
A Predictive Tool Based on DNA Methylation Data for Personalized Weight Loss through Different Dietary Strategies: A Pilot Study
 , , and
, , and 
Highlights
- DNA methylation in blood cells predicts the percentage of weight loss via two different 4-month hypocaloric strategies.
- Epigenetic biomarkers may be used for precision nutrition and the design of personalized dietary strategies to reduce obesity.
- A prediction model that includes epigenetic, genetic, and microbiota data may provide advantages for their implementation in precision nutrition.
Abstract
:1. Introduction
2. Materials and Methods
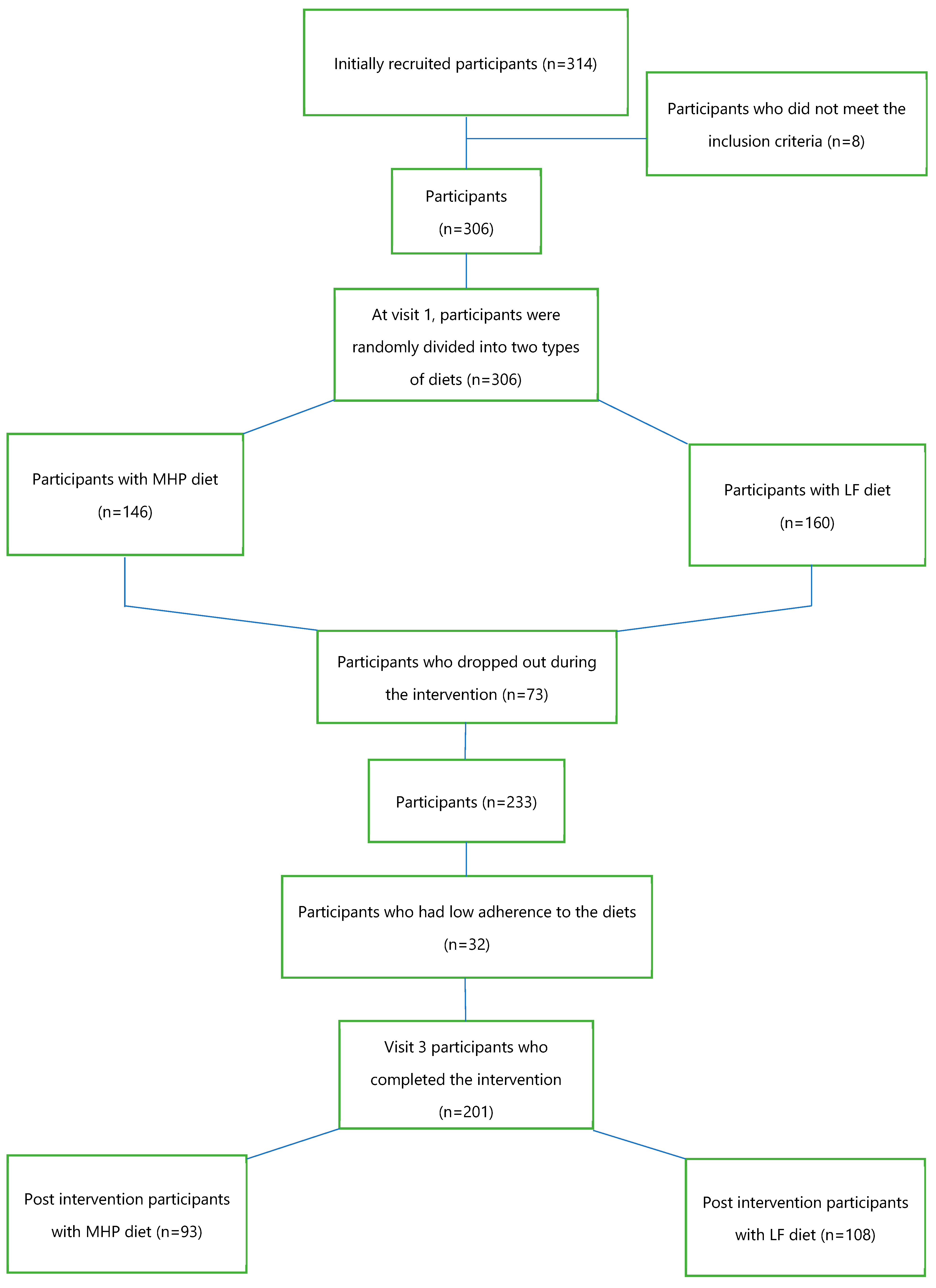
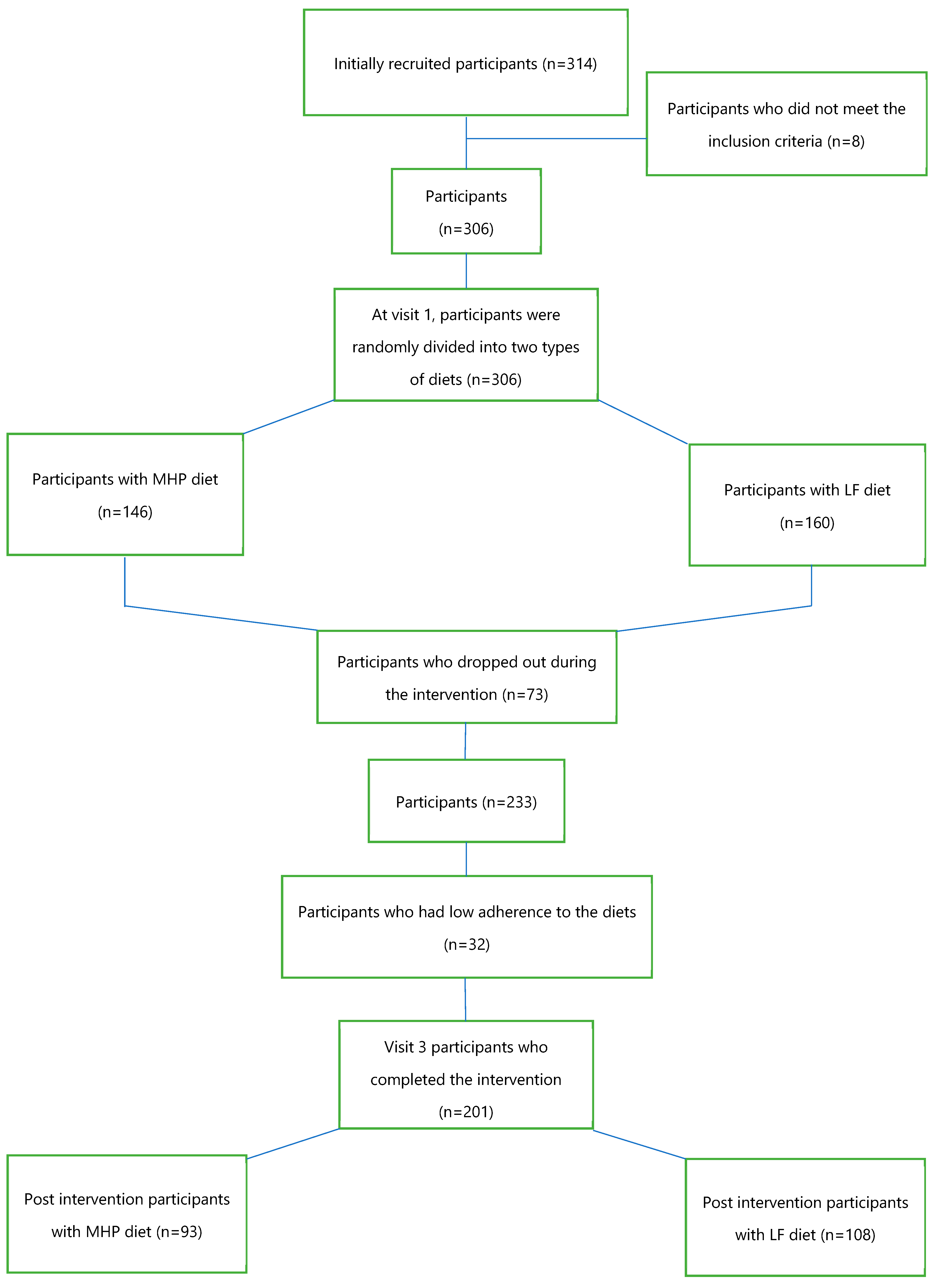
2.1. Study Population
2.2. Study Design
2.3. Nutritional Intervention
2.4. Anthropometric and Biochemical Determinations
2.5. DNA Isolation and Bisulfite Conversion
2.6. Array Analysis
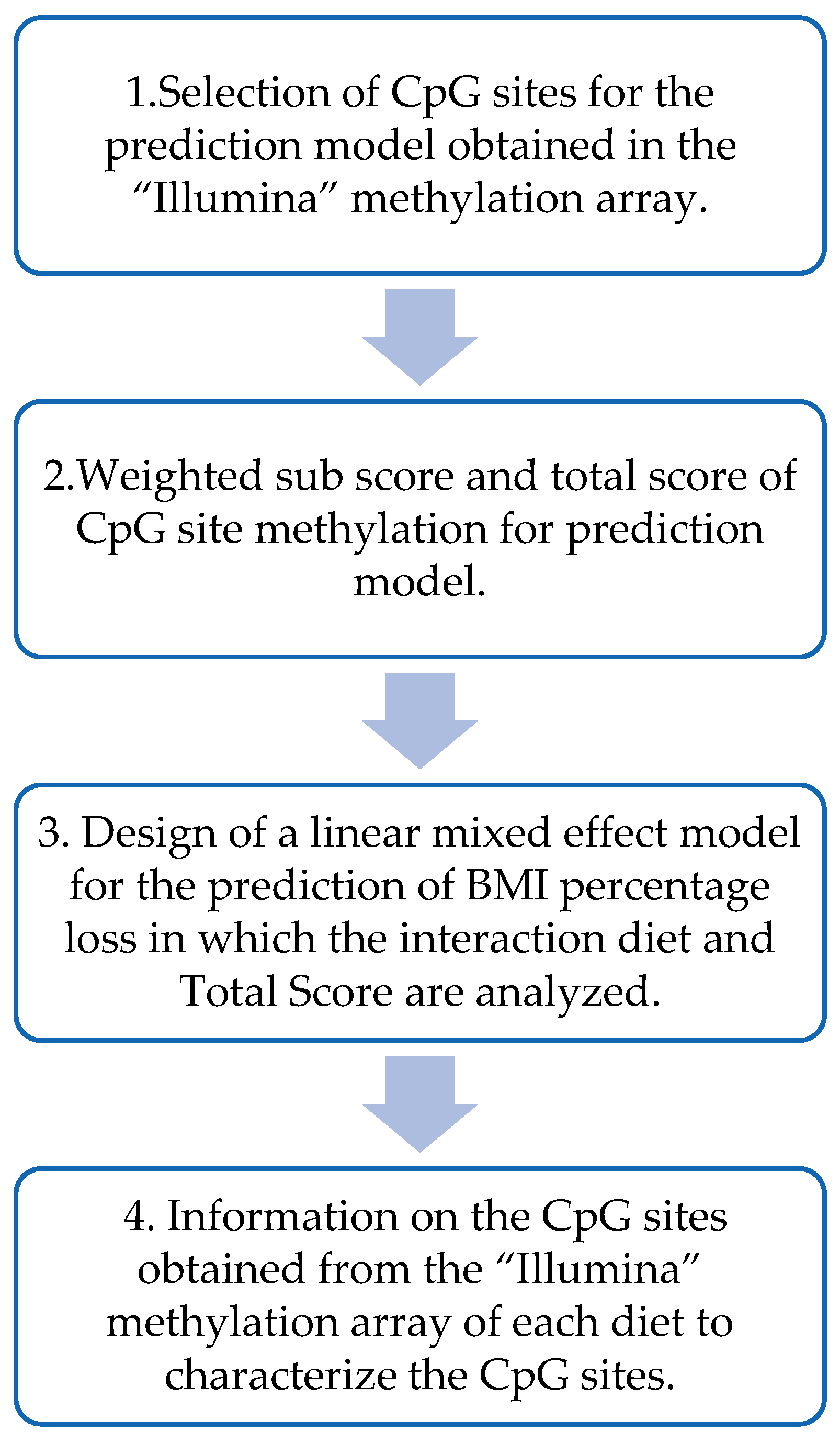
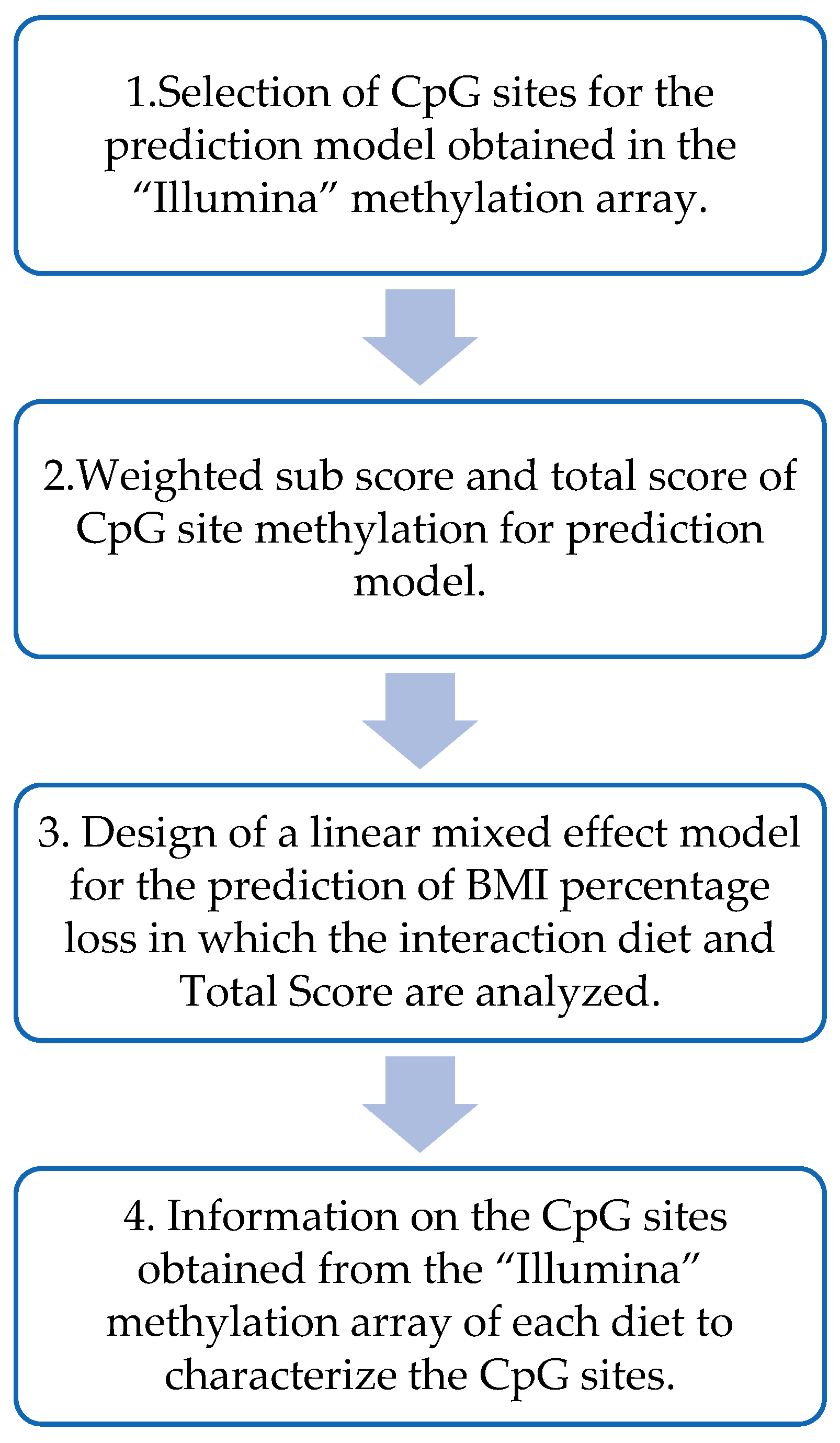
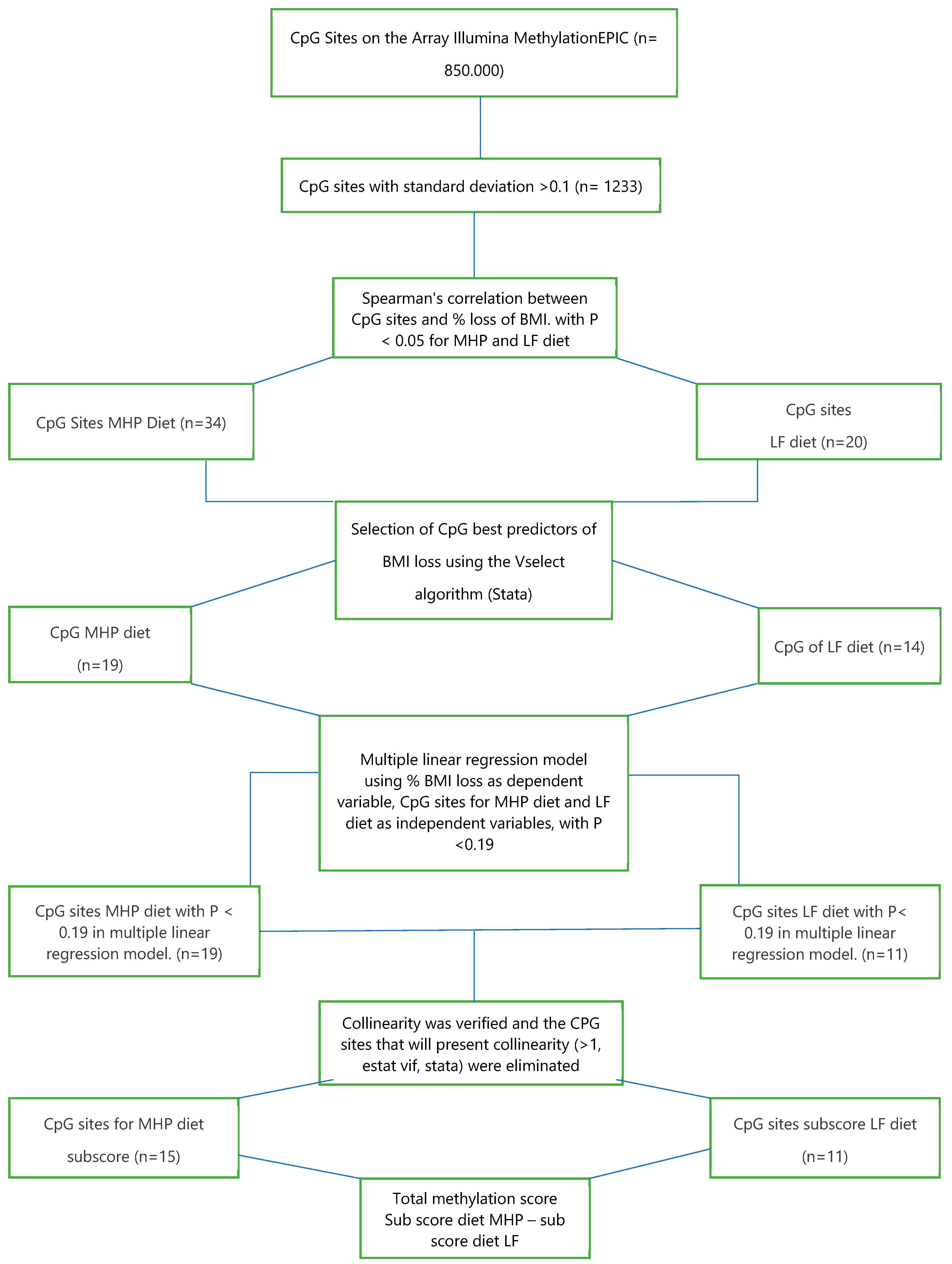
2.7. Design of the BMI Percentage Loss Prediction Model Based on the MHP and LF Diets’ Methylation Data
2.8. Statistical Analysis
2.9. Statistical Analysis for the Prediction Model
3. Results
3.1. Anthropometric and Biochemical Data at Baseline
3.2. Anthropometric and Biochemical Values after the Dietary Intervention and BMI Loss Prediction Model for the MHP and LF Diets Based on DNA Methylation Data
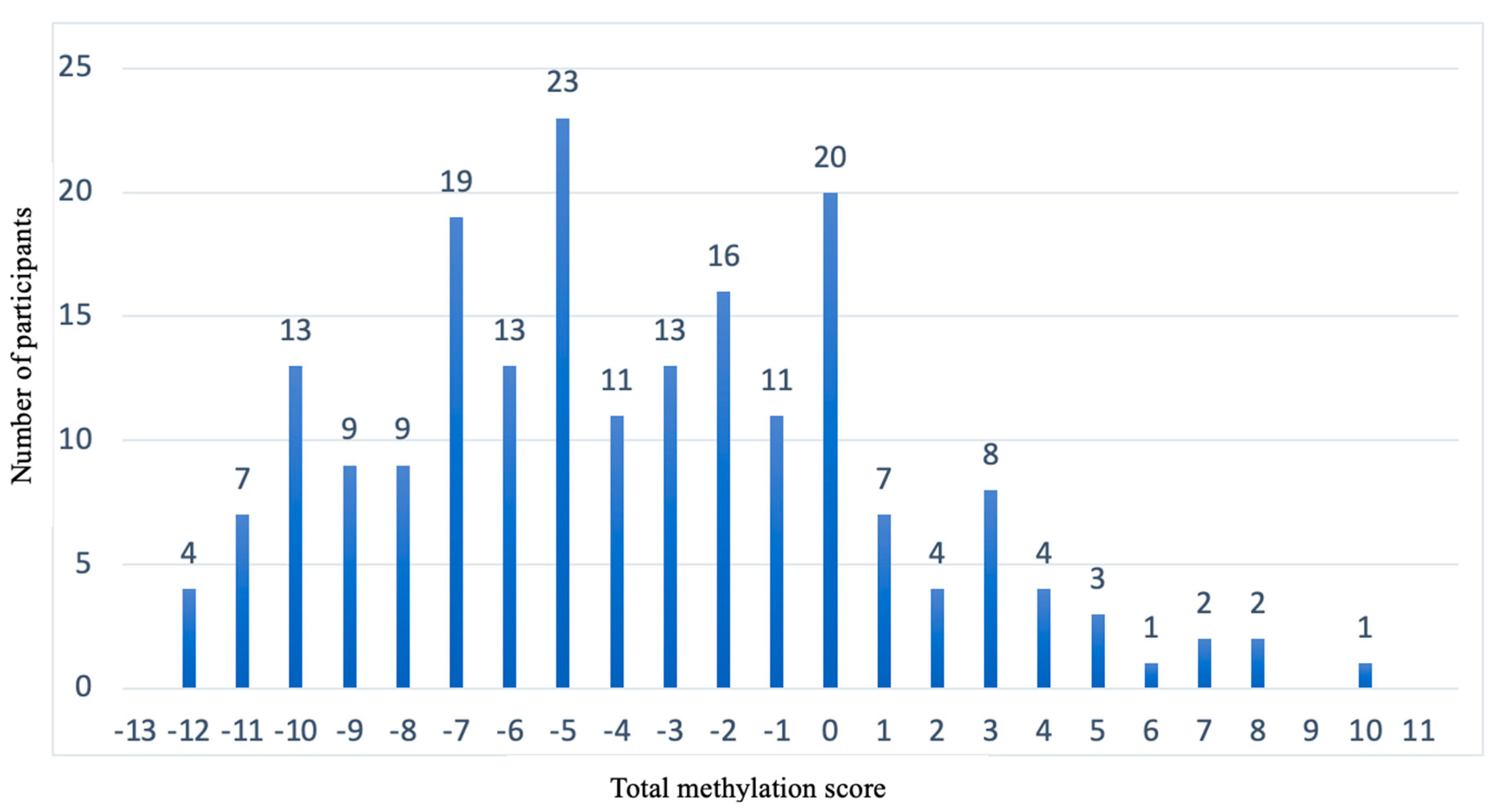
3.3. Design of Weighted Sub-Scores That Contain the CpG Sites of Each Diet and the Calculation of the Total Methylation Score for the Prediction Model
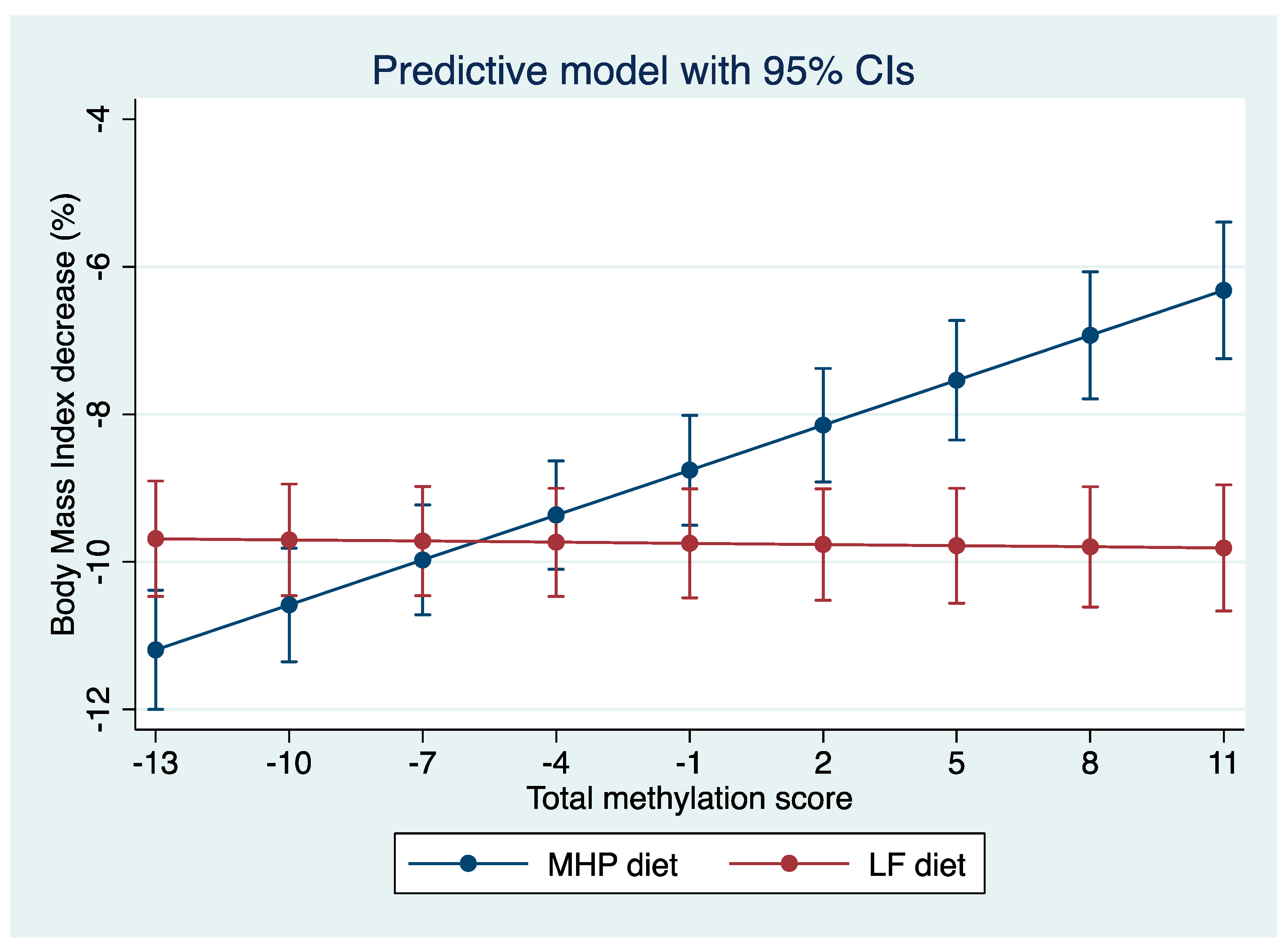
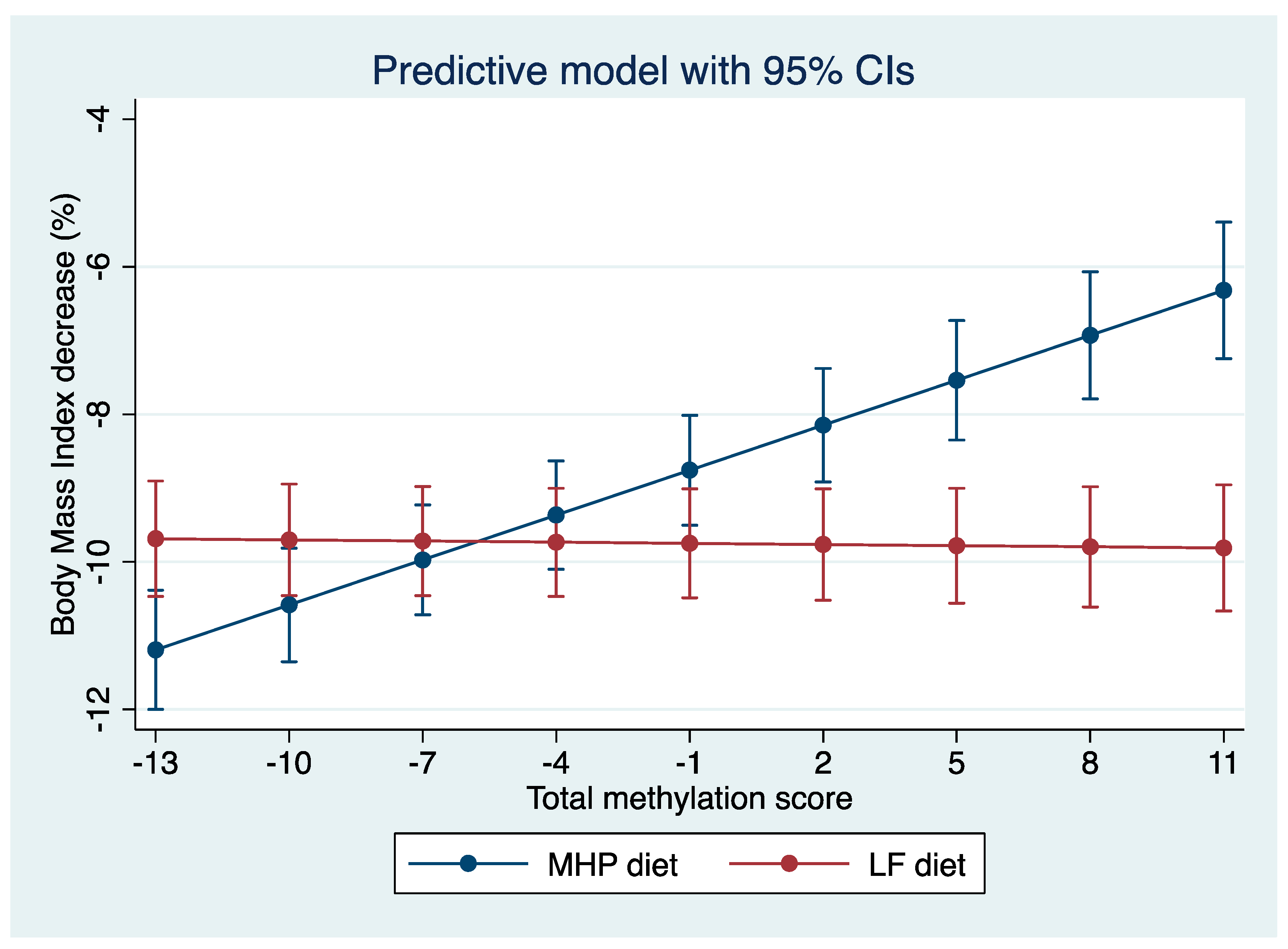
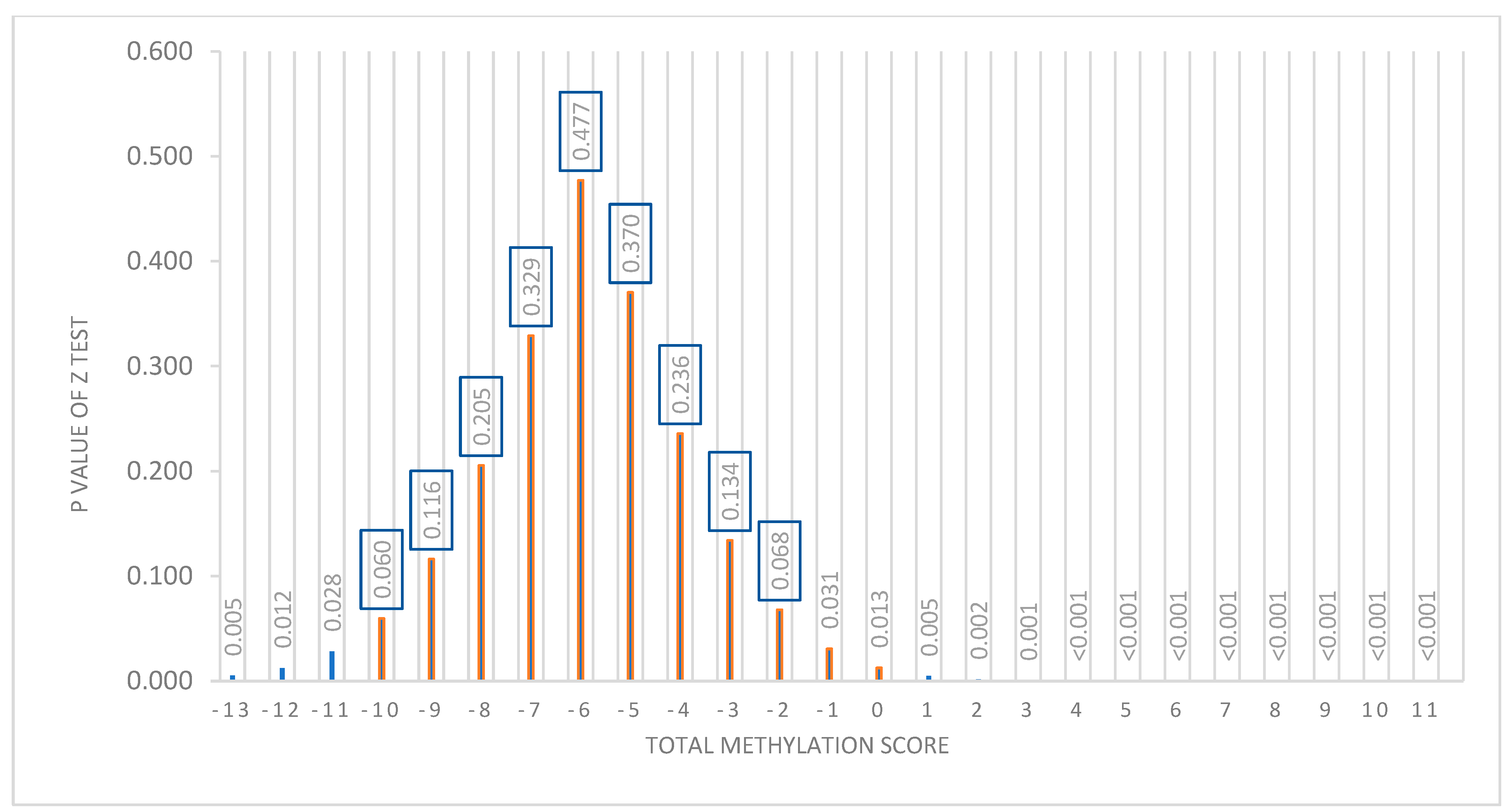
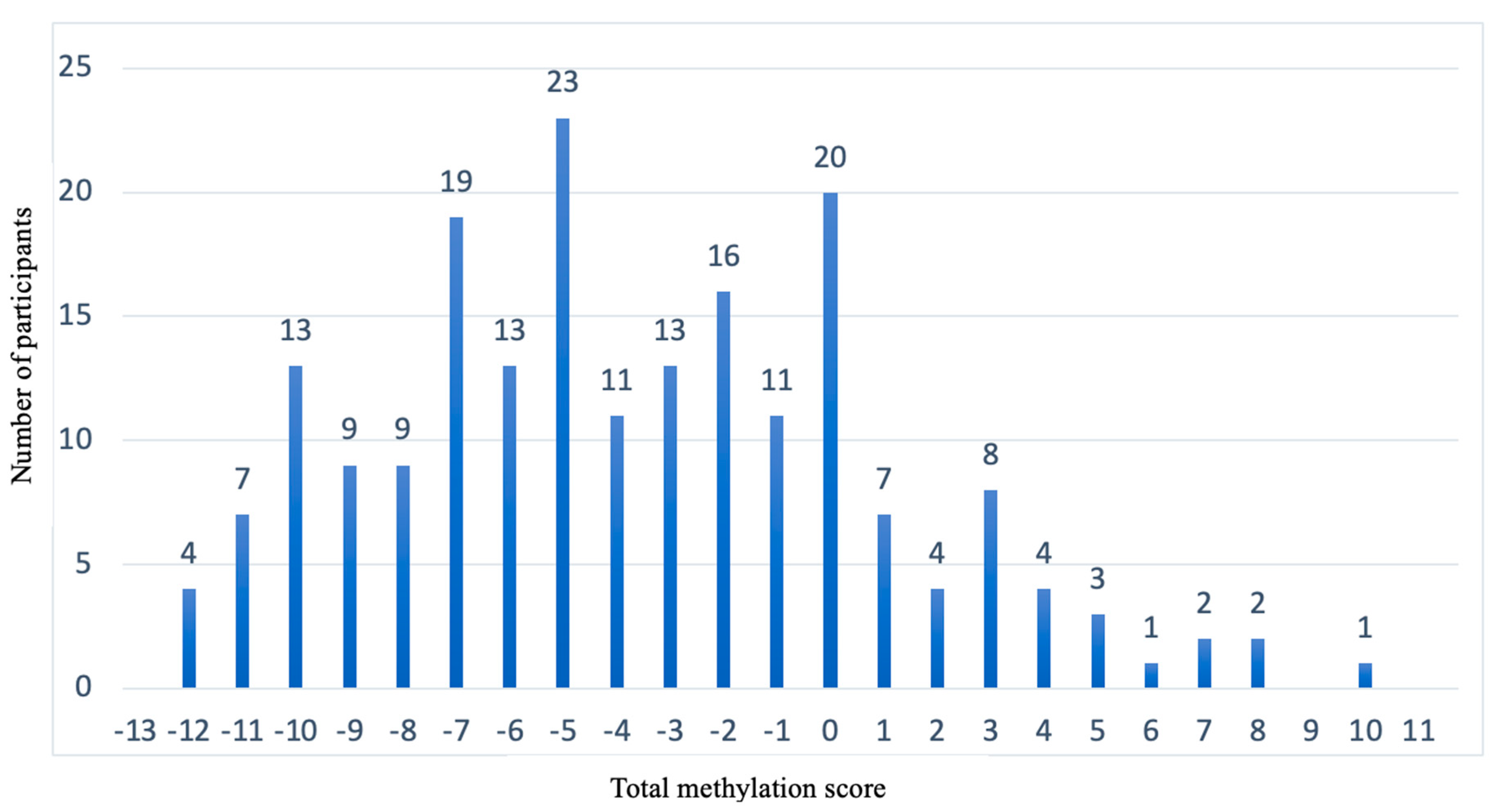
3.4. Representation of the Prediction Model
3.5. Information on the Methylation Sites Selected for the Prediction Model
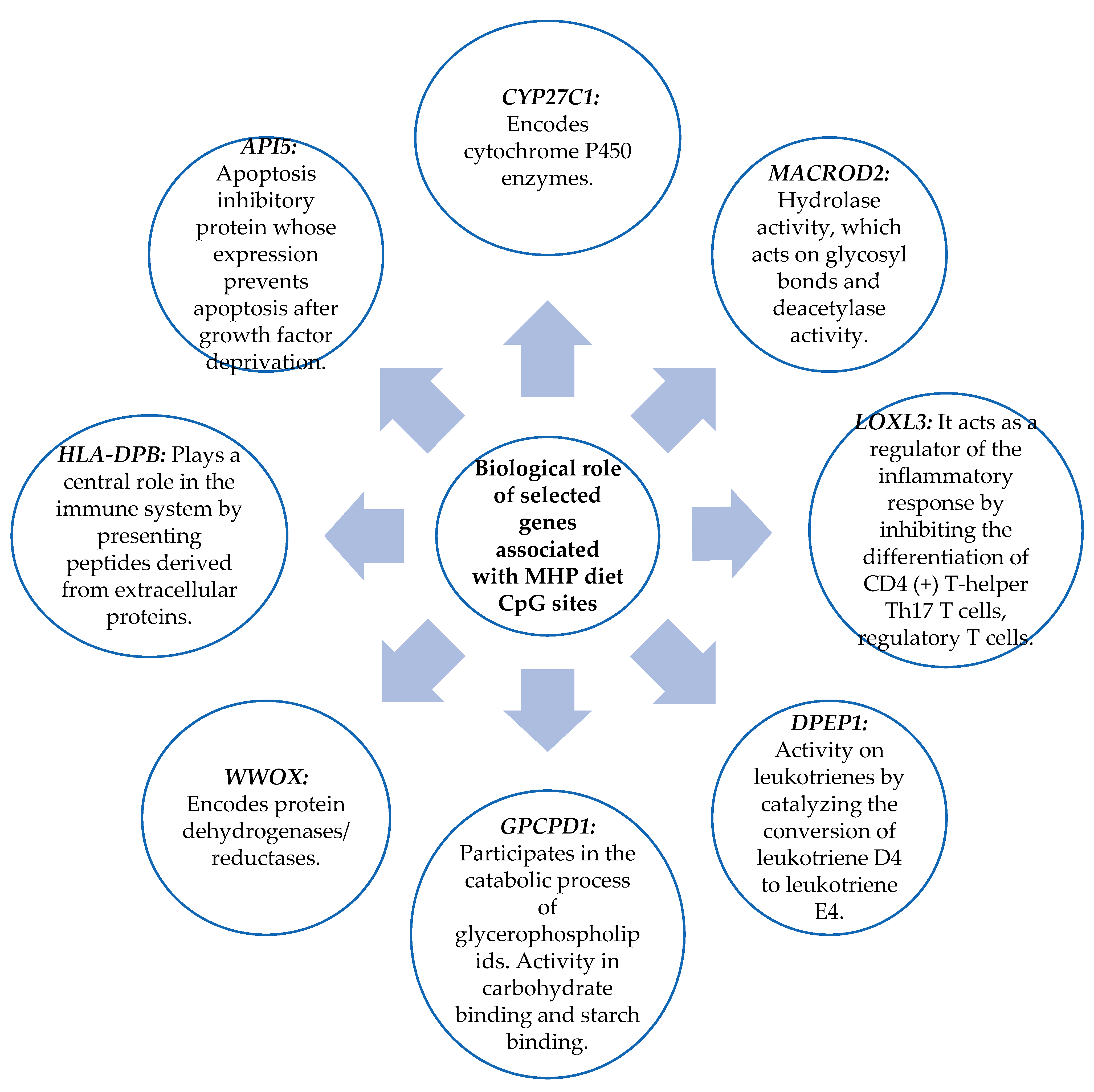
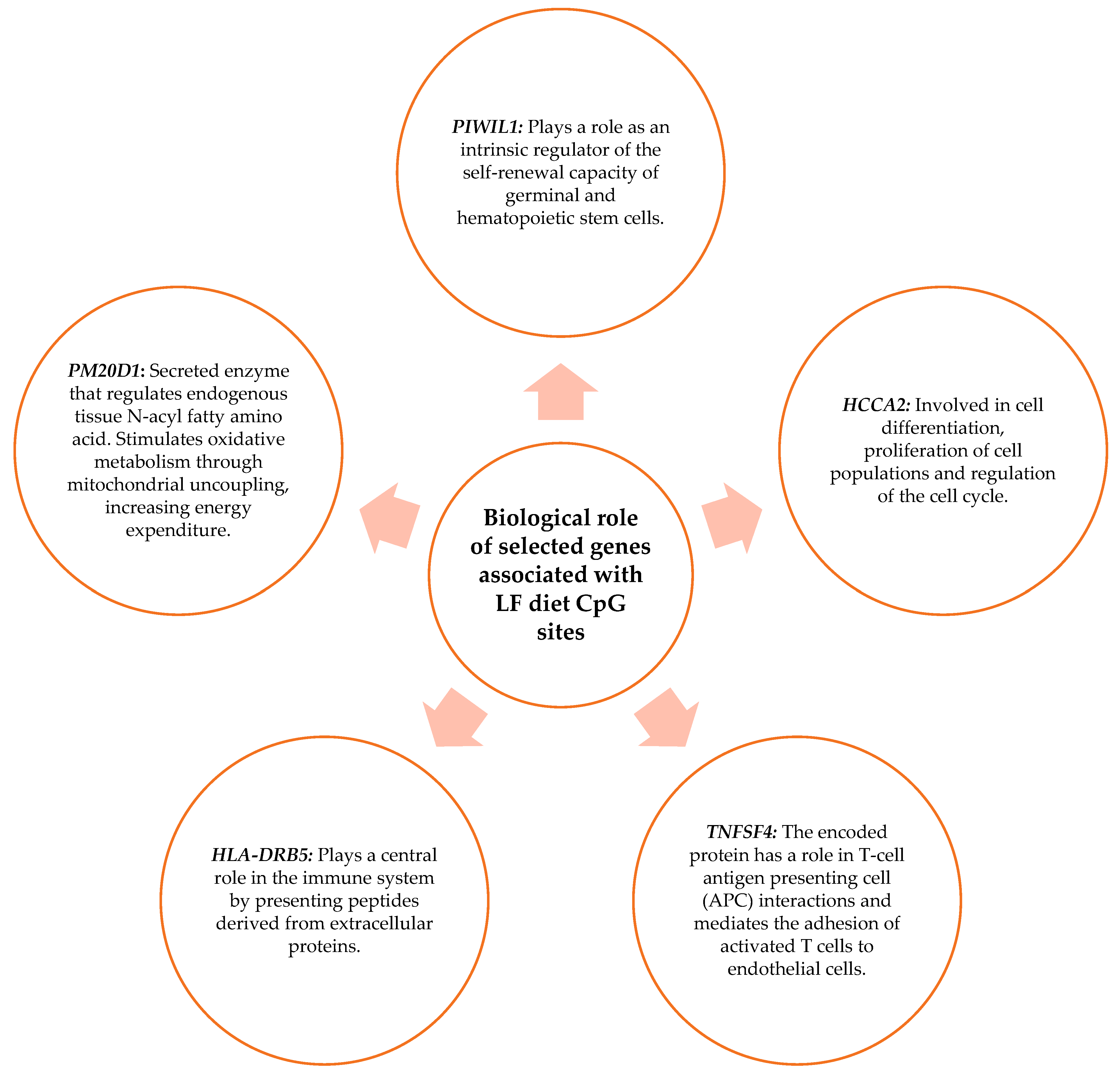
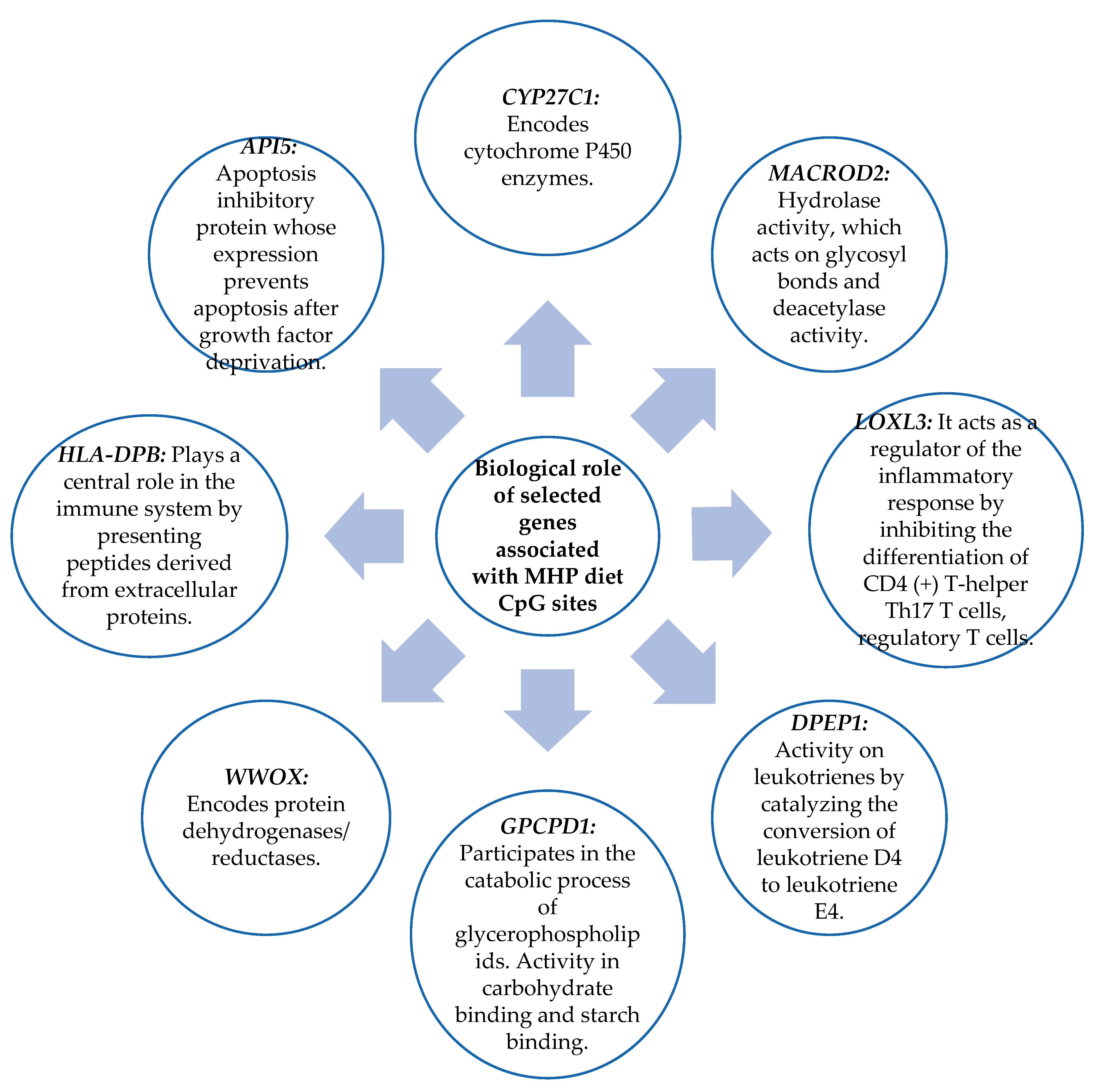
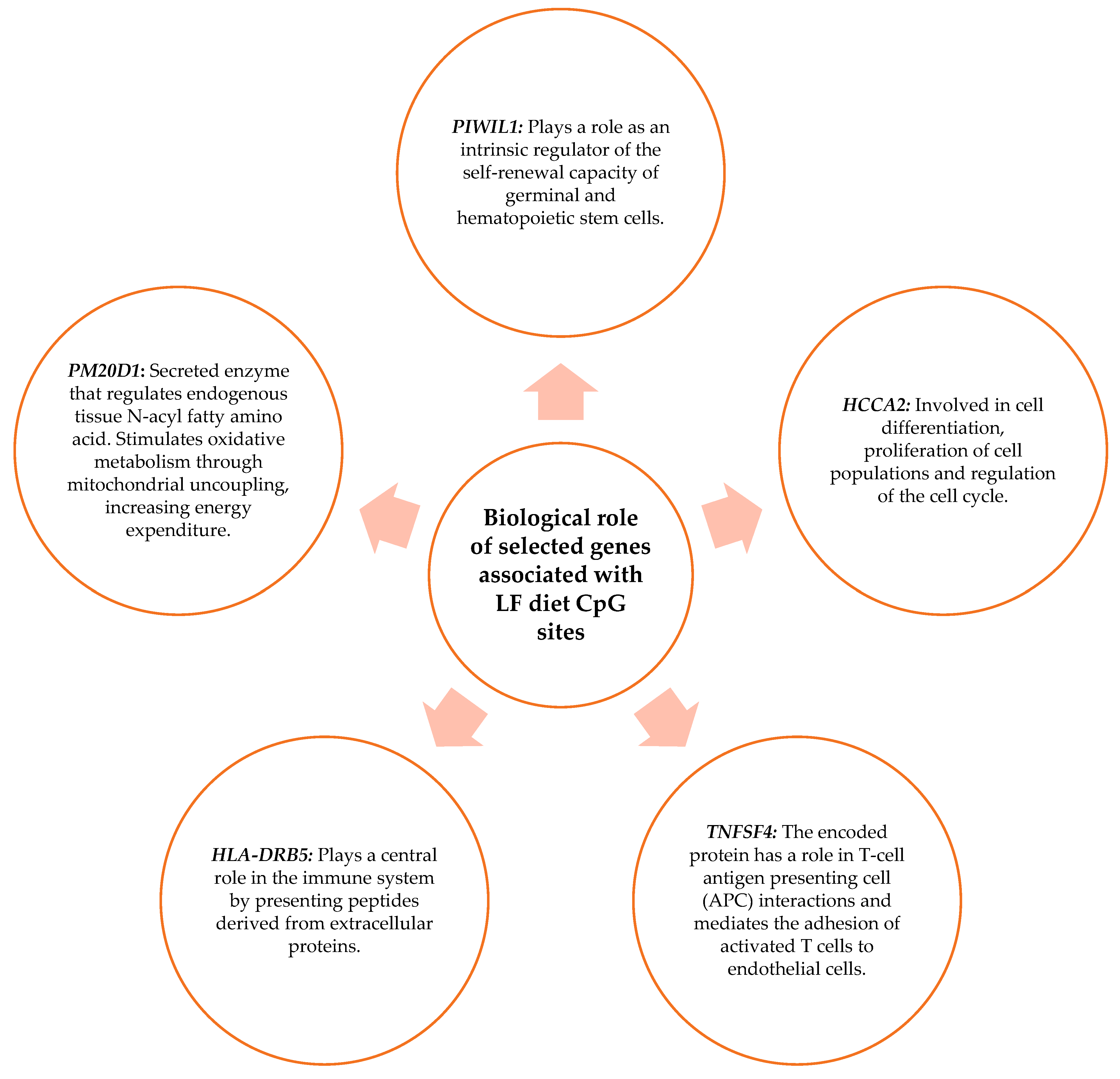
3.6. Biological Role of the Genes Related to the CpG Sites Selected for the Prediction Model
4. Discussion
4.1. Methylation Analyzed in Blood Samples Showing Association with the BMI
4.2. Genes Related to CpG Sites Associated with the Percentage of BMI Loss for the MHP Diet and for the LF Diet
4.3. Prediction Model
4.4. BMI Percentage Loss Prediction Model Based on the MHP and LF Diets’ Methylation Data
4.5. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 23 August 2023).
- Jung, B.C.; Kang, S. Epigenetic regulation of inflammatory factors in adipose tissue. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 159019. [Google Scholar] [CrossRef]
- Li, Y. Modern epigenetics methods in biological research. Methods 2021, 187, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.-Y.; Yin, R.-X. Recent progress in epigenetics of obesity. Diabetol. Metab. Syndr. 2022, 14, 171. [Google Scholar] [CrossRef]
- Matusheski, N.V.; Caffrey, A.; Christensen, L.; Mezgec, S.; Surendran, S.; Hjorth, M.F.; McNulty, H.; Pentieva, K.; Roager, H.M.; Seljak, B.K.; et al. Diets, nutrients, genes and the microbiome: Recent advances in personalized nutrition. Br. J. Nutr. 2021, 126, 1489–1497. [Google Scholar] [CrossRef] [PubMed]
- de la Barreda, N.J. La epigenética. Sus mecanismos y significado en la regulación génica. Cuad. Bioética 2020, 31, 405–419. [Google Scholar]
- Sun, D.; Heianza, Y.; Li, X.; Shang, X.; Smith, S.R.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic, epigenetic and transcriptional variations at NFATC2IP locus with weight loss in response to diet interventions: The POUNDS Lost Trial. Diabetes Obes. Metab. 2018, 20, 2298–2303. [Google Scholar] [CrossRef] [PubMed]
- Samblas, M.; Milagro, F.I.; Martínez, A. DNA methylation markers in obesity, metabolic syndrome, and weight loss. Epigenetics 2019, 14, 421–444. [Google Scholar] [CrossRef] [PubMed]
- Fuso, A.; Raia, T.; Orticello, M.; Lucarelli, M. The complex interplay between DNA methylation and miRNAs in gene expression regulation. Biochimie 2020, 173, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Guo, G.; Bi, Z.; Liu, Y.; Zhao, Y.; Chen, N.; Wang, F.; Wang, Y.; Wang, X. m6A methylation modulates adipogenesis through JAK2-STAT3-C/EBPβ signaling. Biochim. Biophys. Acta BBA-Gene Regul. Mech. 2019, 1862, 796–806. [Google Scholar] [CrossRef]
- Xie, H.; Liu, X.; Zhou, Q.; Huang, T.; Zhang, L.; Gao, J.; Wang, Y.; Liu, Y.; Yan, T.; Zhang, S.; et al. DNA Methylation Modulates Aging Process in Adipocytes. Aging Dis. 2022, 13, 433–446. [Google Scholar] [CrossRef]
- Czogała, W.; Strojny, W.; Schab, M.; Grabowska, A.; Miklusiak, K.; Kowalczyk, W.; Łazarczyk, A.; Tomasik, P.; Skoczeń, S. FTO and PLAG1 Genes Expression and FTO Methylation Predict Changes in Circulating Levels of Adipokines and Gastrointestinal Peptides in Children. Nutrients 2021, 13, 3585. [Google Scholar] [CrossRef]
- Xiao, H.; Kang, S. The role of DNA methylation in thermogenic adipose biology. Epigenetics 2019, 14, 837–843. [Google Scholar] [CrossRef]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef]
- Mahmoud, A.M. An overview of epigenetics in obesity: Th role of lifestyle and therapeutic interventions. Int. J. Mol. Sci. 2022, 23, 1341. [Google Scholar] [CrossRef] [PubMed]
- Lecorguillé, M.; Teo, S.; Phillips, C.M. Maternal dietary quality and dietary inflammation associated with offspring growth, placental development, and DNA methylation. Nutrients 2021, 13, 3130. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, P.; Nettore, I.C.; Franchini, F.; Palatucci, G.; Muscogiuri, G.; Colao, A.; Macchia, P.E. Epigenome Modulation Induced by Ketogenic Diets. Nutrients 2022, 14, 3245. [Google Scholar] [CrossRef] [PubMed]
- Severin, R.; Sabbahi, A.; Mahmoud, A.M.; Arena, R.; Phillips, S.A. Precision Medicine in Weight Loss and Healthy Living. Prog. Cardiovasc. Dis. 2019, 62, 15–20. [Google Scholar] [CrossRef]
- Ramos-Lopez, O.; Riezu-Boj, J.I.; Milagro, F.I. Genetic and epigenetic nutritional interactions influencing obesity risk and adiposity outcomes. Curr. Opin. Clin. Nutr. Metab. Care 2022, 25, 235–240. [Google Scholar] [CrossRef]
- Reguero, M.; de Cedrón, M.G.; Wagner, S.; Reglero, G.; Quintela, J.C.; de Molina, A.R. Precision nutrition to activate thermogenesis as a complementary approach to target obesity and associated-meta-bolic-disorders. Cancers 2021, 13, 866. [Google Scholar] [CrossRef]
- Stover, P.J.; King, J.C. More Nutrition Precision, Better Decisions for the Health of Our Nation. J. Nutr. 2020, 150, 3058–3060. [Google Scholar] [CrossRef]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://www.who.int/data/gho/data/themes/topics/topic-details/GHO/body-mass-index (accessed on 23 August 2023).
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the Concentration of Low-Density Lipoprotein Cholesterol in Plasma, Without Use of the Preparative Ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef]
- Lindsey, C. Stat Softw Components. Boston College Department of Economics. VSELECT: Stata Module to Perform Linear Regression Variable Selection. Available online: https://ideas.repec.org/c/boc/bocode/s457808.html (accessed on 23 August 2023).
- Chen, Y.; Kassam, I.; Lau, S.H.; Kooner, J.S.; Wilson, R.; Peters, A.; Winkelmann, J.; Chambers, J.C.; Chow, V.T.; Khor, C.C.; et al. Impact of BMI and waist circumference on epigenome-wide DNA methylation and identification of epigenetic biomarkers in blood: An EWAS in multi-ethnic Asian individuals. Clin. Epigenetics 2021, 13, 195. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, T.; Su, S.; Hao, G.; Chen, T.; Li, Q.Z.; Bazzano, L.; He, J.; Wang, X.; Li, S.; et al. Body mass index drives changes in DNA methylation: A longitudinal study. Circ. Res. 2019, 125, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-L.; Lin, Z.-J.; Li, C.-C.; Lin, X.; Shan, S.-K.; Guo, B.; Zheng, M.-H.; Li, F.; Yuan, L.-Q.; Li, Z.-H. Epigenetic regulation in metabolic diseases: Mechanisms and advances in clinical study. Signal Transduct. Target. Ther. 2023, 8, 98. [Google Scholar] [CrossRef]
- Salas-Perez, F.; Assmann, T.S.; Ramos-Lopez, O.; Martínez, J.A.; Riezu-Boj, J.I.; Milagro, F.I. Crosstalk between Gut Microbiota and Epigenetic Markers in Obesity Development: Relationship between Ruminococcus, BMI, and MACROD2/SEL1L2 Methylation. Nutrients 2023, 15, 1550. [Google Scholar] [CrossRef] [PubMed]
- Dłuski, D.F.; Wolińska, E.; Skrzypczak, M. Epigenetic changes in gestational diabetes mellitus. Int. J. Mol. Sci. 2021, 22, 7649. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Q.; Yu, P.; Miao, W.; Liu, C.; Pu, Y.; Zhang, C. Methylation of WWOX gene promotes proliferation of osteosarcoma cells. J. Buon. 2020, 25, 2708–2713. [Google Scholar]
- Muhammad, J.S.; Eladl, M.A.; Khoder, G. Helicobacter pylori-induced DNA Methylation as an Epigenetic Modulator of Gastric Cancer: Recent Outcomes and Future Direction. Pathogens 2019, 8, 23. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Diaz-Lagares, A.; Sandoval, J.; Milagro, F.I.; Navas-Carretero, S.; Carreira, M.C.; Gomez, A.; Hervas, D.; Monteiro, M.P.; Casanueva, F.F.; et al. DNA methylation map in circulating leukocytes mirrors subcutaneous adipose tissue methylation pattern: A genome-wide analysis from non-obese and obese patients. Sci. Rep. 2017, 7, 41903. [Google Scholar] [CrossRef]
- Nabais, M.F.; Gadd, D.A.; Hannon, E.; Mill, J.; McRae, A.F.; Wray, N.R. An overview of DNA methylation-derived trait score methods and applications. Genome Biol. 2023, 24, 28. [Google Scholar] [CrossRef]
- Thibault, R.; Abbasoglu, O.; Ioannou, E.; Meija, L.; Ottens-Oussoren, K.; Pichard, C.; Rothenberg, E.; Rubin, D.; Siljamäki-Ojansuu, U.; Vaillant, M.-F.; et al. ESPEN guideline on hospital nutrition. Clin. Nutr. 2021, 40, 5684–5709. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros Pomar, M.D.; Vilarrasa García, N.; Ángel, M.; Herrera, R.; Barahona, M.J.; Bueno, M.; Pedragós, A.C.I.; Continente, A.C.; Ciudin, A.; Carballido, F.C.; et al. Abordaje clínico integral SEEN de la obesidad en la edad adulta. Endocrinol. Diabetes Nutr. 2021, 68, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Do, W.L.; Sun, D.; Meeks, K.; Dugué, P.A.; Demerath, E.; Guan, W.; Li, S.; Chen, W.; Milne, R.; Adeyemo, A.; et al. Epigenome-wide meta-analysis of BMI in nine cohorts: Examining the utility of epigenetically predicted BMI. Am. J. Hum. Genet. 2023, 110, 273–283. [Google Scholar] [CrossRef]
- Jian, C.; Silvestre, M.P.; Middleton, D.; Korpela, K.; Jalo, E.; Broderick, D.; de Vos, W.M.; Fogelholm, M.; Taylor, M.W.; Raben, A.; et al. Gut microbiota predicts body fat change following a low-energy diet: A PREVIEW intervention study. Genome Med. 2022, 14, 54. [Google Scholar] [CrossRef] [PubMed]
- de Luis, D.A.; Izaola, O.; Primo, D.; Gómez, J.J.L. Role of beta-2 adrenergic receptor polymorphism (rs1042714) on body weight and glucose metabolism response to a meal-replacement hypocaloric diet. Nutrition 2023, 116, 112170. [Google Scholar] [CrossRef]
- Crovesy, L.; Rosado, E.L. Interaction between genes involved in energy intake regulation and diet in obesity. Nutrition 2019, 67–68, 110547. [Google Scholar] [CrossRef]
- Ramos-Lopez, O.; Cuervo, M.; Goni, L.; Milagro, F.I.; Riezu-Boj, J.I.; Martinez, J.A. Modeling of an integrative prototype based on genetic, phenotypic, and environmental information for personalized prescription of energy-restricted diets in overweight/obese subjects. Am. J. Clin. Nutr. 2020, 111, 459–470. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MHP (n = 93) | LF (n = 108) | p | |
|---|---|---|---|
| Gender (male/female) n (% male) | 28 (30%) | 33 (31%) | 0.945 |
| Age | 52 ± 1 | 54 ± 1 | 0.277 |
| BMI (kg/m2) | 31.2 ± 0.3 | 32.1 ± 0.3 | 0.062 |
| Body weight (kg) | 86.6 ± 1.4 | 88.7 ± 1.1 | 0.256 |
| Waist circumference (cm) | 101 ± 1 | 102 ± 0 | 0.463 |
| Hip circumference (cm) | 111 ± 1 | 112 ± 1 | 0.225 |
| Lean mass dxa (g) | 47585 ± 1009 | 48646 ± 939 | 0.442 |
| Fat mass dxa (g) | 36190 ± 734 | 37250 ± 752 | 0.317 |
| Visceral fat mass dxa (g) | 1362 ± 91 | 1486 ± 79 | 0.301 |
| Diastolic pressure (mmHg) | 78. ± 1 | 79 ± 1 | 0.399 |
| Systolic pressure (mmHg) | 130 ± 2 | 128 ± 1 | 0.511 |
| Total energy (Kcal) | 1509 ± 24.1 | 1514 ± 19.5 | 0.863 |
| Glucose (mmol/L) | 5 ± 0.5 | 5 ± 0.1 | 0.317 |
| Insulin (mU/l) | 7.7 ± 0.5 | 8.1 ± 0.4 | 0.703 |
| Leptin (ng/mL) | 34.2 ± 2.4 | 38.1 ± 2.9 | 0.325 |
| Adiponectin (μg/mL) | 11.1 ± 0.4 | 11.5 ± 0.4 | 0.577 |
| HOMA-IR | 1.8 ± 0.1 | 1.9 ± 0.1 | 0.726 |
| Cholesterol (mg/dL) | 214 ± 3 | 218 ± 3 | 0.534 |
| HDL-cholesterol (mg/dL) | 53 ± 1 | 55 ± 1 | 0.301 |
| Triglycerides (mg/dL) | 98 ± 4 | 103 ± 5 | 0.505 |
| LDL-c (mg/dL) | 1400 ± 3 | 141 ± 3 | 0.911 |
| ox-LDL (mg/dL) | 44 ± 1 | 46 ± 1 | 0.303 |
| Alt (IU/L) | 24.3 ± 1.7 | 23.1 ± 1.1 | 0.506 |
| Ast (IU/L) | 22.5 ± 1.2 | 21.5 ± 0.6 | 0.468 |
| Uric acid (mg/dL) | 5.1 ± 0.1 | 5.2 ± 0.1 | 0.433 |
| C- Reactive protein (mg/L) | 2.7 ± 0.2 | 3.1 ± 0.3 | 0.551 |
| TNF-α (pg/mL) | 0.9 ± 0.3 | 0.8 ± 0.3 | 0.006 |
| MPH Diet (n = 93) | LF Diet (n = 108) | Comparison of the Differences between the MHP Diet and the LF Diet | ||||
|---|---|---|---|---|---|---|
| Mean ± SEM | p 1 | Mean ±SEM | p 2 | Mean ± SEM | p 3 | |
| Δ BMI (kg/m2) | −3.1 ± 0.1 | <0.001 | −3.2 ± 0.1 | <0.001 | 0.2 ± 0.1 | 0.261 |
| Δ Body weight (kg) | −8.4 ± 0.3 | <0.001 | −9.1 ± 0.3 | <0.001 | 0.5 ± 0.5 | 0.272 |
| Δ Waist circumference (cm) | −8.9 ± 0.4 | <0.001 | −9.4 ± 0.3 | <0.001 | 0.5 ± 0.6 | 0.351 |
| Δ Hip circumference (cm) | −6.2 ± 0.3 | <0.001 | −6.7 ± 0.3 | <0.001 | 0.5 ± 0.5 | 0.295 |
| Δ Lean mass dxa (g) | −1221 ± 130 | <0.001 | −1662 ± 140 | <0.001 | 441 ± 193 | 0.023 |
| Δ Fat mass dxa (g) | −6841 ± 357 | <0.001 | −7103 ± 299 | <0.001 | 262 ± 463 | 0.572 |
| Δ Visceral fat mass dxa (g) | −454 ± 42 | <0.001 | −487 ± 36 | <0.001 | 33 ± 55 | 0.555 |
| Δ Diastolic pressure (mmHg) | −3.7 ± 0.9 | <0.001 | −4.1 ± 1.1 | <0.001 | 0.2 ± 1.4 | 0.863 |
| Δ Systolic pressure (mmHg) | −12.3 ± 1.3 | <0.001 | −10.1 ± 1.1 | <0.001 | −2.1 ± 1.7 | 0.213 |
| Δ Total energy (Kcal) | 415 ± 16 | <0.001 | 428 ± 11 | <0.001 | −13 ± 21 | 0.521 |
| Δ Glucose(mmol/L) | −0.7 ± 0.5 | <0.001 | −0.2 ± 0.4 | <0.001 | −0.5 ± 0.5 | 0.296 |
| Δ Insulin (mU/l) | −2.8 ± 0.4 | <0.001 | −2.6 ± 0.3 | <0.001 | −0.2 ± 0.6 | 0.694 |
| Δ Leptin (ng/mL) | −16.2 ± 2.1 | <0.001 | −19.1 ± 1.9 | <0.001 | 2.9 ± 2.8 | 0.301 |
| Δ Adiponectin (μg/mL) | 0.4 ± 0.2 | 0.047 | 0.1 ± 0.2 | 0.748 | 0.3 ± 0.3 | 0.311 |
| Δ HOMA-IR | −0.7 ± 0.1 | <0.001 | −0.7 ± 0.1 | <0.001 | −0.6 ± 0.1 | 0.701 |
| Δ Cholesterol (mg/dL) | −17.8 ± 2.5 | <0.001 | −22.1 ± 2.5 | <0.001 | 4.2 ± 3.6 | 0.247 |
| Δ HDL-c (mg/dL) | −2.7 ± 0.7 | 0.001 | −4.8 ± 0.8 | <0.001 | 2.1 ± 1.1 | 0.059 |
| Δ Triglycerides (mg/dL) | −19.7 ± 4.1 | <0.001 | −15.1 ± 3.6 | <0.001 | −4.6 ± 5.4 | 0.389 |
| Δ LDL-c (mg/dL) | −11.1 ± 2.1 | <0.001 | −14.2 ± 1.9 | <0.001 | 3.1 ± 2.8 | 0.281 |
| Δ ox-LDL (mg/dL) | −8.1 ± 0.9 | <0.001 | −8.6 ± 1.2 | <0.001 | 0.5 ± 1.6 | 0.722 |
| Δ Alt (IU/L) | −3.9 ± 1.8 | 0.003 | −3.6 ± 0.9 | <0.001 | −0.3 ± 1.9 | 0.867 |
| Δ Ast (IU/L) | −0.6 ± 1.6 | 0.686 | −1.0 ± 0.6 | 0.074 | 0.4 ± 1.6 | 0.787 |
| Δ Uric acid (mg/dL) | −0.1 ± 0.1 | 0.013 | −0.2 ± 0.1 | <0.001 | 0.1 ± 0.1 | 0.087 |
| Δ C- Reactive protein (mg/L) | −0.9 ± 0.2 | <0.001 | −1.0 ± 0.2 | <0.001 | 0.1 ± 0.3 | 0.738 |
| Δ TNF-α(pg/mL) | 0.03 ± 0.01 | 0.079 | 0.013 ± 0.02 | 0.511 | −0.04 ± 0.02 | 0.098 |
| CpG Sites | Annotated Gene | Rho | p |
|---|---|---|---|
| cg00124993 | MIR886 | −0.212 | 0.041 |
| cg00308130 | −0.218 | 0.035 | |
| cg01097406 | −0.238 | 0.022 | |
| cg03447554 | −0.243 | 0.019 | |
| cg04481923 | MIR886 | −0.229 | 0.027 |
| cg06478886 | −0.212 | 0.041 | |
| cg06536614 | MIR886 | −0.211 | 0.042 |
| cg07104639 | MACROD2 | 0.207 | 0.046 |
| cg07782112 | −0.208 | 0.045 | |
| cg08745965 | MIR886 | −0.222 | 0.032 |
| cg09768983 | 0.224 | 0.031 | |
| cg10841563 | −0.233 | 0.024 | |
| cg11460778 | 0.217 | 0.036 | |
| cg14317533 | −0.214 | 0.039 | |
| cg15263617 | GPCPD1 | 0.239 | 0.021 |
| cg15837280 | −0.265 | 0.010 | |
| cg17052675 | −0.245 | 0.018 | |
| cg17764313 | MCM2;MCM2 | −0.237 | 0.022 |
| cg18678645 | MIR886 | −0.207 | 0.046 |
| cg18797653 | MIR886 | −0.210 | 0.043 |
| cg19053046 | HLA-DPB1 | 0.221 | 0.033 |
| cg19148731 | LOXL3 | −0.214 | 0.039 |
| cg19504605 | ZFP41 | −0.204 | 0.049 |
| cg20315590 | HMCN1 | −0.215 | 0.038 |
| cg20684491 | 0.301 | 0.003 | |
| cg21054447 | 0.207 | 0.046 | |
| cg22355889 | ELMOD1;LOC643923;ELMOD1 | 0.207 | 0.046 |
| cg23377942 | WWOX;WWOX | −0.206 | 0.048 |
| cg23899408 | HOOK2;HOOK2 | −0.224 | 0.031 |
| cg24433124 | −0.223 | 0.031 | |
| cg24658778 | SYNE1;SYNE1 | −0.269 | 0.009 |
| cg25340688 | MIR886 | −0.227 | 0.028 |
| cg26896946 | MIR886 | −0.239 | 0.021 |
| cg27149073 | SDHAP3 | 0.222 | 0.032 |
| Sitios CpG | Annotated Genes | Rho | p |
|---|---|---|---|
| cg00481382 | NEDD1;NEDD1;NEDD1;NEDD1 | 0.250 | 0.009 |
| cg03188948 | −0.192 | 0.046 | |
| cg04346459 | NFYA;NFYA;LOC221442 | 0.204 | 0.034 |
| cg07167872 | PM20D1 | 0.196 | 0.042 |
| cg11193064 | SMAD6;SMAD6 | −0.196 | 0.042 |
| cg14050976 | 0.203 | 0.035 | |
| cg14222729 | −0.236 | 0.014 | |
| cg14893161 | PM20D1;PM20D1 | 0.194 | 0.043 |
| cg15011943 | HLA-DRB5 | 0.205 | 0.033 |
| cg15572235 | 0.234 | 0.014 | |
| cg15695738 | 0.190 | 0.049 | |
| cg15837280 | 0.190 | 0.048 | |
| cg16078649 | RNF39;RNF39 | −0.262 | 0.006 |
| cg16600909 | 0.220 | 0.022 | |
| cg17035276 | −0.216 | 0.024 | |
| cg18493115 | HCCA2;KRTAP5-4 | −0.231 | 0.016 |
| cg19424457 | PIWIL1;PIWIL1 | −0.210 | 0.029 |
| cg20057198 | −0.251 | 0.009 | |
| cg24433124 | 0.220 | 0.022 | |
| cg26967960 | CAV3;CAV3 | 0.287 | 0.003 |
| MHP Diet (n = 93 Participants) | |||
|---|---|---|---|
| CpG Sites | Beta Coefficient | SEM | p |
| cg22355889 | 9.818 | 2.3 | <0.001 |
| cg24433124 | −5.813 | 1.7 | 0.002 |
| cg14317533 | −7.578 | 2.3 | 0.002 |
| cg07104639 | 6.259 | 2.14 | 0.005 |
| cg19148731 | −7.880 | 2.2 | 0.001 |
| cg01097406 | −5.982 | 1.6 | 0.001 |
| cg15263617 | 8.975 | 2.8 | 0.002 |
| cg23377942 | −4.191 | 2.3 | 0.075 |
| cg25340688 | −5.795 | 1.8 | 0.003 |
| cg19053046 | 5.080 | 2.6 | 0.055 |
| cg11460778 | 4.222 | 1.7 | 0.016 |
| cg03447554 | −4.080 | 1.9 | 0.035 |
| cg09768983 | 3.989 | 2.5 | 0.125 |
| cg20684491 | 4.803 | 1.9 | 0.015 |
| cg20315590 | −6.713 | 2.8 | 0.022 |
| LF Diet (n = 108 Participants) | |||
|---|---|---|---|
| CpG Sites | Beta Coefficient | SEM | p |
| cg15572235 | 7.570 | 2.8 | 0.010 |
| cg19424457 | −8.048 | 3.1 | 0.011 |
| cg18493115 | −6.267 | 3.1 | 0.048 |
| cg00481382 | 5.789 | 2.8 | 0.047 |
| cg16600909 | 5.380 | 3.5 | 0.129 |
| cg15837280 | 5.830 | 3.0 | 0.058 |
| cg03188948 | −4.308 | 2.4 | 0.081 |
| cg16078649 | −4.163 | 2.6 | 0.121 |
| cg15011943 | 4.649 | 3.3 | 0.169 |
| cg07167872 | 3.986 | 1.7 | 0.027 |
| cg14222729 | −3.407 | 2.5 | 0.187 |
| Score | Calculating Formula |
|---|---|
| Sub-score MHP Diet | (cg22355889 * 9.82) + (cg24433124 * −5.81) + (cg14317533 * − 7.58) + (cg07104639 * 6.25) + (cg19148731 * −7.88) + (cg01097406 * −5.98) + (cg15263617 * 8.97) + (cg23377942 * −4.19) + (cg25340688 * −5.79) + (cg19053046 * 5.08) + (cg11460778 * 4.22) + (cg03447554 * −4.08) + (cg09768983 * 3.98) + (cg20684491 * 4.80) + (cg20315590 * −6.71) |
| Sub-score LF Diet | (cg15572235 * 7.57) + (cg19424457 * −8.04) + (cg18493115 * −6.26) + (cg00481382 * 5.78) + (cg16600909 * 5.38) + (cg15837280 * 5.83) + (cg03188948 * −4.30) + (cg16078649 * −4.16) + (cg15011943 * 4.64) + (cg07167872 * 3.98) + (cg14222729 * −3.40) |
| Total Methylation Score | Sub-score MHP − Sub-Score LF |
| Prob > Chi-Square ≤ 0.001 Participants (n = 201) | Percentage of BMI Loss | |
|---|---|---|
| Independent Variable | Beta Coefficient ± SEM | p-Values Z-Test |
| Age | −0.001 ± < 0.001 | 0.943 |
| Sex | −0.006 ± 0.1 | 0.954 |
| Diet (MHP or LF) | −1.201 ± 0.1 | <0.001 |
| Total methylation score | 0.202 ± 0.01 | <0.001 |
| Diet## total methylation score | −0.208 ± 0.02 | <0.001 |
| Cons | −7.952 ± 0.6 | <0.001 |
| CpG Sites | Chromosome | Map INFO | Gene 1 | Gene Region 2 |
|---|---|---|---|---|
| cg22355889 | 11 | 107461585 | ELMOD1 | TSS1500 |
| cg07104639 | 20 | 15125595 | MACROD2 | Body |
| cg19148731 | 2 | 74780229 | LOXL3 | 5′UTR |
| cg15263617 | 20 | 5574362 | GPCPD1 | Body |
| cg23377942 | 16 | 79042805 | WWOX | Body |
| cg25340688 | 5 | 135416398 | MIR886 | TSS200 |
| cg19053046 | 6 | 33048254 | HLA-DPB1 | Body |
| cg20315590 | 1 | 186003041 | HMCN1 | Body |
| cg20684491 | 1 | 25596433 | RSRP1 | IGR (2476) |
| cg11460778 | 1 | 145385299 | NBPF10 | IGR (24,744) |
| cg03447554 | 11 | 43094025 | API5 | IGR (239,480) |
| cg09768983 | 4 | 183935060 | DCTD | IGR (143,816) |
| cg01097406 | 16 | 89675127 | DPEP1 | IGR (−4589) |
| cg24433124 | 6 | 30755968 | LINC00243 | IGR (−24,675) |
| cg14317533 | 2 | 127886316 | CYP27C1 | IGR (−55,096) |
| CpG Sites | Chromosome | Map INFO | Gene 1 | Gene Region 2 |
|---|---|---|---|---|
| cg19424457 | 12 | 130822308 | PIWIL1 | TSS200 |
| cg18493115 | 11 | 1643842 | HCCA2 | Body |
| cg00481382 | 12 | 97304412 | NEDD1 | 5′UTR |
| cg16078649 | 6 | 30039466 | RNF39 | Body |
| cg15011943 | 6 | 32493917 | HLA-DRB5 | Body |
| cg07167872 | 1 | 205819463 | PM20D1 | TSS200 |
| cg15572235 | 7 | 5183992 | RBAK | IGR (98,540) |
| cg16600909 | 1 | 173145001 | TNFSF4 | IGR (−7869) |
| cg15837280 | 5 | 135415258 | TGFBI | IGR (50,674) |
| cg03188948 | 7 | 1209495 | ZFAND2A-DT | IGR (9367) |
| cg14222729 | 2 | 731215 | TMEM18 | IGR (63,242) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Álvarez, N.C.; Riezu-Boj, J.I.; Martínez, J.A.; García-Calzón, S.; Milagro, F.I. A Predictive Tool Based on DNA Methylation Data for Personalized Weight Loss through Different Dietary Strategies: A Pilot Study. Nutrients 2023, 15, 5023. https://doi.org/10.3390/nu15245023
García-Álvarez NC, Riezu-Boj JI, Martínez JA, García-Calzón S, Milagro FI. A Predictive Tool Based on DNA Methylation Data for Personalized Weight Loss through Different Dietary Strategies: A Pilot Study. Nutrients. 2023; 15(24):5023. https://doi.org/10.3390/nu15245023
Chicago/Turabian StyleGarcía-Álvarez, Nereyda Carolina, José Ignacio Riezu-Boj, J. Alfredo Martínez, Sonia García-Calzón, and Fermín I. Milagro. 2023. "A Predictive Tool Based on DNA Methylation Data for Personalized Weight Loss through Different Dietary Strategies: A Pilot Study" Nutrients 15, no. 24: 5023. https://doi.org/10.3390/nu15245023
APA StyleGarcía-Álvarez, N. C., Riezu-Boj, J. I., Martínez, J. A., García-Calzón, S., & Milagro, F. I. (2023). A Predictive Tool Based on DNA Methylation Data for Personalized Weight Loss through Different Dietary Strategies: A Pilot Study. Nutrients, 15(24), 5023. https://doi.org/10.3390/nu15245023