
Evaluation of Prebiotic Activity of Stellariae Radix Polysaccharides and Its Effects on Gut Microbiota

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation Process of CSRPs
2.3. Evaluation of Prebiotic Activity In Vitro
2.4. Evaluation of Prebiotic Activity In Vivo
2.5. Composition and Diversity of Gut Microbiota
2.6. Statistical Analysis
3. Results
3.1. Composition of Polysaccharide from Stellariae Radix
3.2. Evaluation of Prebiotic Activity In Vitro
3.2.1. Fitting of Growth Curve
3.2.2. Effects of CSRPs on Growth of Lactobacillus Acidophilus and Bifidobacterium Longum
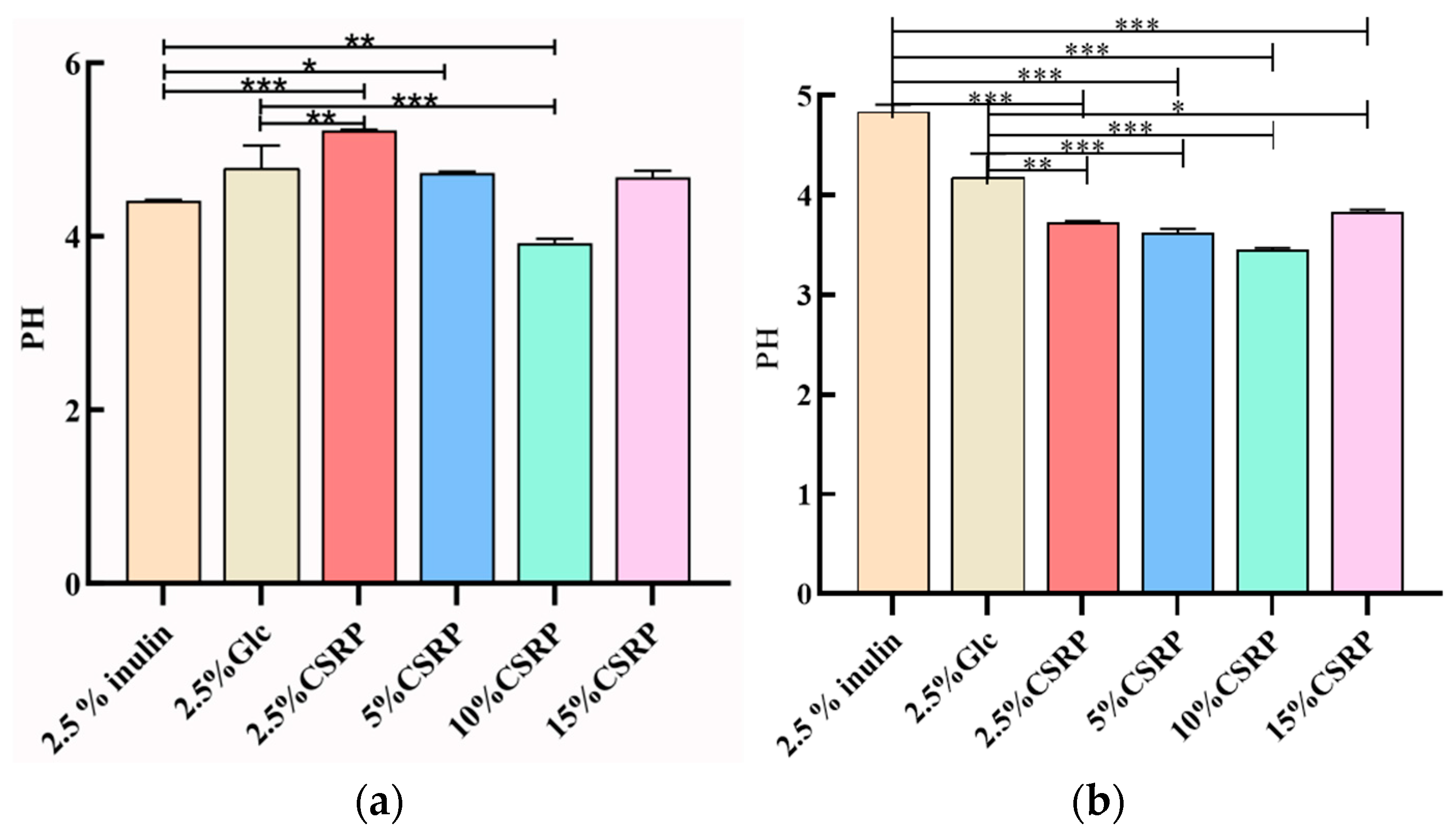
3.2.3. Effects of CSRPs on Acid Production of Two Probiotics
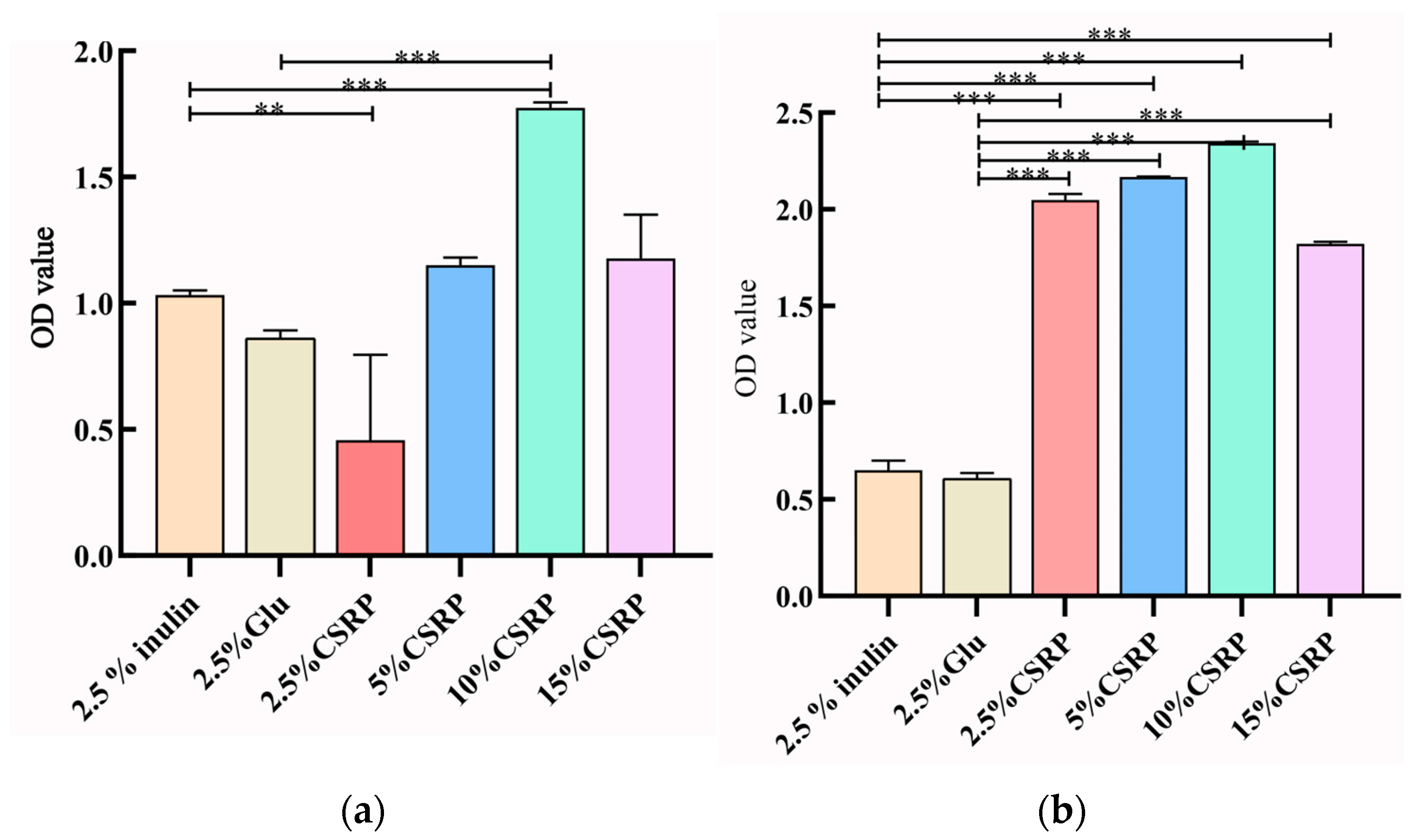
3.2.4. Effects of Different Concentrations of CSRPs on Growth Density of two Probiotic Strains
3.2.5. Probiotic Index Calculation of Different Concentrations of CSRPs on Two Probiotic Strains
3.3. Evaluation of Prebiotic Activity In Vivo
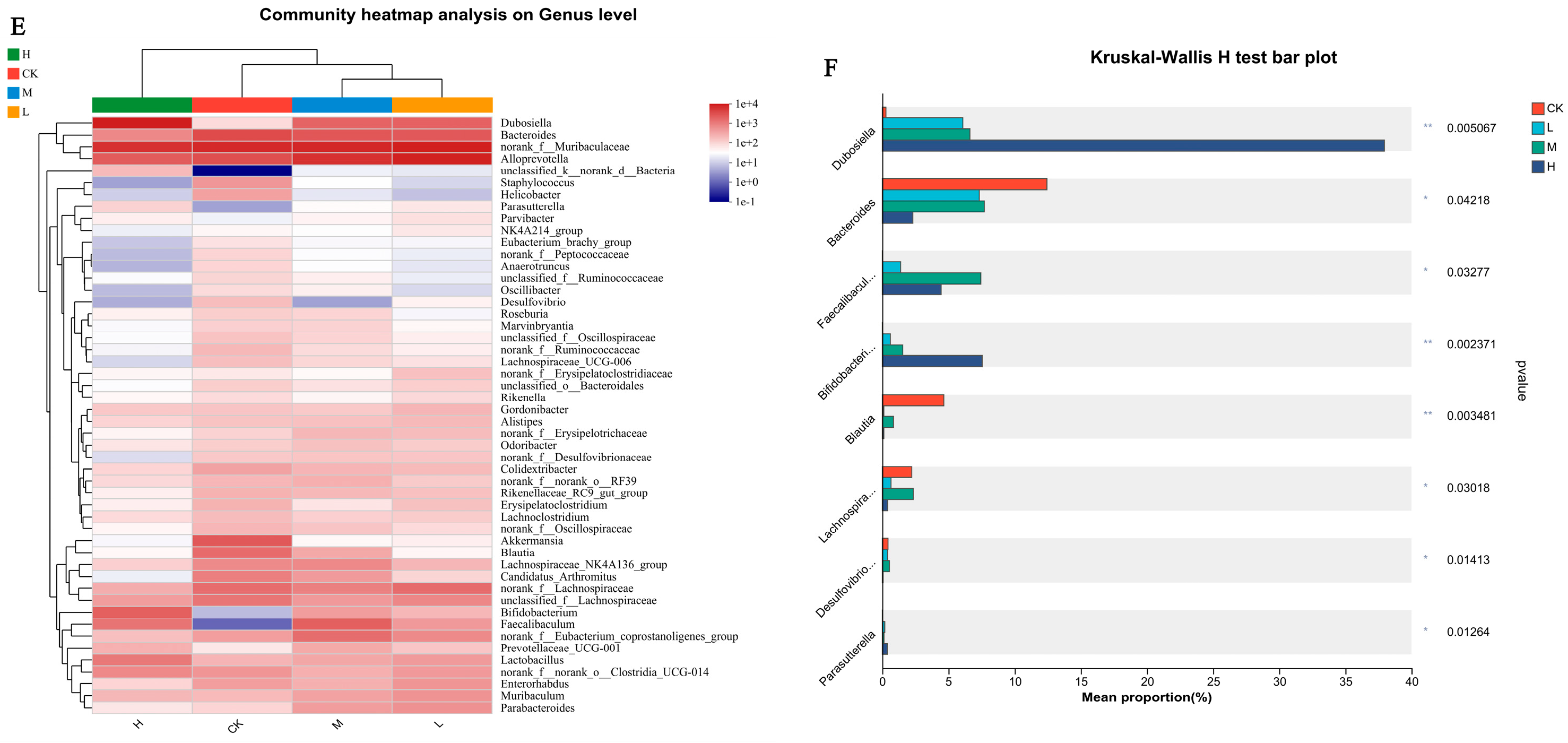
Effects of CSRP Intake on the Composition of Cecal Gut Microbiotas
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Altermann, E.; Russell, W.M.; Azcarate-Peril, M.A.; Barrangou, R.; Buck, B.L.; McAuliffe, O.; Souther, N.; Dobson, N.; Duong, T.; Callanan, M. Complete genome sequence of the probiotic lactic acidbacterium Lactobacillus acidophilus NCFM. Proc. Natl. Acad. Sci. USA 2005, 102, 3906–3912. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wu, Y.-J.; Sun, P.-L.; Zhang, F.-M.; Linhardt, R.-J.; Zhang, A.-Q. Chemically modified polysaccharides: Synthesis, characterization, structure activity relationships of action. Int. J. Biol. Macromol. 2019, 132, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lu, C.; Wu, C.; Xu, M.; Kou, X.; Kong, D.; Jing, G. Polysaccharide of Boschniakia rossica induces apoptosis on laryngeal carcinoma Hep2 cells. Gene 2014, 536, 203–206. [Google Scholar] [CrossRef]
- Yang, Y.; Ji, J.; Di, L.; Li, J.; Hu, L.; Qiao, H.; Wang, L.; Feng, Y. Resource, chemical structure and activity of natural polysaccharides against alcoholic liver damages. Carbohydr. Polym. 2020, 241, 116355. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.-Y.; Aweya, J.J.; Li, N.; We, H.; Sun, C.; Li, J.; Jiang, Y. Microbial catabolism of Porphyra haitanensis polysaccharides by human gut microbiota. Food Chem. 2019, 289, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Fang, Y.; Nishinari, K.; We, H.; Sun, C.; Li, J.; Jiang, Y. Physicochemical characteristics of polysaccharide conjugates prepared from fresh tea leaves and their improving impaired glucose tolerance. Carbohydr. Polym. 2014, 112, 77–84. [Google Scholar] [CrossRef]
- Chen, R.; Liu, B.; Wang, X.; Chen, K.; Zhang, K.; Zhang, L.; Fei, C.; Wang, C.; Liu, Y.; Xue, F. Effects of polysaccharide from Pueraria lobata on gut microbiota in mice. Int. J. Biol. Macromol. 2020, 158, 740–749. [Google Scholar] [CrossRef]
- Porter, N.T.; Martens, E.C. The Critical Roles of Polysaccharides in Gut Microbial Ecology and Physiology. Annu. Rev. Microbiol. 2017, 71, 349–369. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, S.; Liu, J.; McLean, R.; Chu, W. Prebiotic, immuno-stimulating and gut microbiota-modulating effects of Lycium barbarum polysaccharide. Biomed. Pharmacother. 2020, 121, 109591. [Google Scholar] [CrossRef]
- Xie, S.-Z.; Liu, B.; Ye, H.-Y.; Li, Q.-M.; Pan, L.-H.; Zha, X.-Q.; Liu, J.; Duan, J.; Luo, J.-P. Dendrobium huoshanense polysaccharide regionally regulates intestinal mucosal barrier function and intestinal microbiota in mice. Carbohydr. Polym. 2019, 206, 149–162. [Google Scholar] [CrossRef]
- Fang, W.P.; Zhang, Z.R. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 2004. [Google Scholar]
- China Pharmacopoeia; National Pharmacopoeia Committee: Beijing, China, 2020; pp. 330–331.
- Li, Z.K.; Wang, H.; Song, L.; Feng, L.; Li, Y.Q.; Yang, Y.; Peng, L. Origin Characteristics and Correlation Analysis of Stellarae Radix Based on Inorganic Elements and Effective Components. Chin. J. Mod. Appl. Pharm. 2023, 40, 894–902. [Google Scholar]
- Song, L.; Wang, H.; Li, Z.K.; Liu, D.H.; Peng, L. Optimization of Supercritical CO2 Extraction Process and Component Analysis of Extract from Stellariae Radix. Chin. J. Mod. Appl. Pharm. 2022, 39, 2498–2504. [Google Scholar]
- Li, Z.K.; Ma, L.; Bai, M.S.; Huang, Y.X.; Song, L.; Wang, H.; Feng, L.; Peng, L. Infrared fingerprint of Ningxia Stellariae Radix based on chemometrics and discrimination analysis of counterfeit products. Mod. Tradit. Chin. Med. Mater. Medica-World Sci. Technol. 2023, 25, 175–183. [Google Scholar]
- Wang, H.; Peng, L.; Song, L.; Feng, L.; Li, Z.K.; Li, Y.Q.; Gao, T. Optimization of Ultrasonic-assisted Extraction Process and Analysis of Antioxidant Activity of Polysaccharide from Stellariae Radix. Food Ind. Sci. Technol. 2023, 1–12. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Zhang, X.Y.; Wang, D.P.; Xu, X.X. The Effect of Polysaccharide of Poriacocos on Key Metabolites of Bifidobacterium BB-12. Sci. Technol. Food Ind. 2022, 43, 24–33. [Google Scholar]
- Zwietering, M.; Jongenburger, I.; Rombouts, F.; Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef]
- Su, J.; Su, L.; Li, D.; Shuai, O.; Zhang, Y.; Liang, H.; Jiao, C.; Xu, Z.; Lai, Y.; Xie, Y. Antitumor Activity of Extract from the Sporoderm-Breaking Spore of Ganoderma lucidum: Restoration on Exhausted Cytotoxic T Cell with Gut Microbiota Remodeling. Front. Immunol. 2018, 9, 1765. [Google Scholar] [CrossRef]
- Peng, L.; Wang, H.; Song, L.; Feng, L.; Niu, P.L.; Li, Z.K.; Li, Y.Q.; Li, H.S.; Wu, W. A Crude Polysaccharide of Stellariae Radix with Prebiotic Activity, and Its Preparation. Method. Patent CN115677874A, 3 February 2023. [Google Scholar]
- Liu, B.; Liu, T.T.; Zhang, X.K.; Wang, T.; Zhang, C.X.; Li, M.; Zheng, X.Y.; Wang, D.P.; Zhang, D.Z.; Meng, X.J. Research progress of prebiotics and their applications. Food Med. 2021, 23, 485–492. [Google Scholar]
- Huang, L.; Ke, H.; Chen, G.H.; Zheng, Y. Preparation and quality analysis of Polygonatum polysaccharide prebiotic cake. Food Ind. 2020, 41, 90–94. [Google Scholar]
- Zhang, S.; Dang, Y. Roles of gut microbiota and metabolites in overweight and obesity of children. Front. Endocrinol. 2022, 13, 994930. [Google Scholar] [CrossRef]
- Seo, K.H.; Kim, D.H.; Jeong, D. Chardonnay grape seed flour supplemented diets alter intestinal microbiota in diet-induced obese mice. J. Food Biochem. 2017, 41, e12396. [Google Scholar] [CrossRef]
- Zhu, S.L.; Chen, Y.Q.; Jiang, X. Application of Dunaliella in Preventing and Improving Obesity and Related. Diseases. Patent CN111686133A, 22 September 2020. [Google Scholar]
- Zhou, X.; Ma, L.; Dong, L.; Li, D.; Chen, F.; Hu, X. Bamboo shoot dietary fiber alleviates gut microbiota dysbiosis and modulates liver fatty acid metabolism in mice with high-fat diet-induced obesity. Front. Nutr. 2023, 10, 1161698. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Daza, M.C.; Roquim, M.; Dudonné, S.; Pilon, G.; Levy, E.; Marette, A.; Roy, D.; Desjardins, Y. Berry Polyphenols and Fibers Modulate Distinct Microbial Metabolic Functions and Gut Microbiota Enterotype-Like Clustering in Obese Mice. Front. Microbiol. 2020, 11, 2032. [Google Scholar] [CrossRef]
- Gao, J.; Ma, L.; Yin, J.; Liu, G.; Ma, J.; Xia, S.; Gong, S.; Han, Q.; Li, T.; Chen, Y. Camellia (Camellia oleifera bel.) seed oil reprograms gut microbiota and alleviates lipid accumulation in high fat-fed mice through the mTOR pathway. Food Funct. 2022, 13, 4977–4992. [Google Scholar] [CrossRef]
- Yang, K.; Jian, S.; Wen, C.; Guo, D.; Liao, P.; Wen, J.; Kuang, T.; Han, S.; Liu, Q.; Deng, B. Gallnut Tannic Acid Exerts Anti-stress Effects on Stress-Induced Inflammatory Response, Dysbiotic Gut Microbiota, and Alterations of Serum Metabolic Profile in Beagle Dogs. Front. Nutr. 2022, 9, 847966. [Google Scholar] [CrossRef]
- Qiu, X.; Macchietto, M.G.; Liu, X.; Lu, Y.; Ma, Y.; Guo, H.; Saqui-Salces, M.; Bernlohr, D.A.; Chen, C.; Shen, S.; et al. Identification of gut microbiota and microbial metabolites regulated by an antimicrobial peptide lipocalin 2 in high fat diet-induced obesity. Int. J. Obes. 2021, 45, 143–154. [Google Scholar] [CrossRef]
- Zhu, Z.; Huang, R.; Huang, A.; Wang, J.; Liu, W.; Wu, S.; Chen, M.; Chen, M.; Xie, Y.; Jiao, C.; et al. Polysaccharide from Agrocybe cylindracea prevents diet-induced obesity through inhibiting inflammation mediated by gut microbiota and associated metabolites. Int. J. Biol. Macromol. 2022, 209 Pt A, 1430–1438. [Google Scholar] [CrossRef]
- Deng, M.; Zhang, S.; Dong, L.; Huang, F.; Jia, X.; Su, D.; Chi, J.; Muhammad, Z.; Ma, Q.; Zhao, D.; et al. Shatianyu (Citrus grandis L. Osbecontrol) Flavonoids and Dietary Fiber in Combination Are More Effective Than Individually in Alleviating High-Fat-Diet-Induced Hyperlipidemia in Mice by Altering Gut Microbiota. J. Agric. Food Chem. 2022, 70, 14654–14664. [Google Scholar] [CrossRef] [PubMed]
- Poeker, S.A.; Geirnaert, A.; Berchtold, L.; Greppi, A.; Krych, L.; Steinert, R.E.; de Wouters, T.; Lacroix, C. Understanding the prebiotic potential of different dietary fibers using an in vitro continuous adult fermentation model (PolyFermS). Sci. Rep. 2018, 8, 4318. [Google Scholar] [CrossRef]
- Yadav, M.K.; Kumari, I.; Singh, B.; Sharma, K.K.; Tiwari, S.K. Probiotics, prebiotics and synbiotics: Safe options for next-generation therapeutics. Appl. Microbiol. Biotechnol. 2022, 106, 505–521. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Cai, G.; Liu, T.; Liu, Z. Relationships Among Gut Microbiota, Ischemic Stroke and Its Risk Factors: Based on Research Evidence. Int. J. Gen. Med. 2022, 15, 2003–2023. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Wang, Y.; Zeng, L.; Dong, J.; Bi, Q.; Yang, X.; Che, Y.; He, S.; Yu, J. Polysaccharides from Polygonatum kingianum improve Glccose and lipid metabolism in rats fed a high fat diet. Biomed. Pharmacother. 2020, 125, 109910. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| . | Parameters | A1 | A2 | X0 | p | R2 |
|---|---|---|---|---|---|---|
| Lactobacillus acidophilus | 2.5% inulin | 0.024 ± 0.06 | 0.727 ± 0.02 | 4.892 ± 0.55 | 3.951 ± 1.23 | 0.967 |
| 2.5% Glc | 0.024 ± 0.06 | 1.140 ± 0.03 | 5.668 ± 0.46 | 2.857 ± 0.54 | 0.985 | |
| 2.5% CSRP | 0.021 ± 0.03 | 0.954 ± 0.01 | 4.307 ± 0.16 | 4.035 ± 0.37 | 0.996 | |
| 5% CSRP | 0.032 ± 0.04 | 1.391 ± 0.02 | 5.244 ± 0.20 | 4.138 ± 0.54 | 0.996 | |
| 10% CSRP | 0.008 ± 0.02 | 1.872 ± 0.02 | 7.345 ± 0.12 | 3.792 ± 0.20 | 0.999 | |
| 15% CSRP | 0.009 ± 0.02 | 1.801 ± 0.02 | 8.495 ± 0.17 | 3.930 ± 0.27 | 0.998 | |
| Bifidobacterium longum | 2.5% inulin | 0.044 ± 0.04 | 1.923 ± 0.13 | 25.043 ± 1.34 | 4.058 ± 0.66 | 0.991 |
| 2.5% Glc | 0.027 ± 0.04 | 1.487 ± 0.04 | 20.010 ± 0.42 | 4.181 ± 0.36 | 0.997 | |
| 2.5% CSRP | 0.005 ± 0.04 | 2.192 ± 0.05 | 14.899 ± 0.31 | 6.441 ± 0.74 | 0.995 | |
| 5% CSRP | 0.020 ± 0.03 | 2.217 ± 0.02 | 11.330 ± 0.14 | 7.986 ± 0.67 | 0.998 | |
| 10% CSRP | 0.002 ± 0.05 | 2.360 ± 0.05 | 11.820 ± 0.26 | 6.017 ± 0.69 | 0.996 | |
| 15% CSRP | 0.016 ± 0.03 | 1.711 ± 0.05 | 17.966 ± 0.35 | 7.601 ± 0.96 | 0.990 |
| Bacterial Strain | Control Group (%) | Concentration of CSRP (%) | ||||
|---|---|---|---|---|---|---|
| 2.5% Inulin | 2.5% Glc | 2.5% | 5% | 10% | 15% | |
| Lactobacillus acidophilus | 1.36 ± 0.10 b | 0.95 ± 0.05 | 0.8 ± 0.17 a | 1.76 ± 0.02 c | 2.39 ± 0.03 e | 2.06 ± 0.02 d |
| Bifidobacterium longum | 1.49 ± 0.20 a | 0.84 ± 0.03 | 2.08 ± 0.03 b | 2.36 ± 0. 03 d | 2.44 ± 0.02 e | 2.27 ± 0.03 c |
| 2.5% CSRP | 5% CSRP | 10% CSRP | 15% CSRP | |
|---|---|---|---|---|
| Lactobacillus acidophilus | 0.85 ± 0.02 c | 1.28 ± 0.01 b | 1.73 ± 0.01 a | 1.72 ± 0.01 a |
| Bifidobacterium longum | 2.16 ± 0.03 b | 2.16 ± 0.01 b | 2.34 ± 0.02 a | 1.62 ± 0.07 c |
| Sample | Richness | Diversity | Coverage | ||
|---|---|---|---|---|---|
| Chao1 | Ace | Shannon | Simpson | ||
| CK | 288.26 ± 26.66 | 286.53 ± 27.36 | 3.40 ± 0.39 | 0.10 ± 0.04 | 0.999 |
| L | 312.61 ± 19.09 | 306.36 ± 21.03 | 3.38 ± 0.36 | 0.08 ± 0.02 | 0.999 |
| M | 316.24 ± 38.84 | 308.36 ± 39.18 | 3.38 ± 0.81 | 0.06 ± 0.36 | 0.999 |
| H | 284.81 ± 17.29 | 284.57 ± 17.29 | 2.84 ± 0.28 | 0.18 ± 0.05 | 0.998 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Li, H.; Li, Z.; Feng, L.; Peng, L. Evaluation of Prebiotic Activity of Stellariae Radix Polysaccharides and Its Effects on Gut Microbiota. Nutrients 2023, 15, 4843. https://doi.org/10.3390/nu15224843
Wang H, Li H, Li Z, Feng L, Peng L. Evaluation of Prebiotic Activity of Stellariae Radix Polysaccharides and Its Effects on Gut Microbiota. Nutrients. 2023; 15(22):4843. https://doi.org/10.3390/nu15224843
Chicago/Turabian StyleWang, Hong, Haishan Li, Zhenkai Li, Lu Feng, and Li Peng. 2023. "Evaluation of Prebiotic Activity of Stellariae Radix Polysaccharides and Its Effects on Gut Microbiota" Nutrients 15, no. 22: 4843. https://doi.org/10.3390/nu15224843
APA StyleWang, H., Li, H., Li, Z., Feng, L., & Peng, L. (2023). Evaluation of Prebiotic Activity of Stellariae Radix Polysaccharides and Its Effects on Gut Microbiota. Nutrients, 15(22), 4843. https://doi.org/10.3390/nu15224843