handle/10665/44664 (accessed on 14 February 2024).
- Klemm, R.D.; Palmer, A.C.; Greig, A.; Engle-Stone, R.; Dalmiya, N. A changing landscape for vitamin A programs: Implications for optimal intervention packages, program monitoring, and safety. Food Nutr. Bull. 2016, 37, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Tanumihardjo, S.A.; Kaliwile, C.; Boy, E.; Dhansay, M.A.; van Stuijvenberg, M.E. Overlapping vitamin A interventions in the United States, Guatemala, Zambia, and South Africa: Case studies. Ann. N. Y. Acad. Sci. 2019, 1446, 102–116. [Google Scholar] [CrossRef] [PubMed]
- National Statistical Office (NSO). [Malawi]; ICF Malawi Demographic and Health Survey 2015–2016. 2018. Available online: https://dhsprogram.com/publications/citations/cite_FR319.enw (accessed on 24 April 2023).
- Williams, A.M.; Tanumihardjo, S.A.; Rhodes, E.C.; Mapango, C.; Kazembe, B.; Phiri, F.; Kang’ombe, D.D.; Sheftel, J.; Orchardson, V.; Tripp, K.; et al. Vitamin A deficiency has declined in Malawi, but with evidence of elevated vitamin A in children. Am. J. Clin. Nutr. 2021, 113, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Priemel, M.; von Domarus, C.; Klatte, T.O.; Kessler, J.; Schlie, J.; Meier, S.; Proksch, N.; Pastor, F.; Netter, C.; Streichert, T.; et al. Bone mineralization defects and vitamin D deficiency: Histomorphometric analysis of iliac crest bone biopsies and circulating 25-hydroxyvitamin D in 675 patients. J. Bone Miner. Res. 2010, 25, 305–312. [Google Scholar] [CrossRef]
- Lunney, J.K.; Goor, A.V.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef]
- Amundson, L.A.; Hernandez, L.L.; Laporta, J.; Crenshaw, T.D. Maternal dietary vitamin D carry-over alters offspring growth, skeletal mineralisation and tissue mRNA expressions of genes related to vitamin D, calcium and phosphorus homoeostasis in swine. Br. J. Nutr. 2016, 116, 774–787. [Google Scholar] [CrossRef]
- Greaves, R.F.; Woollard, G.A.; Hoad, K.E.; Walmsley, T.A.; Johnson, L.A.; Briscoe, S.; Koetsier, S.; Harrower, T.; Gill, J.P. Laboratory medicine best practice guideline: Vitamins A, E, and the carotenoids in blood. Clin. Biochem. Rev. 2014, 35, 81–114. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4159783/ (accessed on 12 May 2023). [PubMed]
- Riabroy, N.; Tanumihardjo, S.A. Oral doses of α-retinyl ester track chylomicron uptake and distribution of vitamin A in a male piglet model for newborn infants. J. Nutr. 2014, 144, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Rortvedt, L.A.; Crenshaw, T.D. Expression of kyphosis in young pigs is altered by vitamin D supplementation of maternal diets. J. Anim. Sci. 2012, 90, 4905–4915. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012; 226p.
- Halanski, M.A.; Hildahl, B.; Amundson, L.A.; Leiferman, E.; Gendron-Fitzpatrick, A.; Chaudhary, R.; Hartwig-Stokes, H.M.; McCabe, R.; Lenhart, R.; Chin, M.; et al. Maternal diets deficient in vitamin D increase the risk of kyphosis in offspring: A novel kyphotic porcine model. J. Bone Jt. Surg. 2018, 100, 406–415. [Google Scholar] [CrossRef]
- Witschi, A.-K.M.; Liesegang, A.; Gebert, S.; Weber, G.M.; Wenk, C. Effect of source and quantity of dietary vitamin D in maternal and creep diets on bone metabolism and growth in piglets. J. Anim. Sci. 2011, 89, 1844–1852. [Google Scholar] [CrossRef] [PubMed]
- Rohde, C.M.; DeLuca, H.F. Bone resorption activity of all-trans-retinoic acid is independent of vitamin D in rats. J. Nutr. 2003, 133, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Valverde, C.; Caballero-Villarraso, J.; Mata-Granados, J.M.; Casado-Díaz, J.M.; Sosa-Henríquez, A.; Malouf-Sierra, M.; Nogués-Solán, X.; Rodríguez-Mañas, L.; Cortés-Gil, X.; Delgadillo-Duarte, J.; et al. High serum retinol as a relevant contributor to low bone mineral density in postmenopausal osteoporotic women. Calcif. Tissue Int. 2018, 102, 651–656. [Google Scholar] [CrossRef]
- Michaëlsson, K.; Lithell, H.; Vessby, B.; Melhus, H. Serum retinol levels and the risk of fracture. N. Engl. J. Med. 2003, 348, 287–294. [Google Scholar] [CrossRef]
- Lim, L.S.; Harnack, L.J.; Lazovich, D.; Folsom, A.R. Vitamin A intake and the risk of hip fracture in postmenopausal women: The Iowa Women’s Health Study. Osteoporos. Int. 2004, 15, 552–559. [Google Scholar] [CrossRef]
- Rejnmark, L.; Vestergaard, P.; Charles, P.; Hermann, A.P.; Brot, C.; Eiken, P.; Mosekilde, L. No effect of vitamin A intake on bone mineral density and fracture risk in perimenopausal women. Osteoporos. Int. 2004, 15, 872–880. [Google Scholar] [CrossRef]
- Johansson, S.; Melhus, H. Vitamin A antagonizes calcium response to vitamin D in man. J. Bone Miner. Res. 2001, 16, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Ciresi, A.; Giordano, C. Vitamin D across growth hormone (GH) disorders: From GH deficiency to GH excess. Growth Horm. IGF Res. 2017, 33, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Leonardi, A.; Lanciotti, L.; Cofini, M.; Muzi, G.; Penta, L. Vitamin D and growth hormone in children: A review of the current scientific knowledge. J. Transl. Med. 2019, 17, 87. [Google Scholar] [CrossRef] [PubMed]
- Wray, A.E.; Okita, N.; Ross, A.C. Cortical and trabecular bone, bone mineral density, and resistance to ex vivo fracture are not altered in response to life-long vitamin A supplementation in aging rats. J. Nutr. 2011, 141, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Lionikaite, V.; Gustafsson, K.L.; Westerlund, A.; Windahl, S.H.; Koskela, A.; Tuukkanen, J.; Johansson, H.; Ohlsson, C.; Conaway, H.H.; Henning, P.; et al. Clinically relevant doses of vitamin A decrease cortical bone mass in mice. J. Endocrinol. 2018, 239, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Sherman, A.; Monari-Sparks, M.J.; Schweiker, O.; Hunter, K. Correction of low vitamin D improves fatigue: Effect of correction of low vitamin D in fatigue study (EViDiF study). N. Am. J. Med. Sci. 2014, 6, 396–402. [Google Scholar] [CrossRef]
- Fraser, D.R. Vitamin D deficiency and energy metabolism. Endocrinology 2015, 156, 1933–1935. [Google Scholar] [CrossRef]
- Funakoshi, S.; Yoshimura, K.; Hirano, S.; Ohmi, S.; Amano, E.; Fukuda, Y.; Terada, Y.; Fujimoto, S. Undercarboxylated osteocalcin correlates with insulin secretion in Japanese individuals with diabetes. Diabetol. Metab. Syndr. 2020, 12, 72. [Google Scholar] [CrossRef] [PubMed]
- Singhai, A.; Banzal, S. Severe vitamin D deficiency causing kyphoscoliosis. J. Fam. Med. Prim. Care 2013, 2, 384–386. [Google Scholar] [CrossRef]
- Meyer, M.; Pike, J.W. Mechanistic homeostasis of vitamin D metabolism in the kidney through reciprocal modulation of Cyp27b1 and Cyp24a1 expression. J. Steroid Biochem. Mol. Biol. 2020, 196, 105500. [Google Scholar] [CrossRef]
- Schwetz, V.; Trummer, C.; Pandis, M.; Grübler, M.R.; Verheyen, N.; Gaksch, M.; Zittermann, A.; März, W.; Aberer, F.; Lang, A.; et al. Effects of vitamin D supplementation on bone turnover markers: A randomized controlled trial. Nutrients 2017, 9, 432. [Google Scholar] [CrossRef] [PubMed]
- Tanumihardjo, S.A.; Gannon, B.M.; Kaliwile, C.; Chileshe, J.; Binkley, N.C. Restricting vitamin A intake increases bone formation in Zambian children with high liver stores of vitamin. Arch. Osteoporos. 2019, 14, 72. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.; Johansson, S.; Branting, M.; Melhus, H. Vitamin A differentially regulates RANKL and OPG expression in human osteoblasts. Biochem. Biophys. Res. Commun. 2008, 322, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Taylan, A.; Birlik, M.; Kenar, G.; Toprak, B.; Gundogdu, B.; Gurler, O.; Karakas, B.; Akıncı, B.; Sisman, A.R. Osteoprotegrin interacts with biomarkers and cytokines that have roles in osteoporosis, skin fibrosis, and vasculopathy in systemic sclerosis: A potential multifaceted relationship between OPG/RANKL/TRAIL and Wnt inhibitors. Mod. Rheumatol. 2019, 29, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophsy. 2008, 473, 139–146. [Google Scholar] [CrossRef]
- Tang, Y.; Wu, X.; Lei, W.; Pang, L.; Wan, C.; Shi, Z.; Zhao, L.; Nagy, T.R.; Peng, X.; Hu, J.; et al. TGF-β1–induced migration of bone mesenchymal stem cells couples bone resorption with formation. Nat. Med. 2009, 15, 757–765. [Google Scholar] [CrossRef]
- Hardy, E.; Fernandez-Patron, C. Destroy to rebuild: The connection between bone tissue remodeling and matrix metalloproteinases. Front. Physiol. 2020, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Deng, C.; Li, Y.P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed]
- Lind, T.; Sundqvist, A.; Hu, L.; Pejler, G.; Andersson, G.; Jacobson, A.; Melhus, H. Vitamin A is a negative regulator of osteoblast mineralization. PLoS ONE 2013, 8, e82388. [Google Scholar] [CrossRef]
- Gupta, P.M.; Madewell, Z.J.; Gannon, B.M.; Grahn, M.; Akelo, V.; Onyango, D.; Mahtab, S.; Madhi, S.A.; Giri, J.; Blau, D.M.; et al. Hepatic vitamin A concentrations and association with infectious causes of child death. J. Pediatr. 2024, 265, 113816. [Google Scholar] [CrossRef]
- World Health Organization. Serum Retinol Concentrations for Determining the Prevalence of Vitamin A Deficiency in Populations. Vitamin and Mineral Nutrition Information System. 2011. Available online: https://www.who.int/publications/i/item/WHO-NMH-NHD-MNM-11.3 (accessed on 20 February 2024).
- Skalny, A.V.; Aschner, M.; Tsatsakis, A.; Rocha, J.B.T.; Santamaria, A.; Spandidos, D.A.; Martins, A.C.; Lu, R.; Korobeinikova, T.V.; Chen, W.; et al. Role of vitamins beyond vitamin D3 in bone health and osteoporosis (Review). Int. J. Mol. Med. 2024, 53, 9. [Google Scholar] [CrossRef]
- Crandall, C. Vitamin A intake and osteoporosis: A Clinical Review. J. Women’s Health 2004, 13, 939–953. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients, g/kg | Phase 1 Nursery | Phase 2 Grower |
|---|---|---|
| Corn grain | 394.8 | 625.4 |
| Soybean meal, 48% crude protein | 307.2 | 315.7 |
| Oat groats | 150.0 | - |
| Whey | 100.0 | - |
| L-Lysine HCl | 2.5 | 3.0 |
| Methionine | 1.25 | 1.0 |
| Threonine | 1.0 | 1.0 |
| Monocalcium phosphate | 4.50 | 10.6 |
| Calcium carbonate | 5.24 | 8.31 |
| Corn oil | 20.0 | 20.0 |
| Sodium chloride | 3.50 | 5.00 |
| UW VTM-G without A and D3 2,3 | 10.0 | 10.0 |
| Total | 1000.0 | 1000.0 |
| Biomarker | Treatment Duration (wk) | Treatment Group 1 | p-Value | |||
|---|---|---|---|---|---|---|
| −A−D | −A+D | +A−D 2 | +A+D | |||
| Serum retinol 3 (µmol/L) | 3 | 0.67 ± 0.18 | 0.75 ± 0.37 | 0.64 ± 0.085 | 1.09 ± 0.43 | VA < 0.0001 VD = 0.32 VA*VD = 0.68 |
| 5 | 0.20 ± 0.087 | 0.22 ± 0.10 | 0.81 ± 0.15 | 0.84 ± 0.27 | ||
| Serum RE 3 (µmol/L) | 3 | ND | ND | 0.065 ± 0.030 | 0.093 ± 0.11 | VA < 0.0001 VD = 0.16 VA*VD = 0.17 |
| 5 | ND | ND | 0.051 ± 0.021 | 0.073 ± 0.035 | ||
| RE % of total 3 serum VA | 3 | ND | ND | 6.2 ± 2.0 | 7.0 ± 6.0 | VA < 0.0001 VD = 0.15 VA*VD = 0.17 |
| 5 | ND | ND | 6.4 ± 2.8 | 8.9 ± 4.8 | ||
| Serum 25(OH)D3 (ng/mL) | 3 | 3.15 ± 4.97 | 19.8 ± 4.20 | 2.16 ± 0.95 | 16.1 ± 5.47 | VA = 0.31 VD < 0.0001 VA*VD = 0.19 |
| 5 | 1.5 ± 1.11 | 27.7 ± 8.91 | 1.73 ± 0.59 | 23.9 ± 6.67 | ||
| Tissue 1 | Treatment Duration (wk) | −A−D | −A+D | +A−D 2 | +A+D | p |
|---|---|---|---|---|---|---|
| Liver (µmol) | 3 | 9.76 ± 3.19 | 10.3 ± 6.61 | 331 ± 95.6 | 426 ± 310 | VA < 0.0001 VD = 0.28 VA*VD = 0.26 |
| 5 | 5.9 ± 5.83 | 7.17 ± 12.4 | 1170 ± 331 | 1394 ± 392 | ||
| Kidney (nmol) | 3 | 35.7 ± 13.9 | 34.2 ± 1.51 2 | 75.1 ± 25.3 | 101 ± 55.6 | VA < 0.0001 VD = 0.03 VA*VD = 0.06 |
| 5 | 51.0 ± 24.9 | 68.0 ± 35.7 | 267 ± 65.3 | 418 ± 188 | ||
| Lung (nmol) | 3 | 7.07 ± 1.66 | 8.72 ± 5.24 | 36.5 ± 2.13 | 46.8 ± 27.2 | VA < 0.0001 VD = 0.57 VA*VD = 0.91 |
| 5 | 18.1 ± 12.2 | 9.54 ± 5.71 | 125 ± 61.2 | 123 ± 44.6 | ||
| Spleen (nmol) | 3 | 1.79 ± 2.02 | 0.41 ± 0.53 | 3.43 ± 0.74 | 10.2 ± 7.06 | VA < 0.0001 VD = 0.44 VA*VD = 0.73 |
| 5 | 0.57 ± 0.63 | 0.29 ± 0.46 | 15.6 ± 11.4 | 12.1 ± 3.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phiri, C.B.; Davis, C.R.; Grahn, M.; Gannon, B.M.; Kokinos, B.P.; Crenshaw, T.D.; Tanumihardjo, S.A. Vitamin D Maintains Growth and Bone Mineral Density against a Background of Severe Vitamin A Deficiency and Moderate Toxicity in a Swine Model. Nutrients 2024, 16, 2037. https://doi.org/10.3390/nu16132037
Phiri CB, Davis CR, Grahn M, Gannon BM, Kokinos BP, Crenshaw TD, Tanumihardjo SA. Vitamin D Maintains Growth and Bone Mineral Density against a Background of Severe Vitamin A Deficiency and Moderate Toxicity in a Swine Model. Nutrients. 2024; 16(13):2037. https://doi.org/10.3390/nu16132037
Chicago/Turabian StylePhiri, Cacious B., Christopher R. Davis, Michael Grahn, Bryan M. Gannon, Brittney P. Kokinos, Thomas D. Crenshaw, and Sherry A. Tanumihardjo. 2024. "Vitamin D Maintains Growth and Bone Mineral Density against a Background of Severe Vitamin A Deficiency and Moderate Toxicity in a Swine Model" Nutrients 16, no. 13: 2037. https://doi.org/10.3390/nu16132037
APA StylePhiri, C. B., Davis, C. R., Grahn, M., Gannon, B. M., Kokinos, B. P., Crenshaw, T. D., & Tanumihardjo, S. A. (2024). Vitamin D Maintains Growth and Bone Mineral Density against a Background of Severe Vitamin A Deficiency and Moderate Toxicity in a Swine Model. Nutrients, 16(13), 2037. https://doi.org/10.3390/nu16132037