

1,25-Dihydroxyvitamin D3 Provides Benefits in Vitiligo Based on Modulation of CD8+ T Cell Glycolysis and Function
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Procedure
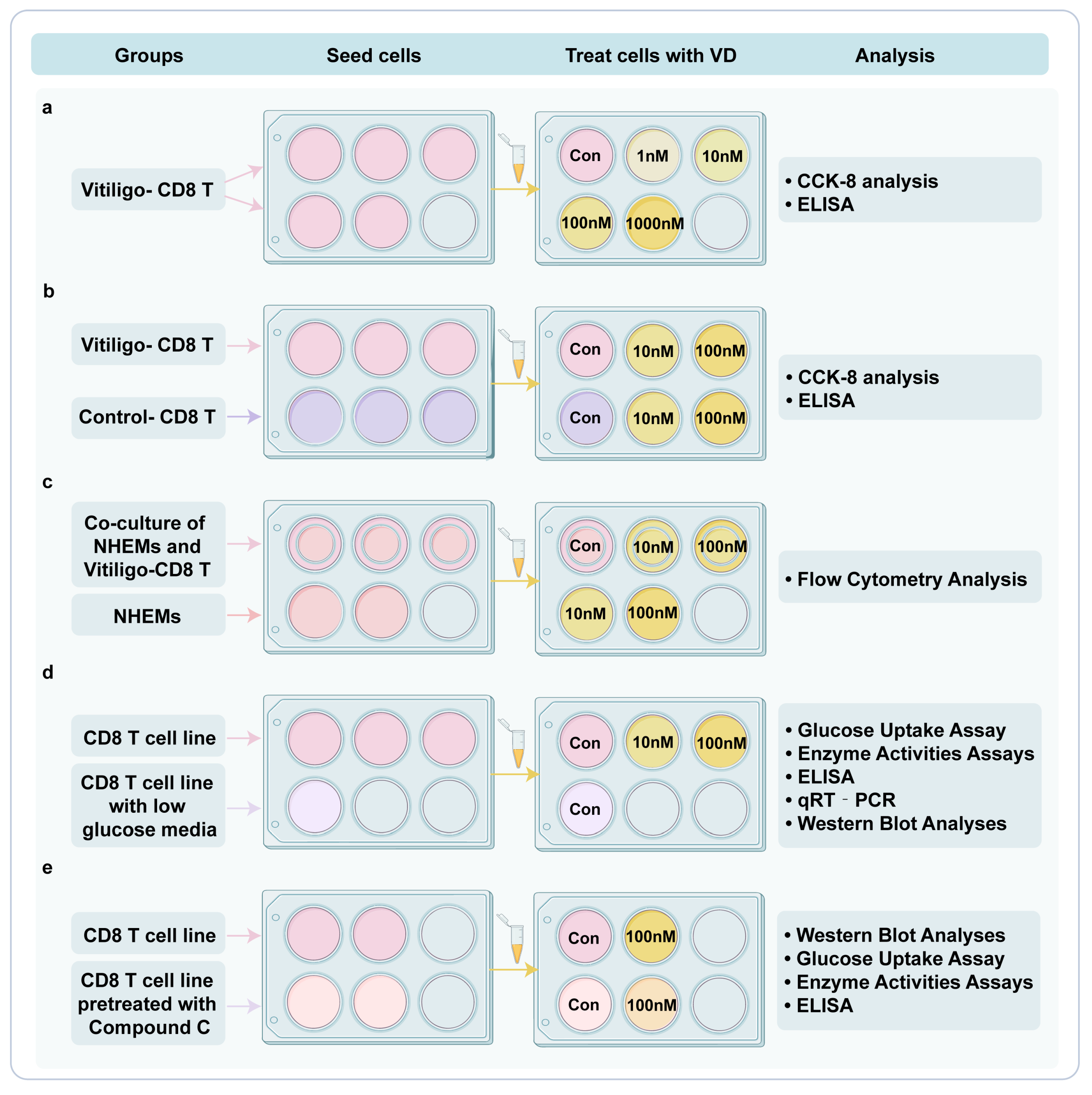
2.2.1. Cell Culture and Treatment
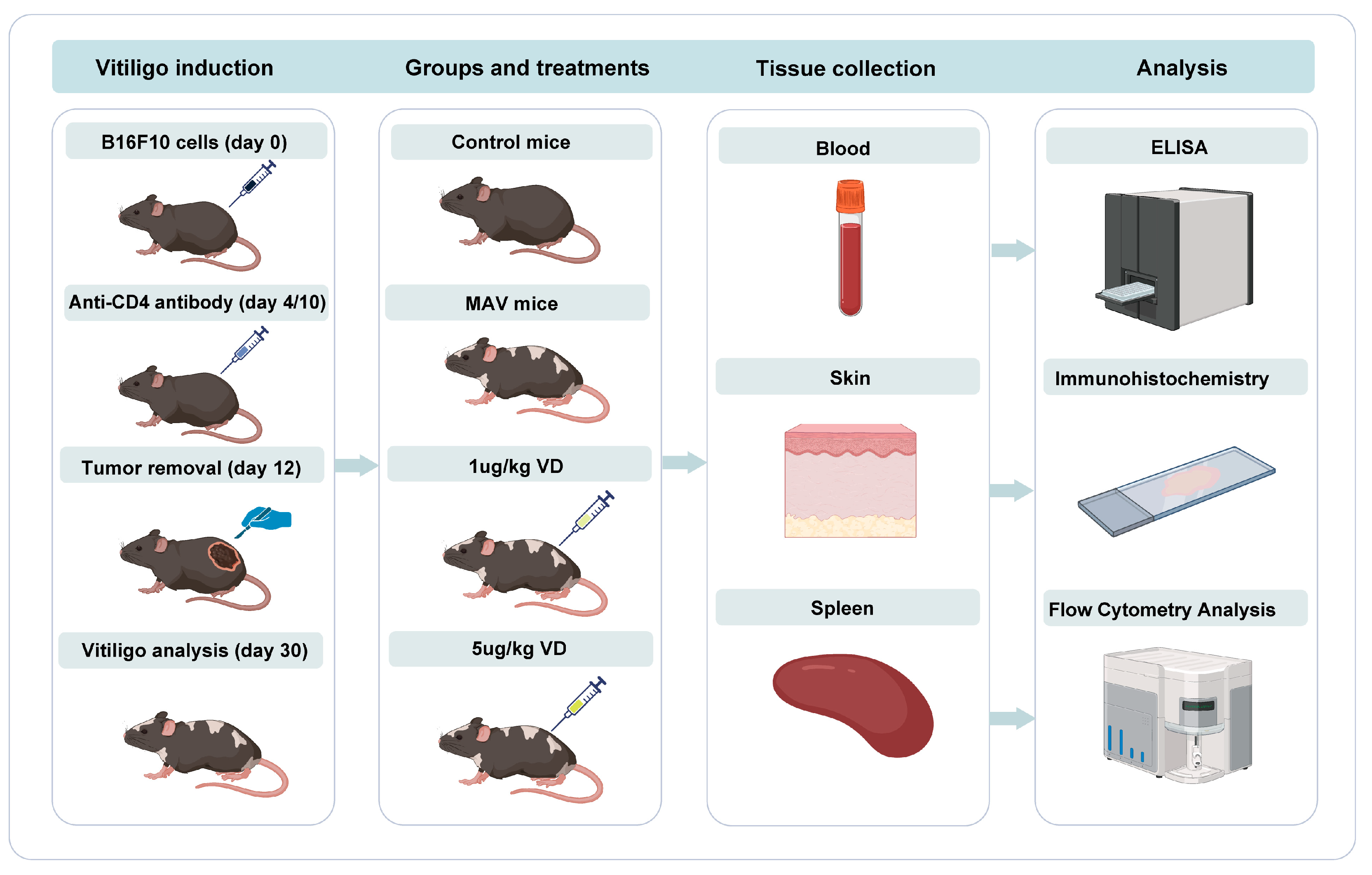
2.2.2. Animals and Drug Treatment Protocols
2.2.3. Cell Viability
2.2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.2.5. Flow Cytometry Analysis
2.2.6. Glucose Uptake Assay
2.2.7. Enzyme Activity Assays
2.2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.2.9. Western Blot Analysis
2.2.10. Immunohistochemistry
2.3. Data Sources and Analysis
2.3.1. Data Source and Bioinformatic Analysis
2.3.2. Data Presentation and Statistical Analysis
3. Results
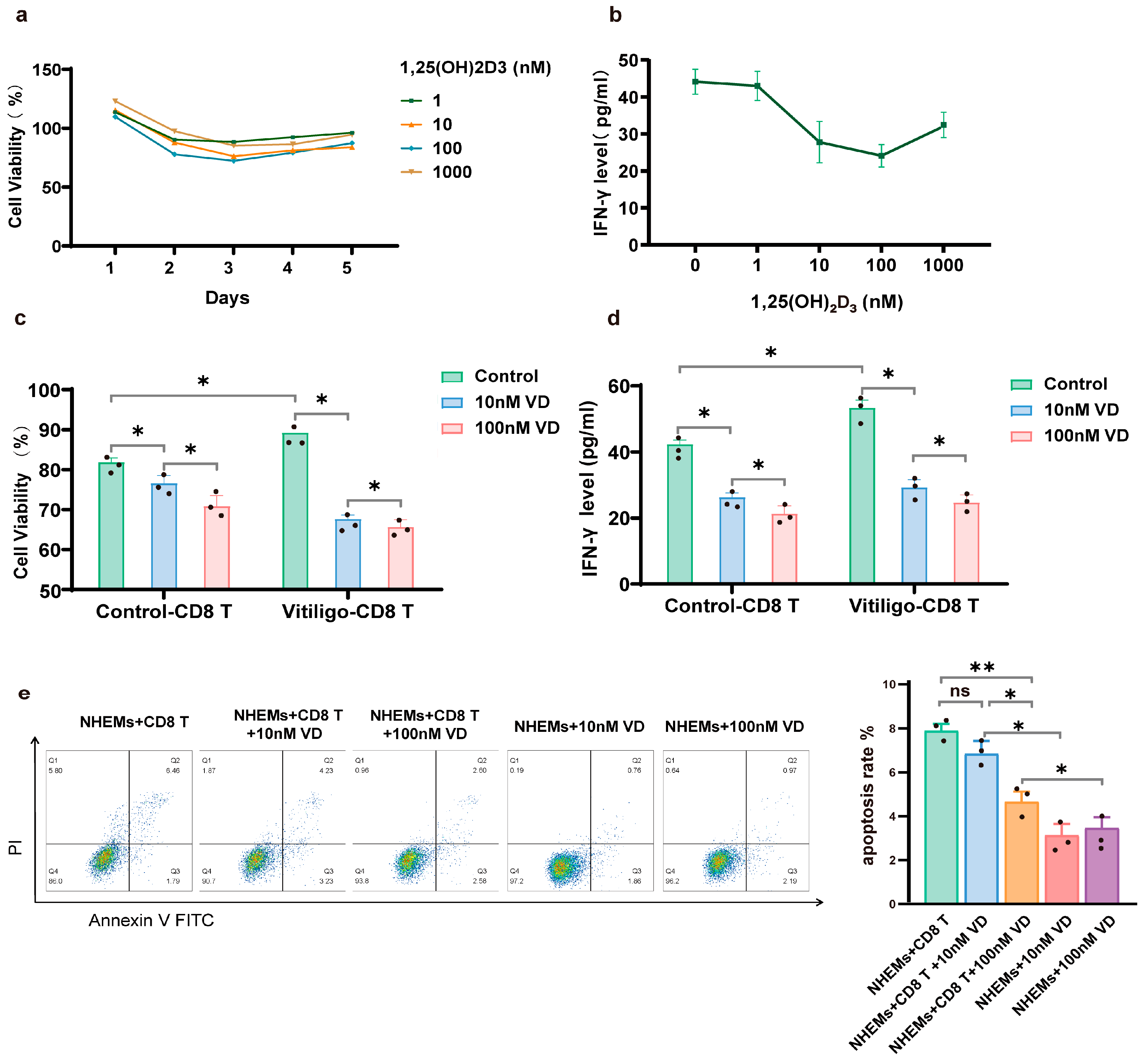
3.1. Deficiency of 1,25(OH)₂D₃ Is Associated with Disease Activity and Dysfunction of CD8+ T Cells in the Vitiligo Cohort
3.2. 1,25(OH)₂D₃ Influences Glucose Metabolism and Signaling Pathways of Circulating Immune Cells
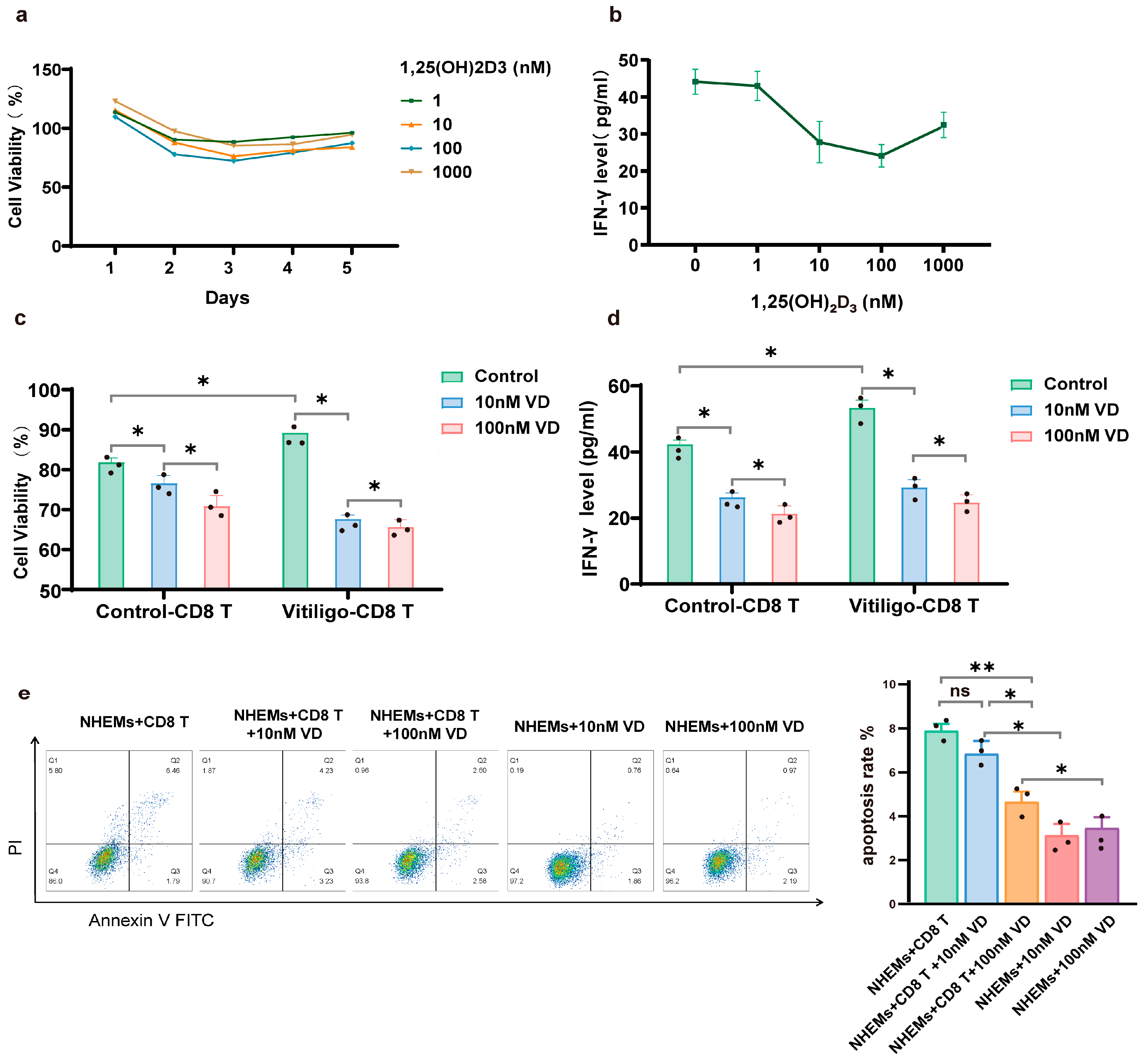
3.3. 1,25(OH)₂D₃ Inhibits Proliferation and Cytotoxicity of CD8+ T Cells
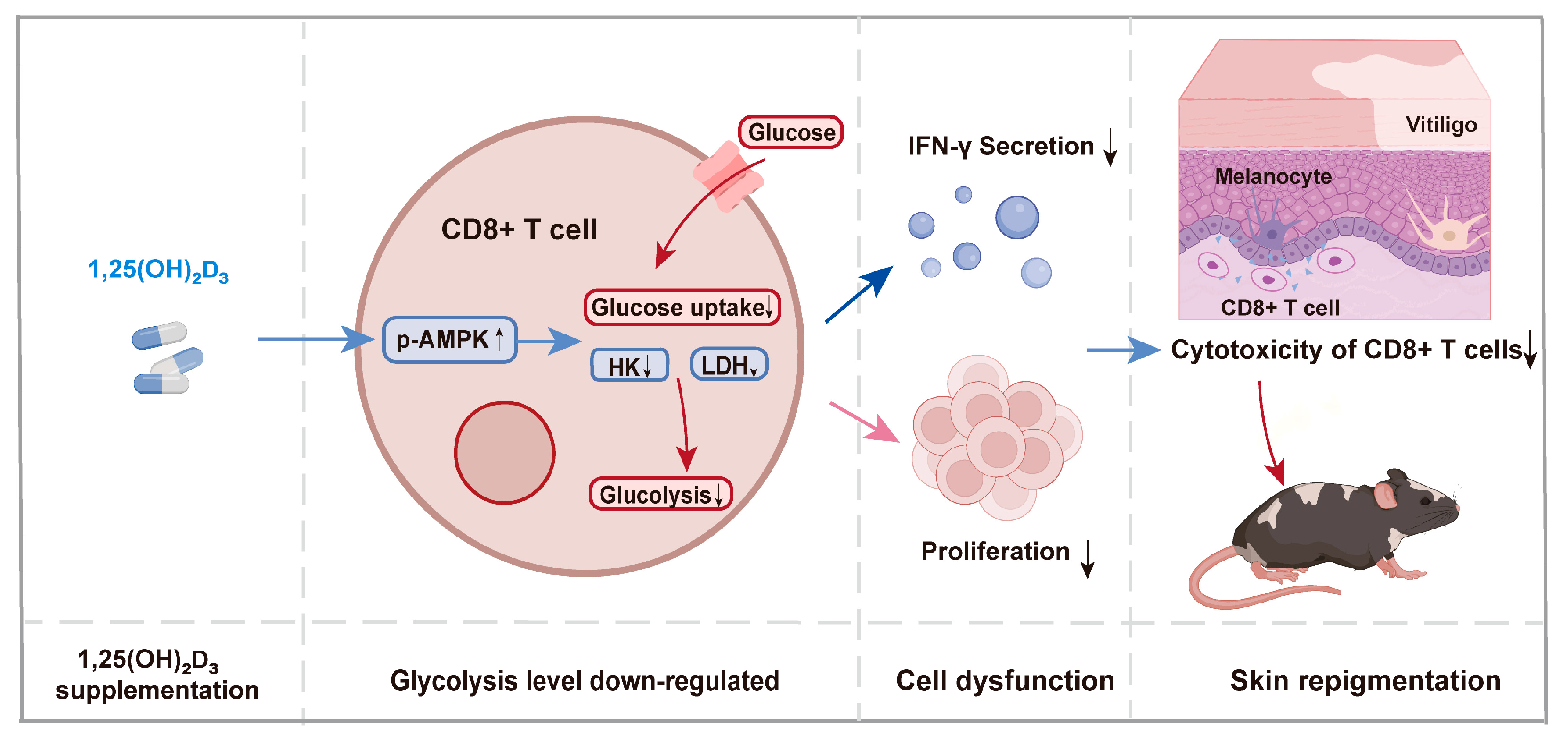
3.4. 1,25(OH)₂D₃ Inhibits Glycolysis of CD8+ T Cells by Regulating the AMPK Pathway
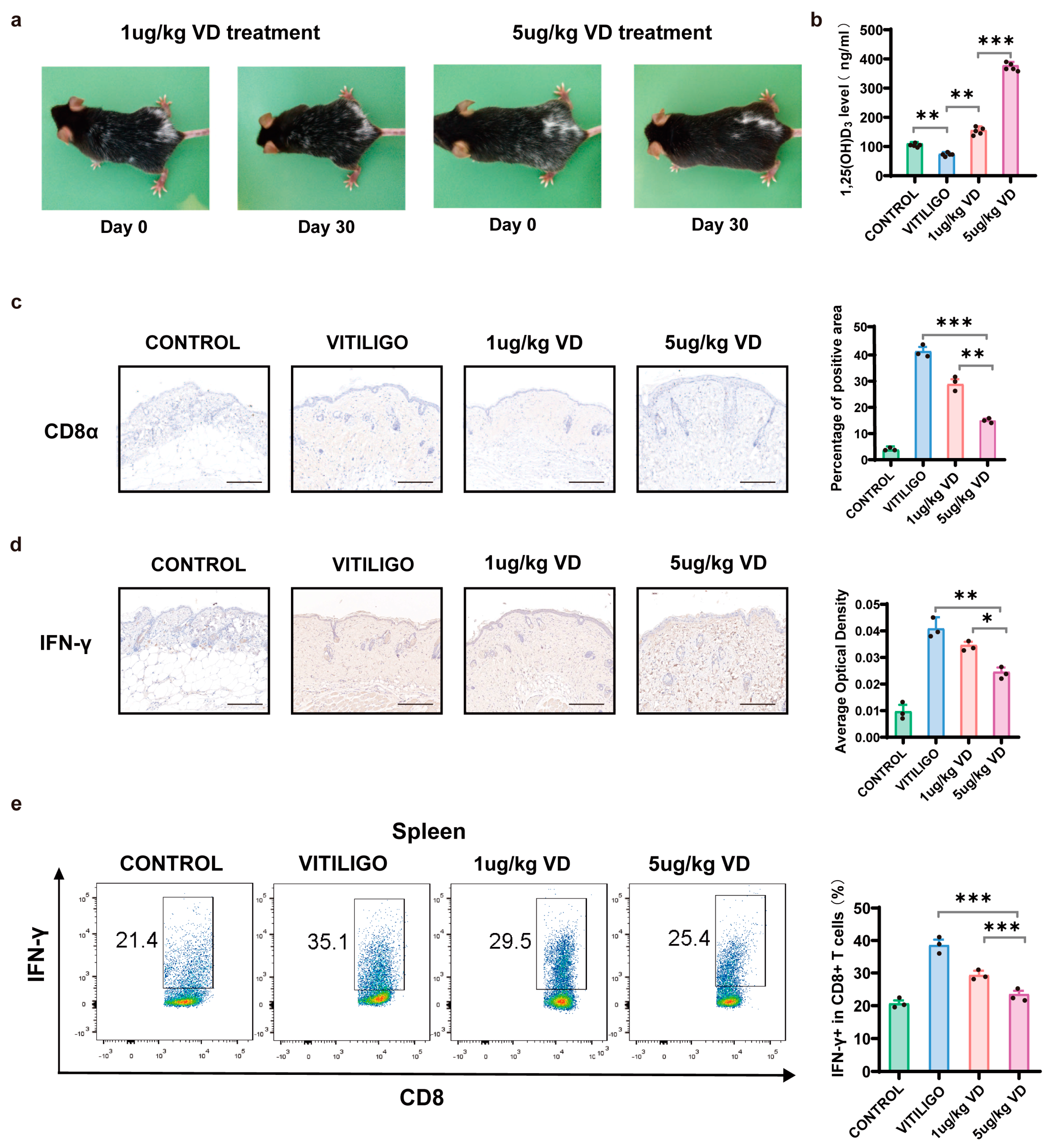
3.5. Therapeutic Effect of 1,25(OH)₂D₃ on Melanocyte-Associated Vitiligo Mice
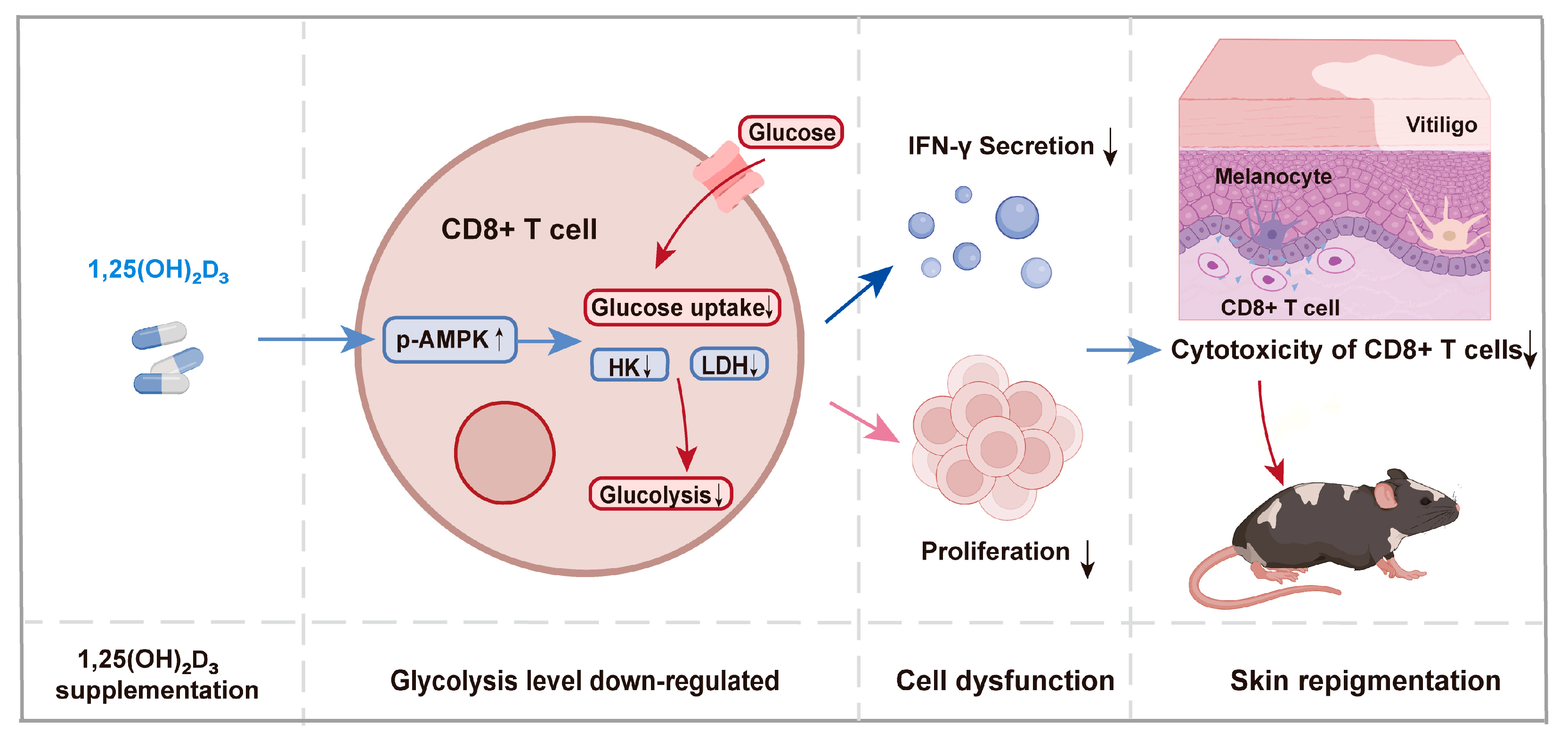
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bergqvist, C.; Ezzedine, K. Vitiligo: A Review. Dermatology 2020, 236, 571–592. [Google Scholar] [CrossRef] [PubMed]
- Frisoli, M.L.; Essien, K.; Harris, J.E. Vitiligo: Mechanisms of Pathogenesis and Treatment. Annu. Rev. Immunol. 2020, 38, 621–648. [Google Scholar] [CrossRef] [PubMed]
- Speeckaert, R.; van Geel, N. Vitiligo: An Update on Pathophysiology and Treatment Options. Am. J. Clin. Dermatol. 2017, 18, 733–744. [Google Scholar] [CrossRef]
- Wilson, L.R.; Tripkovic, L.; Hart, K.H.; Lanham-New, S.A. Vitamin D deficiency as a public health issue: Using vitamin D2 or vitamin D3 in future fortification strategies. Proc. Nutr. Soc. 2017, 76, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Penna, G.; Adorini, L. 1α,25-Dihydroxyvitamin D3 Inhibits Differentiation, Maturation, Activation, and Survival of Dendritic Cells Leading to Impaired Alloreactive T Cell Activation. J. Immunol. 2000, 164, 2405–2411. [Google Scholar] [CrossRef] [PubMed]
- Pludowski, P.; Holick, M.F.; Pilz, S.; Wagner, C.L.; Hollis, B.W.; Grant, W.B.; Shoenfeld, Y.; Lerchbaum, E.; Llewellyn, D.J.; Kienreich, K.; et al. Vitamin D effects on musculoskeletal health, immunity, autoimmunity, cardiovascular disease, cancer, fertility, pregnancy, dementia and mortality-a review of recent evidence. Autoimmun. Rev. 2013, 12, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Atazadeh, F.; Fazeli, Z.; Vahidnezhad, H.; Namazi, N.; Younespour, S.; Youssefian, L.; Abdollahimajd, F.; Uitto, J. Increased level of cathelicidin (LL-37) in vitiligo: Possible pathway independent from vitamin D receptor gene polymorphism. Exp. Dermatol. 2020, 29, 1176–1185. [Google Scholar] [CrossRef]
- Silverberg, J.I.; Silverberg, A.I.; Malka, E.; Silverberg, N.B. A pilot study assessing the role of 25 hydroxy vitamin D levels in patients with vitiligo vulgaris. J. Am. Acad. Dermatol. 2010, 62, 937–941. [Google Scholar] [CrossRef]
- Khurrum, H.; AlGhamdi, K.M. The Relationship Between the Serum Level of Vitamin D and Vitiligo: A Controlled Study on 300 Subjects. J. Cutan. Med. Surg. 2016, 20, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liu, J.; Li, W.; Zheng, H.; Dai, H.; Qiu, G.; Yu, D.; Yao, D.; Yin, X. Causal Associations between Vitamin D Levels and Psoriasis, Atopic Dermatitis, and Vitiligo: A Bidirectional Two-Sample Mendelian Randomization Analysis. Nutrients 2022, 14, 5284. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, D.; Hu, Y.; Jiang, K.; Huang, H.; Du, Y.; Wu, W.; Wang, J.; Sui, J.; Wang, W.; et al. Anatomically distinct fibroblast subsets determine skin autoimmune patterns. Nature 2022, 601, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Sukumar, M.; Liu, J.; Ji, Y.; Subramanian, M.; Crompton, J.G.; Yu, Z.; Roychoudhuri, R.; Palmer, D.C.; Muranski, P.; Karoly, E.D.; et al. Inhibiting glycolytic metabolism enhances CD8+ T cell memory and antitumor function. J. Clin. Investig. 2013, 123, 4479–4488. [Google Scholar] [CrossRef]
- Thien, R.; Baier, K.; Pietschmann, P.; Peterlik, M.; Willheim, M. Interactions of 1α,25-dihydroxyvitamin D3 with IL-12 and IL-4 on cytokine expression of human T lymphocytes. J. Allergy Clin. Immunol. 2005, 116, 683–689. [Google Scholar] [CrossRef]
- Bishop, E.L.; Gudgeon, N.H.; Mackie, G.M.; Chauss, D.; Roberts, J.; Tennant, D.A.; Maslowski, K.M.; Afzali, B.; Hewison, M.; Dimeloe, S. 1,25-Dihydroxyvitamin D3 suppresses CD4(+) T-cell effector functionality by inhibition of glycolysis. Immunology 2022, 166, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Xu, Z.; Cui, J.; Chen, T. A mouse model of vitiligo based on endogenous auto-reactive CD8 + T cell targeting skin melanocyte. Cell Regen. 2022, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Boniface, K.; Seneschal, J.; Picardo, M.; Taïeb, A. Vitiligo: Focus on Clinical Aspects, Immunopathogenesis, and Therapy. Clin. Rev. Allergy Immunol. 2018, 54, 52–67. [Google Scholar] [CrossRef]
- Youssef, R.; Emad, N.; Mogawer, R.M. Comparative Analysis of the Most Used Versus the Recently Developed Vitiligo Activity and Extent Scores and Their Change with Treatment. Dermatol. Pr. Concept. 2023, 13, e2023186. [Google Scholar] [CrossRef]
- Li, D.; Zhou, T.; She, Q.; Nie, X.; Liu, Z.; Pan, R.; Wei, Y.; Deng, Y. Circulating Exosomal miR-493-3p Affects Melanocyte Survival and Function by Regulating Epidermal Dopamine Concentration in Segmental Vitiligo. J. Investig. Dermatol. 2022, 142, 3262–3273. [Google Scholar] [CrossRef]
- Lo, J.A.; Kawakubo, M.; Juneja, V.R.; Su, M.Y.; Erlich, T.H.; LaFleur, M.W.; Kemeny, L.V.; Rashid, M.; Malehmir, M.; Rabi, S.A.; et al. Epitope spreading toward wild-type melanocyte-lineage antigens rescues suboptimal immune checkpoint blockade responses. Sci. Transl. Med. 2021, 13, eabd8636. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Nishikawa, H.; Sugiyama, D.; Ha, D.; Hamaguchi, M.; Saito, T.; Nishioka, M.; Wing, J.B.; Adeegbe, D.; Katayama, I.; et al. Detection of self-reactive CD8+ T cells with an anergic phenotype in healthy individuals. Science 2014, 346, 1536–1540. [Google Scholar] [CrossRef]
- Jin, R.; Zhou, M.; Lin, F.; Xu, W.; Xu, A. Pathogenic Th2 Cytokine Profile Skewing by IFN-γ-Responding Vitiligo Fibroblasts via CCL2/CCL8. Cells 2023, 12, 217. [Google Scholar]
- Halprin, K.M. Epidermal “turnover time”–A re-examination. Br. J. Dermatol. 1972, 86, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Ao, T.; Kikuta, J.; Ishii, M. The Effects of Vitamin D on Immune System and Inflammatory Diseases. Biomolecules 2021, 11, 1624. [Google Scholar] [CrossRef] [PubMed]
- Baeke, F.; Takiishi, T.; Korf, H.; Gysemans, C.; Mathieu, C. Vitamin D: Modulator of the immune system. Curr. Opin. Pharmacol. 2010, 10, 482–496. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S.; Li, C. Clinical Features, Immunopathogenesis, and Therapeutic Strategies in Vitiligo. Clin. Rev. Allergy Immunol. 2021, 61, 299–323. [Google Scholar] [CrossRef] [PubMed]
- Cantorna, M.T.; Waddell, A. The vitamin D receptor turns off chronically activated T cells. Ann. N. Y Acad. Sci. 2014, 1317, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Liao, S.; Zeng, F.; Liao, Q.; Luo, G.; Zhou, Y. Effects of altered glycolysis levels on CD8+ T cell activation and function. Cell Death Dis. 2023, 14, 407. [Google Scholar] [CrossRef]
- Souto-Carneiro, M.M.; Klika, K.D.; Abreu, M.T.; Meyer, A.P.; Saffrich, R.; Sandhoff, R.; Jennemann, R.; Kraus, F.V.; Tykocinski, L.; Eckstein, V.; et al. Effect of Increased Lactate Dehydrogenase A Activity and Aerobic Glycolysis on the Proinflammatory Profile of Autoimmune CD8+ T Cells in Rheumatoid Arthritis. Arthritis Rheumatol. 2020, 72, 2050–2064. [Google Scholar] [CrossRef] [PubMed]
- Prunk, M.; Perišić Nanut, M.; Jakoš, T.; Sabotič, J.; Švajger, U.; Kos, J. Extracellular Cystatin F Is Internalised by Cytotoxic T Lymphocytes and Decreases Their Cytotoxicity. Cancers 2020, 12, 3660. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.; Cesano, A.; Lange, B.; Finan, J.; Nowell, P.C.; Clark, S.C.; Raimondi, S.C.; Rovera, G.; Santoli, D. Growth Factor Requirements of Childhood Acute T-Lymphoblastic Leukemia: Correlation Between Presence of Chromosomal Abnormalities and Ability to Grow Permanently In Vitro. Blood 1991, 77, 1534–1545. [Google Scholar] [CrossRef]
- Ma, E.H.; Poffenberger, M.C.; Wong, A.H.T.; Jones, R.G. The role of AMPK in T cell metabolism and function. Curr. Opin. Immunol. 2017, 46, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Cool, B.; Zinker, B.; Chiou, W.; Kifle, L.; Cao, N.; Perham, M.; Dickinson, R.; Adler, A.; Gagne, G.; Iyengar, R.; et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome. Cell Metab. 2006, 3, 403–416. [Google Scholar] [CrossRef]
- Blagih, J.; Coulombe, F.; Vincent, E.E.; Dupuy, F.; Galicia-Vázquez, G.; Yurchenko, E.; Raissi, T.C.; van der Windt, G.J.W.; Viollet, B.; Pearce, E.L.; et al. The Energy Sensor AMPK Regulates T Cell Metabolic Adaptation and Effector Responses In Vivo. Immunity 2015, 42, 41–54. [Google Scholar] [CrossRef]
- Liu, X.; Chhipa, R.R.; Nakano, I.; Dasgupta, B. The AMPK inhibitor compound C is a potent AMPK-independent antiglioma agent. Mol. Cancer Ther. 2014, 13, 596–605. [Google Scholar] [CrossRef] [PubMed]
- AlGhamdi, K.; Kumar, A.; Moussa, N. The role of vitamin D in melanogenesis with an emphasis on vitiligo. Indian. J. Dermatol. Venereol. Leprol. 2013, 79, 750–758. [Google Scholar] [CrossRef]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef]
- Karagüzel, G.; Sakarya, N.P.; Bahadır, S.; Yaman, S.; Ökten, A. Vitamin D status and the effects of oral vitamin D treatment in children with vitiligo: A prospective study. Clin. Nutr. ESPEN 2016, 15, 28–31. [Google Scholar] [CrossRef]
- Finamor, D.C.; Sinigaglia-Coimbra, R.; Neves, L.C.; Gutierrez, M.; Silva, J.J.; Torres, L.D.; Surano, F.; Neto, D.J.; Novo, N.F.; Juliano, Y.; et al. A pilot study assessing the effect of prolonged administration of high daily doses of vitamin D on the clinical course of vitiligo and psoriasis. Dermatoendocrinol 2013, 5, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Glebocka, A.; Chiellini, G. A-ring analogs of 1,25-dihydroxyvitamin D3. Arch. Biochem. Biophys. 2012, 523, 48–57. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| GLUT1 | GGCCAAGAGTGTGCTAAAGAA | ACAGCGTTGATGCCAGACAG |
| LDHA | ATGGCAACTCTAAAGGATCAGC | CCAACCCCAACAACTGTAATCT |
| HKII | GAGCCACCACTCACCCTACT | CCAGGCATTCGGCAATGTG |
| IFNG | TCGGTAACTGACTTGAATGTCCA | TCGCTTCCCTGTTTTAGCTGC |
| β-actin | CATGTACGTTGCTATCCAGGC | CTCCTTAATGTCACGCACGAT |
| All (N = 327) | 1,25(OH)₂D₃ Sufficient (≥20 ng/mL, N = 94, 28.74%) | 1,25(OH)₂D₃ Deficient (<20 ng/mL, N = 233, 71.62%) | p | |
|---|---|---|---|---|
| Serum 1,25(OH)₂D₃ concentration, mean ± SD, μg/mL | 16.73 ± 6.353 | 24.82 ± 3.745 | 13.47 ± 3.730 | <0.0001 |
| Disease duration, mean ± SD, month | 46.2 6 ± 82.62 | 50.36 ± 97.79 | 44.61 ± 75.82 | 0.1826 |
| total counts of T cells, mean ± SD | 1442 ± 511.8 | 1372 ± 527.5 | 1470 ± 503.7 | 0.0830 |
| CD8+ T cells/T cells, mean ± SD, % | 24.36 ± 5.793 | 22.88 ± 5.444 | 24.96 ± 5.834 | 0.0019 |
| IFN−γ+ CD8+ T cells/CD8+ T cells, mean ± SD, % | 38.69 ± 12.90 | 36.26 ± 15.466 | 39.67 ± 11.61 | 0.0011 |
| VASI, mean ± SD | 1.904 ± 5.437 | 1.422 ± 2.238 | 2.099 ± 6.277 | 0.2683 |
| VIDA, No (%) | 0.0024 | |||
| −1 | 7 (2.14%) | 3 (3.19%) | 4 (1.72%) | |
| 0 | 13 (3.98%) | 7 (7.45%) | 6 (2.58%) | |
| 1 | 21 (6.42%) | 4 (4.26%) | 17 (7.3%) | |
| 2 | 70 (21.41%) | 30 (31.91%) | 40 (17.17%) | |
| 3 | 85 (25.99%) | 25 (26.6%) | 60 (25.75%) | |
| 4 | 131 (40.06%) | 25 (26.6%) | 106 (45.49%) | |
| Types of vitiligo, No (%) | 0.7556 | |||
| Nonsegmental | 194 (59.33%) | 54 (57.45%) | 140 (60.09%) | |
| Segmental | 95 (29.05%) | 30 (31.91%) | 65 (27.9%) | |
| Unclassified | 38 (11.62%) | 10 (10.64%) | 28 (12.02%) | |
| Sex, No (%) | 0.0930 | |||
| Male | 157 (48.01%) | 52 (55.32%) | 105 (45.06%) | |
| Female | 170 (51.99%) | 42 (44.68%) | 128 (54.94%) | |
| Smoking, No. (%) | 0.2121 | |||
| No | 304 (92.97%) | 90 (95.74%) | 214 (91.85%) | |
| Yes | 23 (7.03%) | 4 (4.26%) | 19 (8.15%) | |
| Drinking, No (%) | 0.3650 | |||
| No | 311 (95.11%) | 91 (96.81%) | 220 (94.42%) | |
| Yes | 16 (4.89%) | 3 (3.19%) | 13 (5.58%) | |
| Insomnia, No (%) | 0.1619 | |||
| No | 278 (85.02%) | 84 (89.36%) | 194 (83.26%) | |
| Yes | 49 (14.98%) | 10 (10.64%) | 39 (16.74%) | |
| Hereditary, No (%) | 0.8362 | |||
| No | 263 (80.43%) | 78 (82.98%) | 187 (80.26%) | |
| Yes | 64 (19.57%) | 18 (19.15%) | 46 (19.74%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Wang, T.; Nie, X.; Shi, Z.; Liu, Z.; Zeng, Y.; Pan, R.; Zhang, R.; Deng, Y.; Li, D. 1,25-Dihydroxyvitamin D3 Provides Benefits in Vitiligo Based on Modulation of CD8+ T Cell Glycolysis and Function. Nutrients 2023, 15, 4697. https://doi.org/10.3390/nu15214697
Wei Y, Wang T, Nie X, Shi Z, Liu Z, Zeng Y, Pan R, Zhang R, Deng Y, Li D. 1,25-Dihydroxyvitamin D3 Provides Benefits in Vitiligo Based on Modulation of CD8+ T Cell Glycolysis and Function. Nutrients. 2023; 15(21):4697. https://doi.org/10.3390/nu15214697
Chicago/Turabian StyleWei, Yujia, Tingmei Wang, Xiaoqi Nie, Zeqi Shi, Zhong Liu, Ying Zeng, Ronghua Pan, Ri Zhang, Yunhua Deng, and Dong Li. 2023. "1,25-Dihydroxyvitamin D3 Provides Benefits in Vitiligo Based on Modulation of CD8+ T Cell Glycolysis and Function" Nutrients 15, no. 21: 4697. https://doi.org/10.3390/nu15214697
APA StyleWei, Y., Wang, T., Nie, X., Shi, Z., Liu, Z., Zeng, Y., Pan, R., Zhang, R., Deng, Y., & Li, D. (2023). 1,25-Dihydroxyvitamin D3 Provides Benefits in Vitiligo Based on Modulation of CD8+ T Cell Glycolysis and Function. Nutrients, 15(21), 4697. https://doi.org/10.3390/nu15214697