

Effect of Cordyceps militaris Powder Prophylactic Supplementation on Intestinal Mucosal Barrier Impairment and Microbiota-Metabolites Axis in DSS-Injured Mice

Abstract
:
1. Introduction
2. Materials and Methods
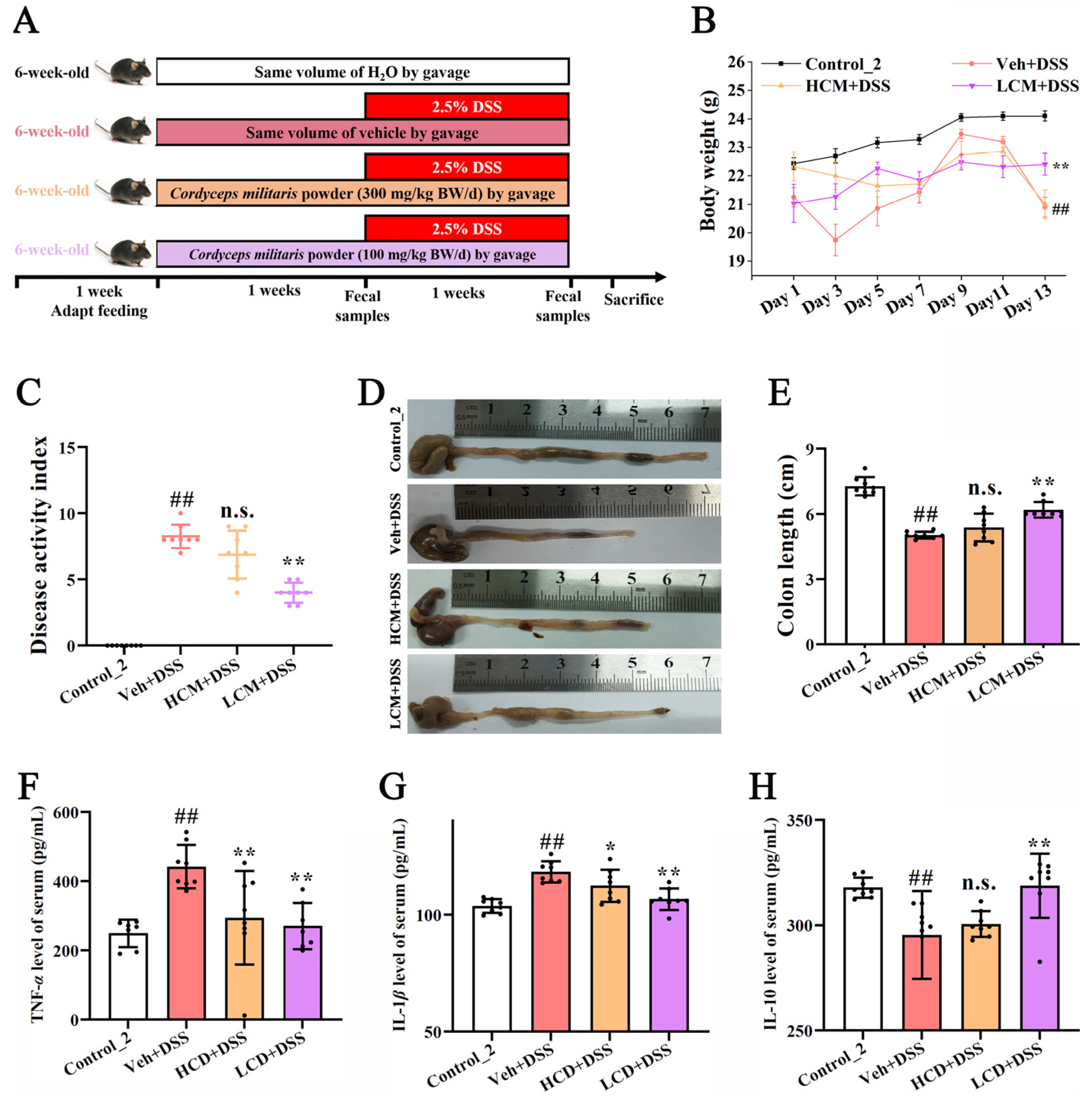
2.1. Animals and Treatments
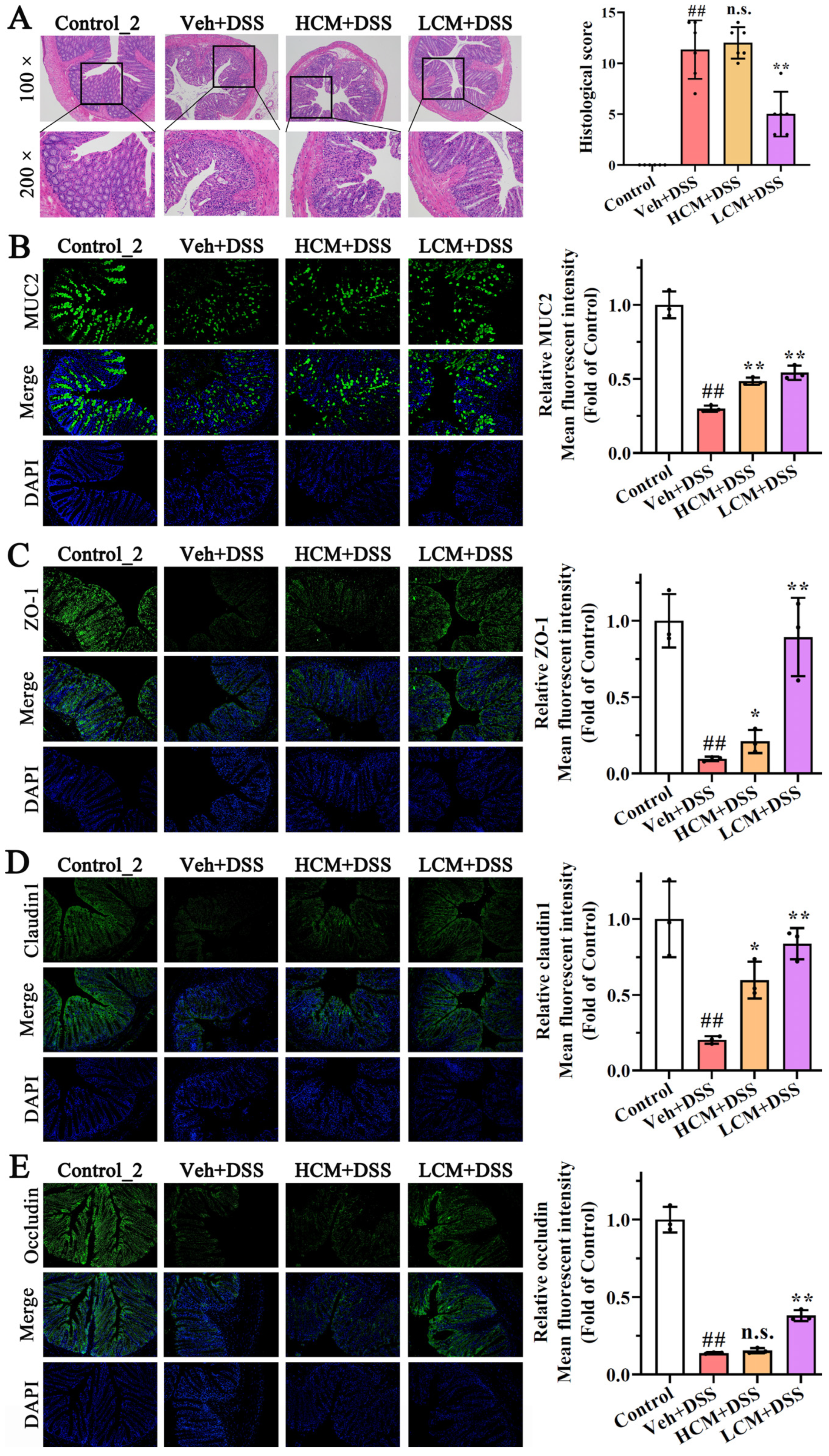
2.2. Histopathological Examination and Evaluation of Colons
2.3. Disease Activity Index and Histological Score
2.4. Cytokine Analysis of Serum
2.5. Immunofluorescence Assessment
2.6. 16S rRNA Gene Sequencing for Fecal Matter
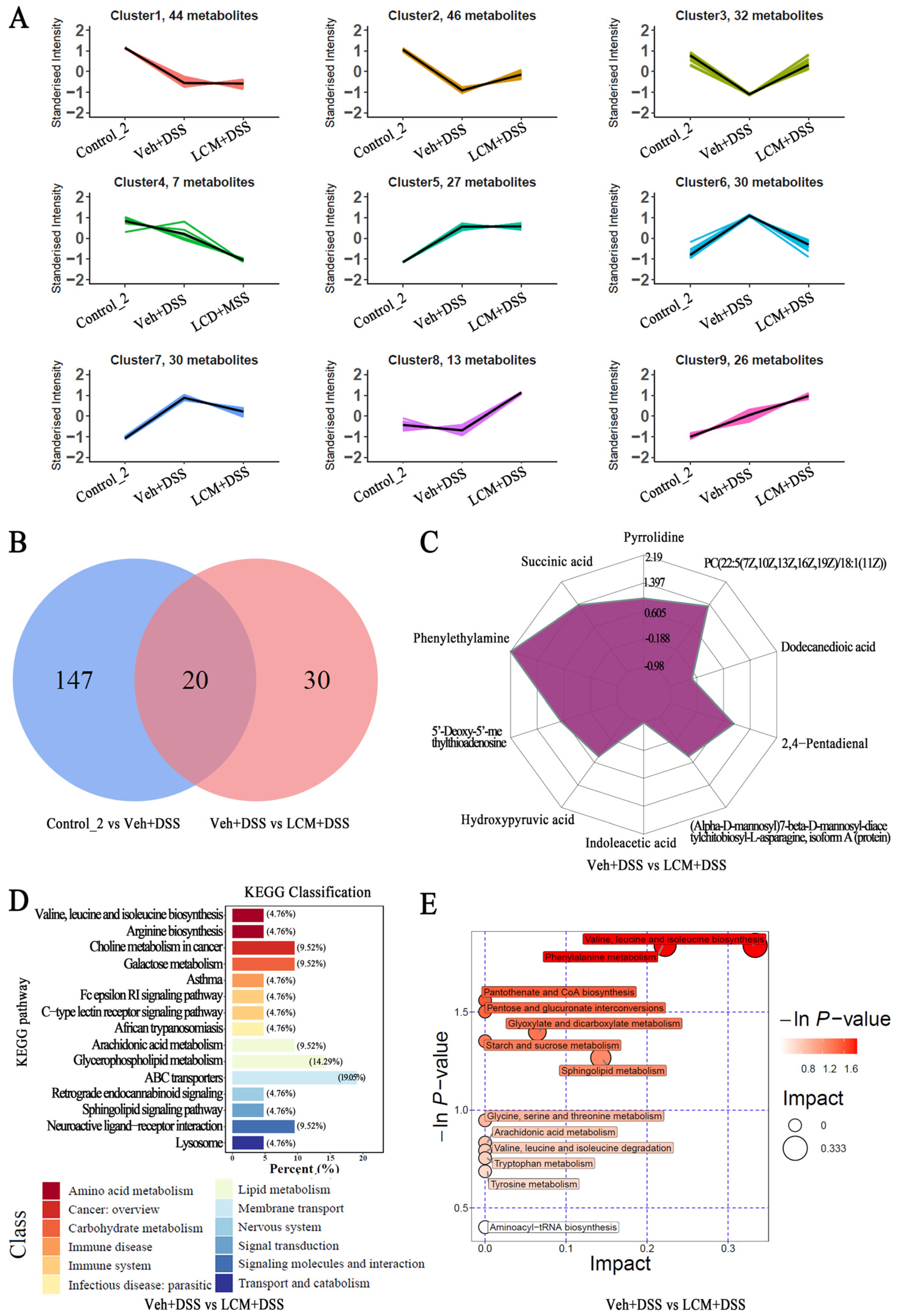
2.7. Untargeted Fecal Metabolomics Analysis
2.8. In Vitro Fermentation of Peptides and Polysaccharides from CM
2.9. Statistical Analysis
3. Results
3.1. CM Powder Alleviated DSS-Induced UC in Mice
3.2. CM Powder Enhanced Intestinal Mucosal Barrier Function
3.3. CM Powder Intervention Changed the Fecal Microbiota Composition
3.4. CM Powder Modulated the Fecal Metabolome in DSS-Injured Mice
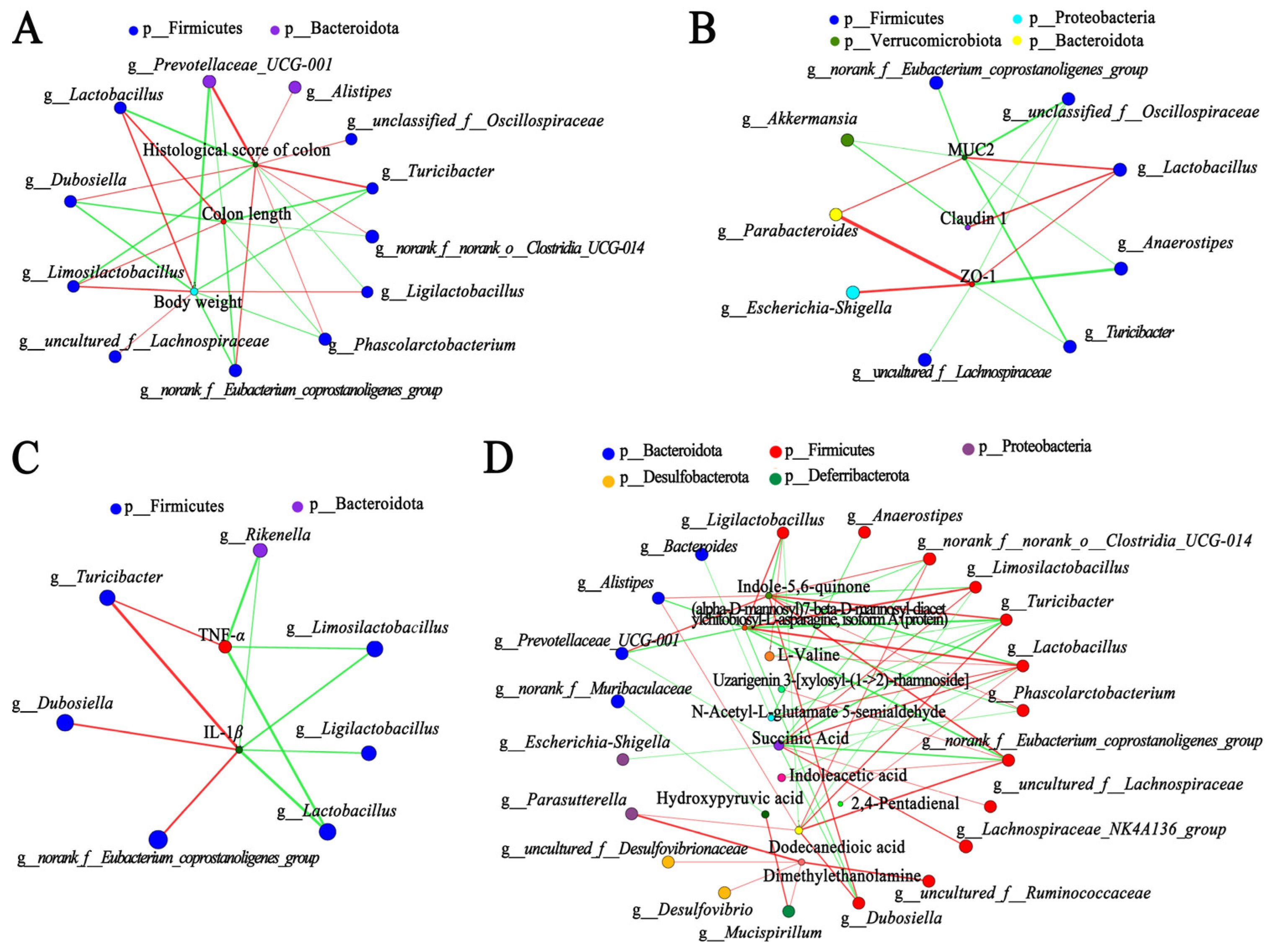
3.5. Correlation Analysis among Gut Microbiota, Metabolites, and Other Parameters in DSS-Injured Mice
3.6. Peptides and Polysaccharides of CM Changed the Gut Microbiota Structure of UC Patient’s Faeces
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.; Chan, F.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Shivaji, U.N.; Nardone, O.M.; Cannatelli, R.; Smith, S.C.; Ghosh, S.; Iacucci, M. Small molecule oral targeted therapies in ulcerative colitis. Lancet Gastroenterol. Hepatol. 2020, 5, 850–861. [Google Scholar] [CrossRef]
- Ferretti, F.; Cannatelli, R.; Monico, M.C.; Maconi, G.; Ardizzone, S. An Update on Current Pharmacotherapeutic Options for the Treatment of Ulcerative Colitis. J. Clin. Med. 2022, 11, 2302. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhao, J.; Xie, H.; Cai, M.; Yao, L.; Li, J.; Han, L.; Chen, W.; Yu, N.; Peng, D. Dendrobium huoshanense polysaccharides ameliorate ulcerative colitis by improving intestinal mucosal barrier and regulating gut microbiota. J. Funct. Food 2022, 96, 105231. [Google Scholar] [CrossRef]
- Zhou, F.; Lin, Y.; Chen, S.; Bao, X.; Fu, S.; Lv, Y.; Zhou, M.; Chen, Y.; Zhu, B.; Qian, C.; et al. Ameliorating role of Tetrastigma hemsleyanum polysaccharides in antibiotic-induced intestinal mucosal barrier dysfunction in mice based on microbiome and metabolome analyses. Int. J. Biol. Macromol. 2023, 241, 124419. [Google Scholar] [CrossRef]
- Zou, J.; Liu, C.; Jiang, S.; Qian, D.; Duan, J. Cross Talk between Gut Microbiota and Intestinal Mucosal Immunity in the Development of Ulcerative Colitis. Infect. Immun. 2021, 89, e1421. [Google Scholar] [CrossRef]
- Huang, L.; He, F.; Wu, B. Mechanism of effects of nickel or nickel compounds on intestinal mucosal barrier. Chemosphere 2022, 305, 135429. [Google Scholar]
- van der Post, S.; Jabbar, K.S.; Birchenough, G.; Arike, L.; Akhtar, N.; Sjovall, H.; Johansson, M.E.V.; Hansson, G.C. Structural weakening of the colonic mucus barrier is an early event in ulcerative colitis pathogenesis. Gut 2019, 68, 2142–2151. [Google Scholar] [CrossRef]
- Hill, J.H.; Round, J.L. SnapShot: Microbiota effects on host physiology. Cell 2021, 184, 2796. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Bhat, Z.F.; Gounder, R.S.; Ahmed, I.A.M.; Al-Juhaimi, F.Y.; Ding, Y.; Bekhit, A.E.D.A. Effect of Dietary Protein and Processing on Gut Microbiota—A Systematic Review. Nutrients 2022, 14, 453. [Google Scholar] [CrossRef]
- Yadav, D.; Negi, P.S. Chapter 30—Role of Mushroom Polysaccharides in Improving Gut Health and Associated Diseases; Bagchi, D., Downs, B.W., Eds.; Microbiome, Immunity, Digestive Health and Nutrition Academic Press: Houston, TX, USA, 2022; pp. 431–448. [Google Scholar]
- Wang, L.; Huang, Q.; Huang, Y.; Xie, J.; Qu, C.; Chen, J.; Zheng, L.; Yi, T.; Zeng, H.; Li, H. Comparison of protective effect of ordinary Cordyceps militaris and selenium-enriched Cordyceps militaris on triptolide-induced acute hepatotoxicity and the potential mechanisms. J. Funct. Food 2018, 46, 365–377. [Google Scholar] [CrossRef]
- Sun, Y.; Shao, Y.; Zhang, Z.; Wang, L.; Mariga, A.M.; Pang, G.; Geng, C.; Ho, C.; Hu, Q.; Zhao, L. Regulation of human cytokines by Cordyceps militaris. J. Food Drug Anal. 2014, 22, 463–467. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, Z.; Wu, S.; Wang, J.; Chen, M.; Liu, W.; Huang, A.; Zhang, J.; Wu, Q.; Ding, Y. Polysaccharides from Cordyceps militaris prevent obesity in association with modulating gut microbiota and metabolites in high-fat diet-fed mice. Food Res. Int. 2022, 157, 111197. [Google Scholar] [CrossRef]
- Wu, S.; Wu, Q.; Wang, J.; Li, Y.; Chen, B.; Zhu, Z.; Huang, R.; Chen, M.; Huang, A.; Xie, Y.; et al. Novel Selenium Peptides Obtained from Selenium-EnrichedCordyceps militaris Alleviate Neuroinflammation and Gut Microbiota Dysbacteriosis in LPS-Injured Mice. J. Agric. Food Chem. 2022, 70, 3194–3206. [Google Scholar] [CrossRef]
- Liu, X.; Dun, M.; Jian, T.; Sun, Y.; Wang, M.; Zhang, G.; Ling, J. Cordyceps militaris extracts and cordycepin ameliorate type 2 diabetes mellitus by modulating the gut microbiota and metabolites. Front. Pharmacol. 2023, 14, 1134429. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Huang, R.; Huang, A.; Wang, J.; Liu, W.; Wu, S.; Chen, M.; Chen, M.; Xie, Y.; Jiao, C.; et al. Polysaccharide from Agrocybe cylindracea prevents diet-induced obesity through inhibiting inflammation mediated by gut microbiota and associated metabolites. Int. J. Biol. Macromol. 2022, 209, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Chen, M.; Liao, X.; Huang, R.; Wang, J.; Xie, Y.; Hu, H.; Zhang, J.; Wu, Q.; Ding, Y. Protein hydrolysates from Pleurotus geesteranus obtained by simulated gastrointestinal digestion exhibit neuroprotective effects in H2O2-injured PC12 cells. J. Food Biochem. 2022, 46, e13879. [Google Scholar] [CrossRef] [PubMed]
- Soufli, I.; Toumi, R.; Rafa, H.; Touil-Boukoffa, C. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World J. Gastrointest. Pharmacol. Ther. 2016, 7, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wu, L.; Dou, C.; Guan, X.; Wu, H.; Liu, H.; Oz, H. Research Advance in Intestinal Mucosal Barrier and Pathogenesis of Crohn’s Disease. Gastroenterol. Res. Pract. 2016, 2016, 9686236–9686238. [Google Scholar] [CrossRef]
- Yu, X.; Zou, Y.; Zheng, Q.; Lu, F.; Li, D.; Guo, L.; Lin, J. Physicochemical, functional and structural properties of the major protein fractions extracted from Cordyceps militaris fruit body. Food Res. Int. 2021, 142, 110211. [Google Scholar] [CrossRef]
- Lu, Q.; Li, R.; Yang, Y.; Zhang, Y.; Zhao, Q.; Li, J. Ingredients with anti-inflammatory effect from medicine food homology plants. Food Chem. 2022, 368, 130610. [Google Scholar] [CrossRef]
- Hu, Q.; Yuan, B.; Wu, X.; Du, H.; Gu, M.; Han, Y.; Yang, W.; Song, M.; Xiao, H. Dietary Intake of Pleurotus eryngii Ameliorated Dextran-Sodium-Sulfate-Induced Colitis in Mice. Mol. Nutr. Food Res. 2019, 63, e1801265. [Google Scholar] [CrossRef]
- Li, M.; Ge, Q.; Du, H.; Jiang, P.; Bao, Z.; Chen, D.; Lin, S. Potential Mechanisms Mediating the Protective Effects of Tricholoma matsutake-Derived Peptides in Mitigating DSS-Induced Colitis. J. Agric. Food Chem. 2021, 69, 5536–5546. [Google Scholar] [CrossRef]
- Liu, F.; Wang, X.; Li, D.; Cui, Y.; Li, X. Apple polyphenols extract alleviated dextran sulfate sodium-induced ulcerative colitis in C57BL/6 male mice by restoring bile acid metabolism disorder and gut microbiota dysbiosis. Phytother. Res. 2021, 35, 1468–1485. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Dorofeyev, A.E.; Vasilenko, I.V.; Rassokhina, O.A.; Kondratiuk, R.B.; Amre, D. Mucosal Barrier in Ulcerative Colitis and Crohn’s Disease. Gastroenterol. Res. Pract. 2013, 2013, 431231–431239. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, Q.Y.; Wang, W.Z.; Chu, S.; Liu, X.X.; Liu, Y.J.; Tan, C.; Zhu, F.; Deng, S.J.; Dong, Y.L.; et al. Compound sophorae decoction enhances intestinal barrier function of dextran sodium sulfate induced colitis via regulating notch signaling pathway in mice. Biomed. Pharmacother. 2021, 133, 110937. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Dai, W.; Dong, M.; Dai, C.; Wu, S. MUC2 and related bacterial factors: Therapeutic targets for ulcerative colitis. EBioMedicine 2021, 74, 103751. [Google Scholar] [CrossRef]
- Van Klinken, B.J.; Van der Wal, J.W.; Einerhand, A.W.; Buller, H.A.; Dekker, J. Sulphation and secretion of the predominant secretory human colonic mucin MUC2 in ulcerative colitis. Gut 1999, 44, 387–393. [Google Scholar] [CrossRef]
- Seo, K.; Seo, J.; Yeun, J.; Choi, H.; Kim, Y.; Chang, S. The role of mucosal barriers in human gut health. Arch. Pharm. Res. 2021, 44, 325–341. [Google Scholar]
- Zeng, W.; He, D.; Xing, Y.; Liu, J.; Su, N.; Zhang, C.; Wang, Y.; Xing, X. Internal connections between dietary intake and gut microbiota homeostasis in disease progression of ulcerative colitis: A review. Food Sci. Hum. Wellness 2021, 10, 119–130. [Google Scholar] [CrossRef]
- Guo, X.Y.; Liu, X.J.; Hao, J.Y. Gut microbiota in ulcerative colitis: Insights on pathogenesis and treatment. J. Dig. Dis. 2020, 21, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Xie, L.; Huang, L. Regulation of host immune responses by Lactobacillus through aryl hydrocarbon receptors. Med. Microecol. 2023, 16, 100081. [Google Scholar] [CrossRef]
- Dhillon, P.; Singh, K. Therapeutic applications of probiotics in ulcerative colitis: An updated review. Pharmanutrition 2020, 13, 100194. [Google Scholar] [CrossRef]
- Nan, Q.; Ye, Y.; Tao, Y.; Jiang, X.; Miao, Y.; Jia, J.; Miao, J. Alterations in metabolome and microbiome signatures provide clues to the role of antimicrobial peptide KT2 in ulcerative colitis. Front. Microbiol. 2023, 14, 1027658. [Google Scholar] [CrossRef]
- Wan, J.; Yu, X.; Liu, J.; Li, J.; Ai, T.; Yin, C.; Liu, H.; Qin, R. A special polysaccharide hydrogel coated on Brasenia schreberi: Preventive effects against ulcerative colitis via modulation of gut microbiota. Food Funct. 2023, 14, 3564–3575. [Google Scholar] [CrossRef]
- Bunker, J.J.; Flynn, T.M.; Koval, J.C.; Shaw, D.G.; Meisel, M.; McDonald, B.D.; Ishizuka, I.E.; Dent, A.L.; Wilson, P.C.; Jabri, B.; et al. Innate and Adaptive Humoral Responses Coat Distinct Commensal Bacteria with Immunoglobulin A. Immunity 2015, 43, 541–553. [Google Scholar] [CrossRef]
- Lima, S.F.; Gogokhia, L.; Viladomiu, M.; Chou, L.; Putzel, G.; Jin, W.; Pires, S.; Guo, C.; Gerardin, Y.; Crawford, C.V.; et al. Transferable Immunoglobulin A–Coated Odoribacter splanchnicus in Responders to Fecal Microbiota Transplantation for Ulcerative Colitis Limits Colonic Inflammation. Gastroenterology 2022, 162, 166–178. [Google Scholar] [CrossRef]
- Peng, L.; Gao, X.; Nie, L.; Xie, J.; Dai, T.; Shi, C.; Tao, L.; Wang, Y.; Tian, Y.; Sheng, J. Astragalin Attenuates Dextran Sulfate Sodium (DSS)-Induced Acute Experimental Colitis by Alleviating Gut Microbiota Dysbiosis and Inhibiting NF-kappaB Activation in Mice. Front. Immunol. 2020, 11, 2058. [Google Scholar] [CrossRef]
- Gierynska, M.; Szulc-Dabrowska, L.; Struzik, J.; Mielcarska, M.B.; Gregorczyk-Zboroch, K.P. Integrity of the Intestinal Barrier: The Involvement of Epithelial Cells and Microbiota-A Mutual Relationship. Animals 2022, 12, 145. [Google Scholar] [CrossRef]
- Fang, J.; Wang, H.; Zhou, Y.; Zhang, H.; Zhou, H.; Zhang, X. Slimy partners: The mucus barrier and gut microbiome in ulcerative colitis. Exp. Mol. Med. 2021, 53, 772–787. [Google Scholar] [CrossRef]
- Qin, D.; Ma, Y.; Wang, Y.; Hou, X.; Yu, L. Contribution of Lactobacilli on Intestinal Mucosal Barrier and Diseases: Perspectives and Challenges of Lactobacillus casei. Life 2022, 12, 1910. [Google Scholar] [PubMed]
- Rodriguez-Pineiro, A.M.; Johansson, M.E. The colonic mucus protection depends on the microbiota. Gut Microbes 2015, 6, 326–330. [Google Scholar] [CrossRef]
- Zhao, B.; Xia, B.; Li, X.; Zhang, L.; Liu, X.; Shi, R.; Kou, R.; Liu, Z.; Liu, X. Sesamol Supplementation Attenuates DSS-Induced Colitis via Mediating Gut Barrier Integrity, Inflammatory Responses, and Reshaping Gut Microbiome. J. Agric. Food Chem. 2020, 68, 10697–10708. [Google Scholar] [CrossRef] [PubMed]
- Vanden Bussche, J.; Marzorati, M.; Laukens, D.; Vanhaecke, L. Validated High Resolution Mass Spectrometry-Based Approach for Metabolomic Fingerprinting of the Human Gut Phenotype. Anal. Chem. 2015, 87, 10927–10934. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Han, X.; Li, J.; Shi, R.; Liu, L.; Wang, K.; Liao, Y.; Jiang, H.; Zhang, Y.; Hu, J.; et al. Differential analysis of intestinal microbiota and metabolites in mice with dextran sulfate sodium-induced colitis. World J. Gastroenterol. WJG 2022, 28, 6109–6130. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wu, X.; Jiang, S.; Wang, Y.; Kong, D.; Zhao, Y.; Wang, W. The application of omics techniques to evaluate the effects of Tanshinone IIA on dextran sodium sulfate induced ulcerative colitis. Mol. Omics 2022, 18, 666–676. [Google Scholar] [CrossRef]
- Yuan, Z.; Yang, L.; Zhang, X.; Ji, P.; Hua, Y.; Wei, Y. Mechanism of Huang-lian-Jie-du decoction and its effective fraction in alleviating acute ulcerative colitis in mice: Regulating arachidonic acid metabolism and glycerophospholipid metabolism. J. Ethnopharmacol. 2020, 259, 112872. [Google Scholar] [CrossRef]
- Zhu, S.; Han, M.; Liu, S.; Fan, L.; Shi, H.; Li, P. Composition and diverse differences of intestinal microbiota in ulcerative colitis patients. Front. Cell. Infect. Microbiol. 2022, 12, 953962. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight Loss | Stool Consistency for Diarrhea | Fecal Occult Blood | Score |
|---|---|---|---|
| None | Normal | Normal | 0 |
| 1–5% | Soft but still formed | Melena and negative blood | 1 |
| 6–10% | Soft | Positive blood | 2 |
| 11–15% | Soft and wet | Blood stains can be seen on the stool | 3 |
| >15% | Watery diarrhea | Rectal bleeding | 4 |
| Inflammation | Mucosal Damage | Crypt Loss | Pathological Change Range | Score |
|---|---|---|---|---|
| None | None | None | None | 0 |
| Mild | Mucus layer | 1–33% | 1–25% | 1 |
| Moderate | Submucosa | 34–66% | 26–50% | 2 |
| Severe | Muscular and serosa | 67–100% + intact epithelium | 51–75% | 3 |
| - | - | 100% with epithelium lose | 76–100% | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Wu, Z.; Chen, Y. Effect of Cordyceps militaris Powder Prophylactic Supplementation on Intestinal Mucosal Barrier Impairment and Microbiota-Metabolites Axis in DSS-Injured Mice. Nutrients 2023, 15, 4378. https://doi.org/10.3390/nu15204378
Wu S, Wu Z, Chen Y. Effect of Cordyceps militaris Powder Prophylactic Supplementation on Intestinal Mucosal Barrier Impairment and Microbiota-Metabolites Axis in DSS-Injured Mice. Nutrients. 2023; 15(20):4378. https://doi.org/10.3390/nu15204378
Chicago/Turabian StyleWu, Shujian, Zaoxuan Wu, and Ye Chen. 2023. "Effect of Cordyceps militaris Powder Prophylactic Supplementation on Intestinal Mucosal Barrier Impairment and Microbiota-Metabolites Axis in DSS-Injured Mice" Nutrients 15, no. 20: 4378. https://doi.org/10.3390/nu15204378
APA StyleWu, S., Wu, Z., & Chen, Y. (2023). Effect of Cordyceps militaris Powder Prophylactic Supplementation on Intestinal Mucosal Barrier Impairment and Microbiota-Metabolites Axis in DSS-Injured Mice. Nutrients, 15(20), 4378. https://doi.org/10.3390/nu15204378