
Influence of Lactose Supplementation on Regulation of Streptococcus thermophilus on Gut Microbiota

 , , ,
, , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Experimental Design
2.3. 16S rRNA Gene Sequencing
2.4. Metabolite Analysis of Fecal and Serum Samples
2.5. Statistical Analyses
3. Results
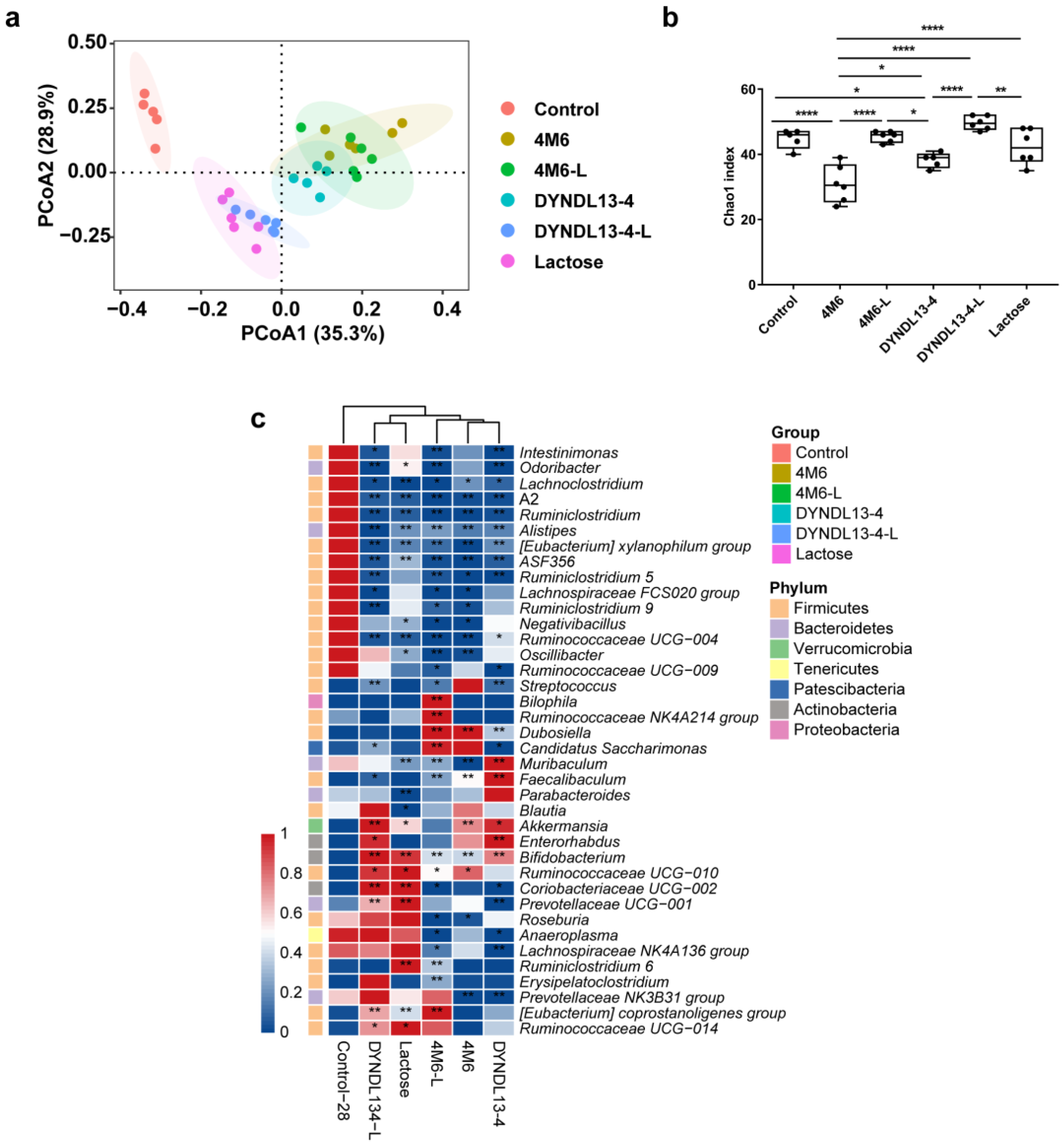
3.1. Effect of Lactose Supplementation on Gut Microbiota Induced by S. thermophilus Ingestion Is Strain-Specific
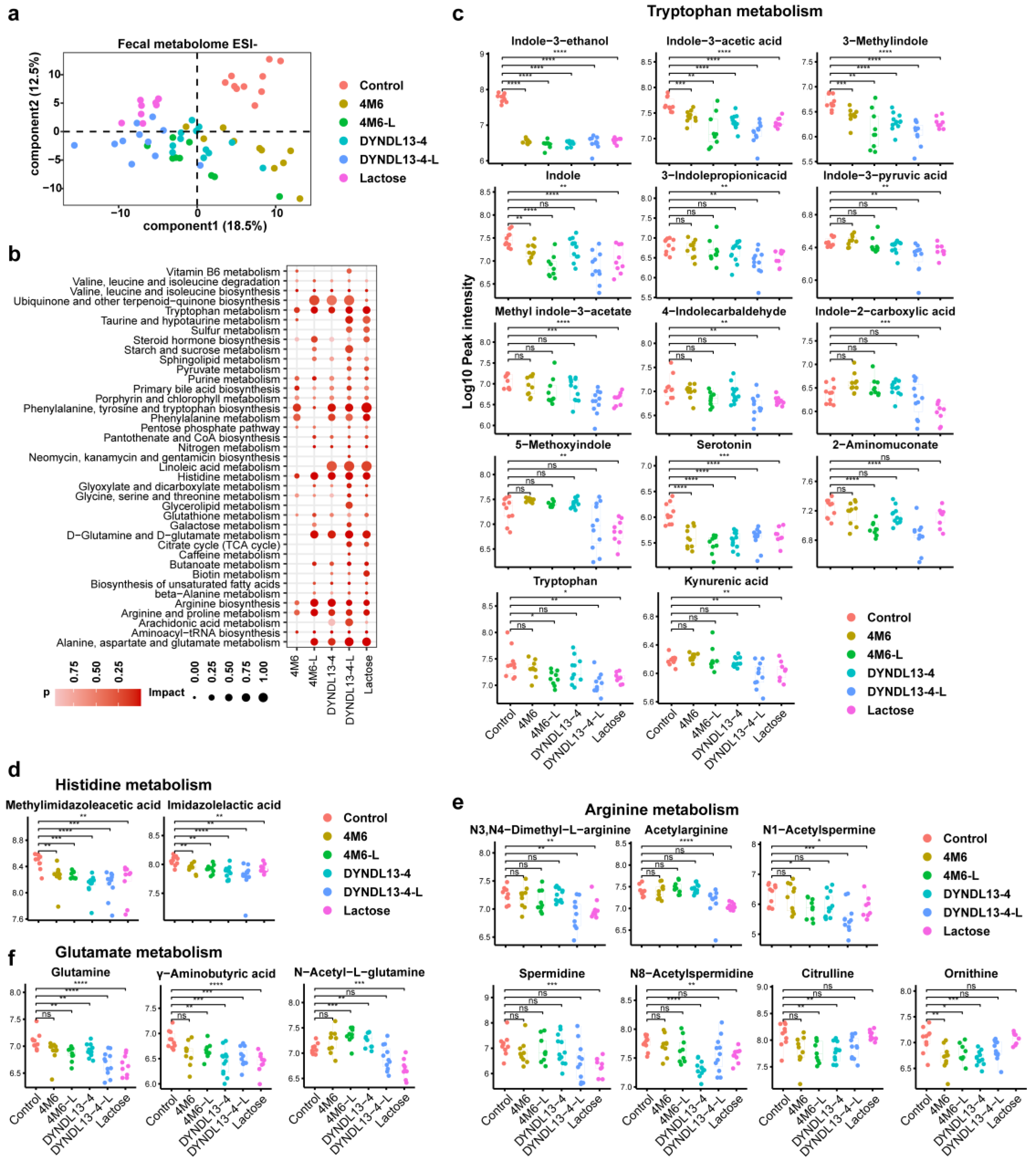
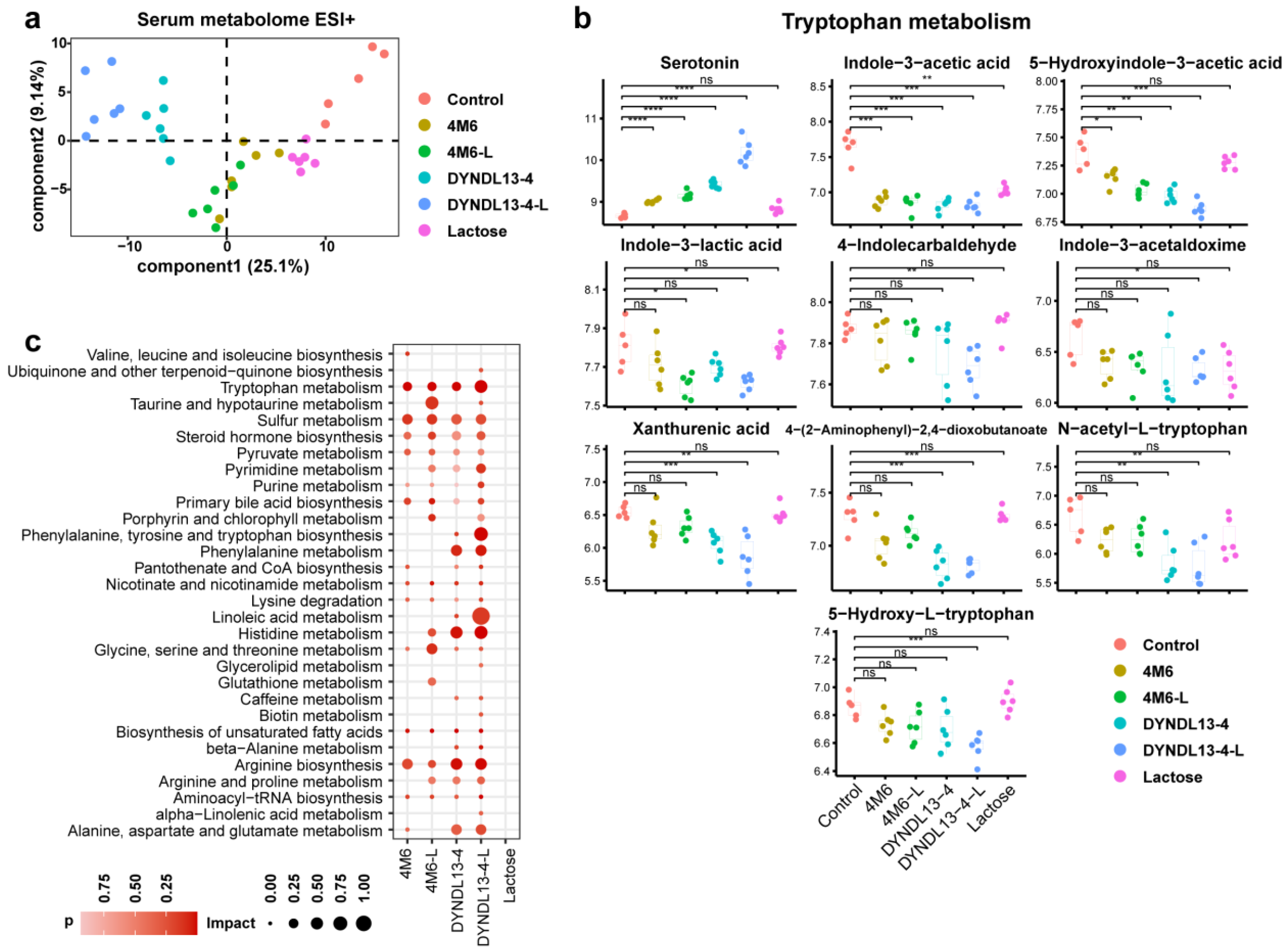
3.2. Effect of Lactose Supplementation on Fecal Metabolic Profile Induced by S. thermophilus Ingestion Is Similar to Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Q.; Hu, W.; Liu, W.X.; Zhao, L.Y.; Huang, D.; Liu, X.D.; Chan, H.; Zhang, Y.; Zeng, J.D.; Coker, O.O.; et al. Streptococcus thermophilus inhibits colorectal tumorigenesis through secreting beta-galactosidase. Gastroenterology 2021, 160, 1179–1193. [Google Scholar] [CrossRef] [PubMed]
- Tooley, K.L.; Howarth, G.S.; Lymn, K.A.; Lawrence, A.; Butler, R.N. Oral ingestion of Streptococcus thermophilus diminishes severity of small intestinal mucositis in methotrexate treated rats. Cancer Biol. Ther. 2006, 5, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, J.M.; Abi-Hanna, A.; Moore, N.; Yolken, R.H. Long-term consumption of infant formulas containing live probiotic bacteria: Tolerance and safety. Am. J. Clin. Nutr. 2004, 79, 261–267. [Google Scholar] [CrossRef]
- Canani, R.B.; Cirillo, P.; Terrin, G.; Cesarano, L.; Spagnuolo, M.I.; De Vincenzo, A.; Albano, F.; Passariello, A.; De Marco, G.; Manguso, F.; et al. Probiotics for treatment of acute diarrhoea in children: Randomised clinical trial of five different preparations. BMJ 2007, 335, 340. [Google Scholar] [CrossRef] [PubMed]
- Tillisch, K.; Labus, J.; Kilpatrick, L.; Jiang, Z.; Stains, J.; Ebrat, B.; Guyonnet, D.; Legrain-Raspaud, S.; Trotin, B.; Naliboff, B.; et al. Consumption of fermented milk product with probiotic modulates brain activity. Gastroenterology 2013, 144, 1394–1401. [Google Scholar] [CrossRef]
- Toca, M.D.; Fernandez, A.; Orsi, M.; Tabacco, O.; Vinderola, G. Lactose intolerance: Myths and facts. An update. Arch. Argent. Pediatr. 2022, 120, 59–66. [Google Scholar]
- JH, B.; MD, L. Quantitative measurement of lactose absorption. Gastroenterology 1976, 70, 1058–1062. [Google Scholar]
- Starz, E.; Wzorek, K.; Folwarski, M.; Kazmierczak-Siedlecka, K.; Stachowska, L.; Przewlocka, K.; Stachowska, E.; Skonieczna-Zydecka, K. The Modification of the Gut Microbiota via Selected Specific Diets in Patients with Crohn’s Disease. Nutrients 2021, 13, 2125. [Google Scholar] [CrossRef]
- Cederlund, A.; Kai-Larsen, Y.; Printz, G.; Yoshio, H.; Alvelius, G.; Lagercrantz, H.; Strömberg, R.; Jörnvall, H.; Gudmundsson, G.H.; Agerberth, B. Lactose in human breast milk an inducer of innate immunity with implications for a role in intestinal homeostasis. PLoS ONE 2013, 8, e53876. [Google Scholar] [CrossRef]
- Forsgard, R.A. Lactose digestion in humans: Intestinal lactase appears to be constitutive whereas the colonic microbiome is adaptable. Am. J. Clin. Nutr. 2019, 110, 273–279. [Google Scholar] [CrossRef]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Garay, J.A.R.; Finnicum, C.T.; Liu, X.R.; et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Sprenger, N.; Ghyselinck, J.; Marsaux, B.; Marzorati, M.; Rochat, F. A Comparison of the In vitro Effects of 2′Fucosyllactose and Lactose on the Composition and Activity of Gut Microbiota from Infants and Toddlers. Nutrients 2021, 13, 726. [Google Scholar] [CrossRef] [PubMed]
- Salli, K.; Anglenius, H.; Hiryonen, J.; Hibberd, A.A.; Ahonen, I.; Saarinen, M.T.; Tiihonen, K.; Maukonen, J.; Ouwehand, A.C. The effect of 2′-fucosyllactose on simulated infant gut microbiome and metabolites; a pilot study in comparison to GOS and lactose. Sci. Rep. 2019, 9, 13232. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Wrzosek, L.; Ben-Yahia, L.; Noordine, M.L.; Gitton, C.; Chevret, D.; Langella, P.; Mayeur, C.; Cherbuy, C.; Rul, F. Carbohydrate metabolism is essential for the colonization of Streptococcus thermophilus in the digestive tract of gnotobiotic rats. PLoS ONE 2011, 6, e28789. [Google Scholar] [CrossRef]
- Pan, Y.Q.; Yu, P.; Jiang, Y.; Liu, X.M.; Zhao, J.X.; Zhang, H.; Chen, W. The protective effect of lactose on the bile salt stress response of Streptococcus thermophilus is strain dependent. Food Biosci. 2023, 53, 102560. [Google Scholar] [CrossRef]
- Yang, B.; Chen, H.Q.; Gao, H.; Wang, J.T.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, W. Bifidobacterium breve CCFM683 could ameliorate DSS-induced colitis in mice primarily via conjugated linoleic acid production and gut microbiota modulation. J. Funct. Foods 2018, 49, 61–72. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J.G. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.Y.; Xia, J.G. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, C.; Yu, L.; Tian, F.; Wu, Y.; Zhao, J.; Zhang, H.; Yang, R.; Chen, W.; Hill, C.; et al. Human gut-derived B. longum subsp. longum strains protect against aging in a D-galactose-induced aging mouse model. Microbiome 2021, 9, 180. [Google Scholar] [CrossRef]
- Duque, A.L.R.F.; Demarqui, F.M.; Santoni, M.M.; Zanelli, C.F.; Adorno, M.A.T.; Milenkovic, D.; Mesa, V.; Sivieri, K. Effect of probiotic, prebiotic, and synbiotic on the gut microbiota of autistic children using an in vitro gut microbiome model. Food Res. Int. 2021, 149, 110657. [Google Scholar] [CrossRef]
- Shi, Y.; Zhai, Q.X.; Li, D.Y.; Mao, B.Y.; Liu, X.M.; Zhao, J.X.; Zhang, H.; Chen, W. Restoration of cefixime-induced gut microbiota changes by Lactobacillus cocktails and fructooligosaccharides in a mouse model. Microbiol. Res. 2017, 200, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, N.; Yang, J.J.; Zhao, D.M.; Chen, B.; Zhang, G.Q.; Chen, S.; Cao, R.F.; Yu, H.; Zhao, C.Y.; et al. Probiotics and fructo-oligosaccharide intervention modulate the microbiota-gut brain axis to improve autism spectrum reducing also the hyper-serotonergic state and the dopamine metabolism disorder. Pharmacol. Res. 2020, 157, 104784. [Google Scholar] [CrossRef] [PubMed]
- Succi, M.; Tremonte, P.; Pannella, G.; Tipaldi, L.; Cozzolino, A.; Romaniello, R.; Sorrentino, E.; Coppola, R. Pre-cultivation with Selected Prebiotics Enhances the Survival and the Stress Response of Lactobacillus rhamnosus Strains in Simulated Gastrointestinal Transit. Front. Microbiol. 2017, 8, 1067. [Google Scholar] [CrossRef] [PubMed]
- Kadlec, R.; Jakubec, M. The effect of prebiotics on adherence of probiotics. J. Dairy. Sci. 2014, 97, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Avila-Reyes, S.V.; Garcia-Suarez, F.J.; Jimenez, M.T.; San Martin-Gonzalez, M.F.; Bello-Perez, L.A. Protection of L. rhamnosus by spray-drying using two prebiotics colloids to enhance the viability. Carbohyd Polym. 2014, 102, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.Q.; Chen, Z.B.; Han, S.Y.; Zhang, S.B.; Zhang, T.F.; Lu, Y.M.; Berglund, B.; Xiao, H.; Li, L.J.; Yao, M.F. Role of prebiotics in enhancing the function of next-generation probiotics in gut microbiota. Crit. Rev. Food Sci. Nutr. 2023, 63, 1037–1054. [Google Scholar] [CrossRef]
- Firrman, J.; Liu, L.S.; Mahalak, K.; Hu, W.M.; Bittinger, K.; Moustafa, A.; Jones, S.M.; Narrowe, A.; Tomasula, P. An in vitro analysis of how lactose modifies the gut microbiota structure and function of adults in a donor-independent manner. Front. Nutr. 2023, 9, 1040744. [Google Scholar] [CrossRef]
- Sharon, G.; Garg, N.; Debelius, J.; Knight, R.; Dorrestein, P.C.; Mazmanian, S.K. Specialized Metabolites from the Microbiome in Health and Disease. Cell Metab. 2014, 20, 719–730. [Google Scholar] [CrossRef]
- Laurans, L.; Venteclef, N.; Haddad, Y.; Chajadine, M.; Alzaid, F.; Metghalchi, S.; Sovran, B.; Denis, R.G.P.; Dairou, J.; Cardellini, M.; et al. Genetic deficiency of indoleamine 2,3-dioxygenase promotes gut microbiota-mediated metabolic health. Nat. Med. 2018, 24, 1113–1120. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef]
- Lamas, B.; Richard, M.L.; Leducq, V.; Pham, H.P.; Michel, M.L.; Da Costa, G.; Bridonneau, C.; Jegou, S.; Hoffmann, T.W.; Natividad, J.M.; et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nat. Med. 2016, 22, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Alexeev, E.E.; Lanis, J.M.; Kao, D.J.; Campbell, E.L.; Kelly, C.J.; Battista, K.D.; Gerich, M.E.; Jenkins, B.R.; Walk, S.T.; Kominsky, D.J.; et al. Microbiota-Derived Indole Metabolites Promote Human and Murine Intestinal Homeostasis through Regulation of Interleukin-10 Receptor. Am. J. Pathol. 2018, 188, 1183–1194. [Google Scholar] [CrossRef]
- Whitfield-Cargile, C.M.; Cohen, N.D.; Chapkin, R.S.; Weeks, B.R.; Davidson, L.A.; Goldsby, J.S.; Hunt, C.L.; Steinmeyer, S.H.; Menon, R.; Suchodolski, J.S.; et al. The microbiota-derived metabolite indole decreases mucosal inflammation and injury in a murine model of NSAID enteropathy. Gut Microbes 2016, 7, 246–261. [Google Scholar] [CrossRef] [PubMed]
- Rosas, H.D.; Doros, G.; Bhasin, S.; Thomas, B.; Gevorkian, S.; Malarick, K.; Matson, W.; Hersch, S.M. A systems-level “misunderstanding”: The plasma metabolome in Huntington’s disease. Ann. Clin. Transl. Neur 2015, 2, 756–768. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.H.; Huang, J.X.; Xiong, X.; Guo, Y.M.; Zhang, B.K. Role of Nutrient-sensing Receptor GPRC6A in Regulating Colonic Group 3 Innate Lymphoid Cells and Inflamed Mucosal Healing. J. Crohns Colitis 2022, 16, 1293–1305. [Google Scholar] [CrossRef] [PubMed]
- Yeung, S.L.A.; Lin, S.L.; Lam, H.S.H.S.; Schooling, C.M. Effect of L-arginine, asymmetric dimethylarginine, and symmetric dimethylarginine on ischemic heart disease risk: A Mendelian randomization study. Am. Heart J. 2016, 182, 54–61. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Besevic, J.; Dimou, N.; Breeur, M.; Murphy, N.; Jenab, M.; Wedekind, R.; Viallon, V.; Ferrari, P.; Achaintre, D.; et al. Circulating amino acid levels and colorectal cancer risk in the European Prospective Investigation into Cancer and Nutrition and UK Biobank. BMC Med. 2023, 21, 80. [Google Scholar] [CrossRef]
- Molinaro, A.; Bel Lassen, P.; Henricsson, M.; Wu, H.; Adriouch, S.; Belda, E.; Chakaroun, R.; Nielsen, T.; Bergh, P.O.; Rouault, C.; et al. Imidazole propionate is increased in diabetes and associated with dietary patterns and altered microbial ecology. Nat. Commun. 2020, 11, 5881. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef]
- Trimigno, A.; Münger, L.; Picone, G.; Freiburghaus, C.; Pimentel, G.; Vionnet, N.; Pralong, F.; Capozzi, F.; Badertscher, R.; Vergères, G. GC-MS Based Metabolomics and NMR Spectroscopy Investigation of Food Intake Biomarkers for Milk and Cheese in Serum of Healthy Humans. Metabolites 2018, 8, 26. [Google Scholar] [CrossRef]
- Kim, J.; Blaser, C.; Portmann, R.; Badertscher, R.; Marmonier, C.; Blot, A.; David, J.; Stoffers, H.; von Ah, U.; Butikofer, U.; et al. Postprandial Responses on Serum Metabolome to Milk and Yogurt Intake in Young and Older Men. Front. Nutr. 2022, 9, 851931. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, P.; Pan, Y.; Pei, Z.; Guo, M.; Yang, B.; Lee, Y.-K.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Influence of Lactose Supplementation on Regulation of Streptococcus thermophilus on Gut Microbiota. Nutrients 2023, 15, 4767. https://doi.org/10.3390/nu15224767
Yu P, Pan Y, Pei Z, Guo M, Yang B, Lee Y-K, Liu X, Zhao J, Zhang H, Chen W. Influence of Lactose Supplementation on Regulation of Streptococcus thermophilus on Gut Microbiota. Nutrients. 2023; 15(22):4767. https://doi.org/10.3390/nu15224767
Chicago/Turabian StyleYu, Peng, Yuqi Pan, Zhiwen Pei, Min Guo, Bo Yang, Yuan-Kun Lee, Xiaoming Liu, Jianxin Zhao, Hao Zhang, and Wei Chen. 2023. "Influence of Lactose Supplementation on Regulation of Streptococcus thermophilus on Gut Microbiota" Nutrients 15, no. 22: 4767. https://doi.org/10.3390/nu15224767
APA StyleYu, P., Pan, Y., Pei, Z., Guo, M., Yang, B., Lee, Y.-K., Liu, X., Zhao, J., Zhang, H., & Chen, W. (2023). Influence of Lactose Supplementation on Regulation of Streptococcus thermophilus on Gut Microbiota. Nutrients, 15(22), 4767. https://doi.org/10.3390/nu15224767