
Alpha-Glucosidase Inhibitory Peptides: Sources, Preparations, Identifications, and Action Mechanisms
Abstract
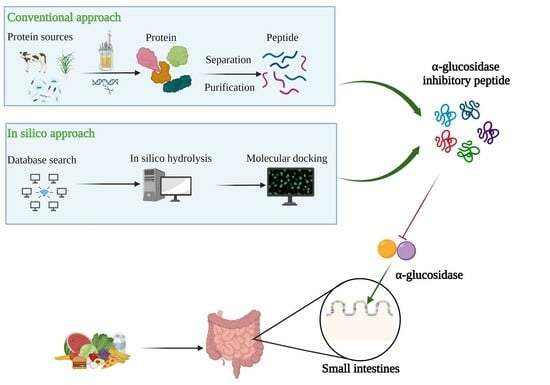
:
1. Introduction
2. Production, Purification, and Characterization of Bioactive Peptides
2.1. Conventional Approach
2.2. Bioinformatics-Assisted Methods
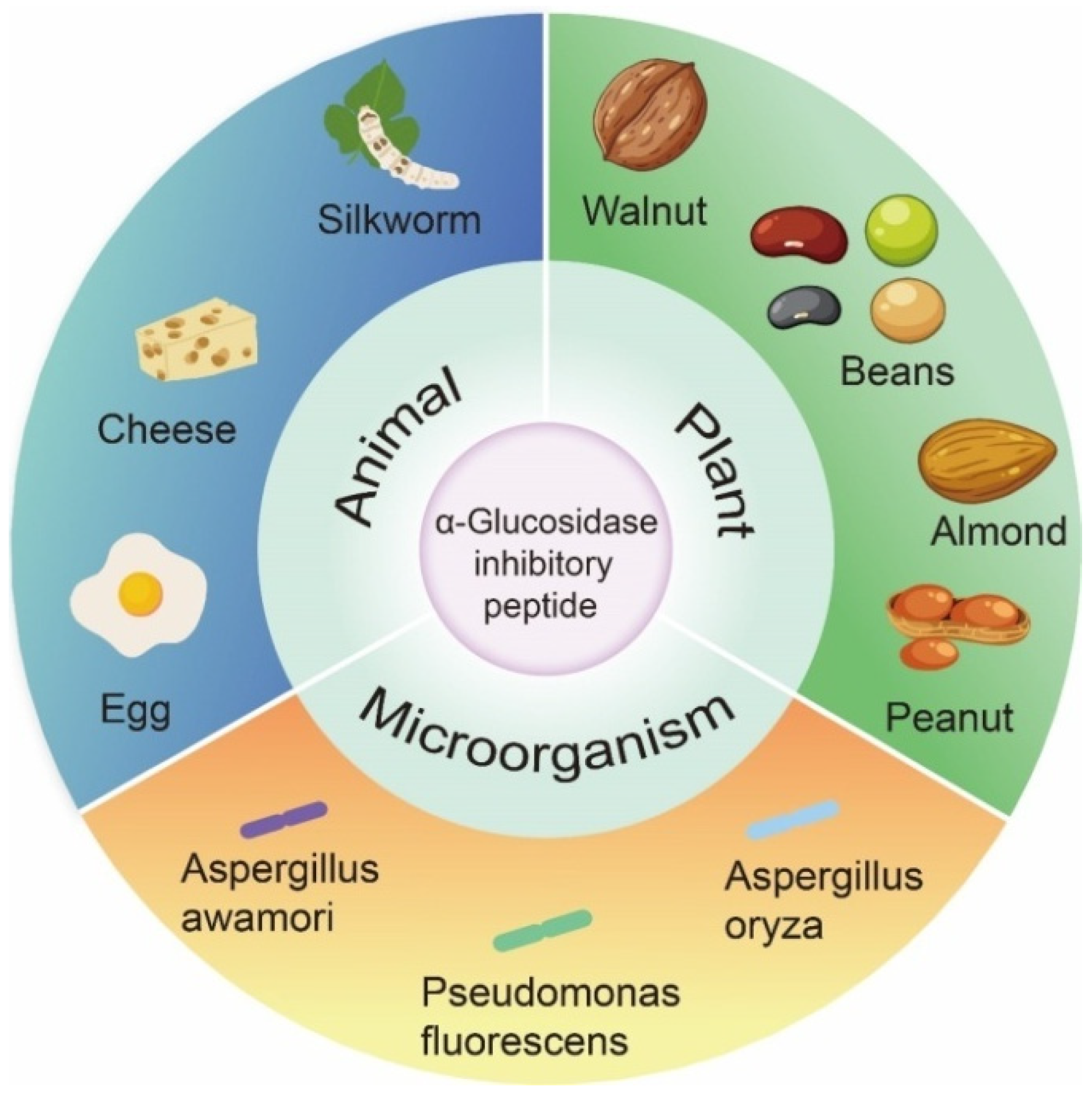
3. Source and Structural Characteristics of α-Glucosidase Inhibitory Peptide
3.1. Animal Origin
3.2. Plant Origin
3.3. Microbial Origin
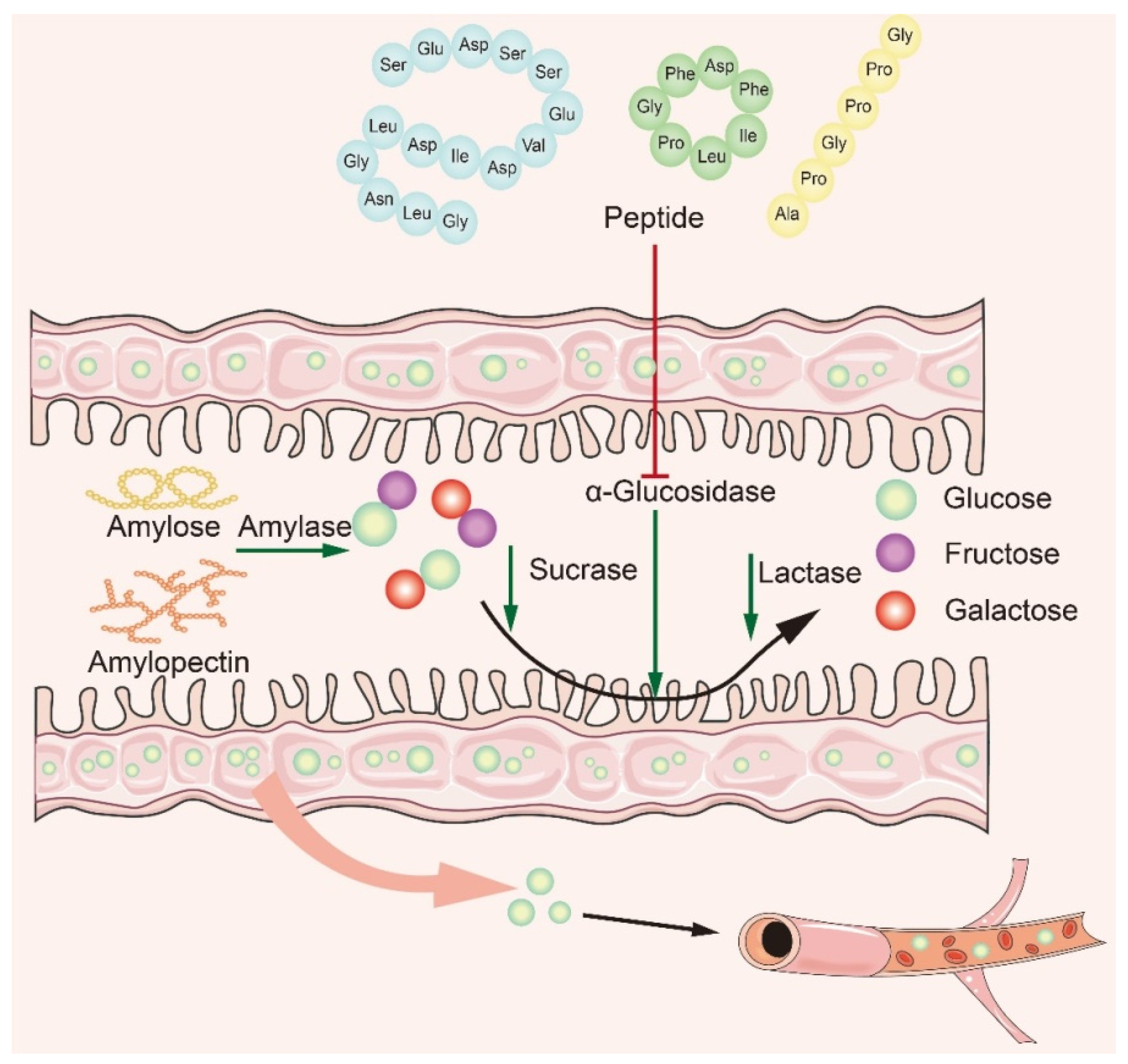
4. Mechanism of α-Glucosidase Inhibitory Peptide
5. Bioavailability of α-Glucosidase Inhibitory Peptides
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACE | Angiotension Converting Enzyme |
| DPP-IV | Dipeptidyl Peptidase IV |
| ESI-MA | Electrospray Ionization Mass Spectrometry |
| GDM | Gestational Diabetes Mellitus |
| IC50 | Half Maximal Inhibitory Concentration |
| LC-MS | Liquid Chromatography–Mass Spectrometry |
| MW | Molecular Weight |
| MS | Mass Spectrum |
| MALDI-TOF | Matrix-Assisted Laser Desorption Time-of-Flight Mass Spectrometry |
| Q-TOFMS | Time-of-Flight Tandem Mass Spectrometer |
| RP-HPLC | Reverse Phase–High Performance Liquid Chromatography |
| SEC | Size Exclusion Chromatography |
| SDS-PAGE | Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis |
| T1DM | Type 1 Diabetes Mellitus |
| T2DM | Type 2 Diabetes Mellitus |
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Dehdashtian, E.; Mehrzadi, S.; Yousefi, B.; Hosseinzadeh, A.; Reiter, R.J.; Safa, M.; Ghaznavi, H.; Naseripour, M. Diabetic retinopathy pathogenesis and the ameliorating effects of melatonin; involvement of autophagy, inflammation and oxidative stress. Life Sci. 2018, 193, 20–33. [Google Scholar]
- Simó-Servat, O.; Hernández, C.; Simó, R. Diabetic retinopathy in the context of patients with diabetes. Ophthalmic Res. 2019, 62, 211–217. [Google Scholar] [CrossRef]
- Sandholm, N.; Groop, P.H. Genetic basis of diabetic kidney disease and other diabetic complications. Curr. Opin. Genet. Dev. 2018, 50, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Harding, J.L.; Pavkov, M.E.; Magliano, D.J.; Shaw, J.E.; Gregg, E.W. Global trends in diabetes complications: A review of current evidence. Diabetologia 2019, 62, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Gloster, T.M.; Davies, G.J. Glycosidase inhibition: Assessing mimicry of the transition state. Org. Biomol. Chem. 2010, 8, 305–320. [Google Scholar] [CrossRef]
- Chiarelli, F.; Marzio, D.D. Peroxisome proliferator-activated receptor-γ agonists and diabetes: Current evidence and future perspectives. Vasc. Health Risk Man. 2008, 4, 297–304. [Google Scholar]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.H.; Gaspar, A.R.M. Structural properties of bioactive peptides with α-glucosidase inhibitory activity. Chem. Biol. Drug Des. 2018, 91, 370–379. [Google Scholar] [CrossRef]
- Zhang, B.W.; Li, X.; Sun, W.L.; Xing, Y.; Xiu, Z.L.; Zhuang, C.L.; Dong, Y.S. Dietary flavonoids and acarbose synergistically inhibit α-glucosidase and lower postprandial blood glucose. J. Agric. Food Chem. 2017, 65, 8319–8330. [Google Scholar] [CrossRef]
- Ha, T.J.; Song, S.B.; Ko, J.; Park, C.H.; Ko, J.M.; Choe, M.E.; Kwak, D.Y.; Lee, J.H. Isolation and identification of α-glucosidase inhibitory constituents from the seeds of vigna nakashimae: Enzyme kinetic study with active phytochemical. Food Chem. 2018, 266, 483–489. [Google Scholar] [CrossRef]
- Aslam, M.Z.; Aslam, M.S.; Firdos, S.; Ghous, G.; Firdos, G.; Zhao, H.F.; Zhang, B.L. Role of bioactive peptides in reducing the severity of hypertension with the inhibition of ACE. Int. J. Pept. Res. Ther. 2019, 25, 1639–1649. [Google Scholar] [CrossRef]
- Yan, J.; Zhao, J.G.; Yang, R.J.; Zhao, W. Bioactive peptides with antidiabetic properties: A review. Int. J. Food Sci. Technol. 2019, 54, 1909–1919. [Google Scholar] [CrossRef]
- Wu, Q.; Luo, F.; Wang, X.L.; Lin, Q.; Liu, G.Q. Angiotensin I-converting enzyme inhibitory peptide: An emerging candidate for vascular dysfunction therapy. Crit. Rev. Biotechnol. 2022, 42, 736–755. [Google Scholar] [CrossRef]
- Liu, J.; Wu, Q.; Yang, T.; Yang, F.; Guo, T.; Zhou, Y.; Han, S.; Luo, Y.; Guo, T.; Luo, F.; et al. Bioactive peptide F2d isolated from rice residue exerts antioxidant effects via Nrf2 signaling pathway. Oxid. Med. Cell Longev. 2021, 2021, 2637577. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tomé, S.; Sanchón, J.; Recio, I.; Hernandez-Ledesma, B. Transepithelial transport of lunasin and derived peptides: Inhibitory effects on the gastrointestinal cancer cells viability. J. Food Compos. Anal. 2018, 68, 101–110. [Google Scholar] [CrossRef]
- Sun, S.; Niu, H.; Yang, T.; Lin, Q.; Luo, F.; Ma, M. Antioxidant and anti-fatigue activities of egg white peptides prepared by pepsin digestion. J. Sci. Food Agric. 2014, 94, 3195–3200. [Google Scholar] [CrossRef]
- Mora, L.; Gallego, M.; Reig, M.; Toldra, F. Challenges in the quantitation of naturally generated bioactive peptides in processed meats. Trends Food Sci. Technol. 2017, 69, 306–314. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Y.P.; Liu, Y.H.; Ruan, R. Bioactive peptides derived from traditional Chinese medicine and traditional Chinese food: A review. Food Res. Int. 2016, 89, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.M.; Fan, X.; Wang, D.P.; Li, X.A.; Wang, X.Y.; Yang, J.Y.; Qiu, C.G.; Liu, X.L.; Pang, G.X.; Abra, R.; et al. Study on preparation of chickpea peptide and its effect on blood glucose. Front. Nutr. 2022, 9, 988628. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Liu, X.Y.; Wang, W.; Wang, J.H.; Li, X.; Sun, S.L. Preparation and Identification of Peptides with α-Glucosidase Inhibitory Activity from Shiitake Mushroom (Lentinus edodes) Protein. Foods 2023, 12, 2534. [Google Scholar] [CrossRef] [PubMed]
- Espitia, P.J.P.; Soares, N.D.F.; Coimbra, J.S.D.; de Andrade, N.J.; Cruz, R.S.; Medeiros, E.A.A. Bioactive Peptides: Synthesis, Properties, and Applications in the Packaging and Preservation of Food. Compr. Rev. Food Sci. Food Saf. 2012, 11, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, V.; Venkatesan, M.; Ramachandran, S.; Sundaresan, U. Bioactive peptides from marine ascidians and future drug development—A review. Int. J. Pept. Res. Ther. 2018, 24, 13–18. [Google Scholar] [CrossRef]
- Huang, F.J.; Wu, T. Purification and characterization of a new peptide (s-8300) from shark liver. J. Food Biochem. 2010, 34, 962–970. [Google Scholar] [CrossRef]
- Olagunju, A.I.; Omoba, O.S.; Enujiugha, V.N.; Alashi, A.M.; Aluko, R.E. Thermoase-hydrolysed pigeon pea protein and its membrane fractions possess in vitro bioactive properties (antioxidative, antihypertensive, and antidiabetic). J. Food Biochem. 2021, 45, e13429. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.F.; Lai, X.H.; Wu, X.D.; Wang, H.Y.; Weng, N.H.; Lu, J.; Lyu, M.; Wang, S.J. Isolation of a novel anti-diabetic α-glucosidase oligo-peptide inhibitor from fermented rice bran. Foods 2023, 12, 183. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.G.; Wang, X.B.; Ma, L.; Kong, L.Y. Gypsophin: A novel α-glucosidase inhibitory cyclic peptide from the roots of gypsophila oldhamiana. Bioorg. Med. Chem. Lett. 2007, 17, 4460–4463. [Google Scholar] [CrossRef] [PubMed]
- Zambrowicz, A.; Pokora, M.; Setner, B.; Dąbrowska, A.; Szołtysik, M.; Babij, K.; Szewczuk, Z.; Trziszka, T.; Lubec, G.; Chrzanowska, J. Multifunctional peptides derived from an egg yolk protein hydrolysate: Isolation and characterization. Amino Acids 2015, 47, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.T.; Lin, L.K.; Li, T.T.; Li, C.; Chen, B.; Shen, Y.H. Separation, identification, and design of α-glucosidase inhibitory peptides based on the molecular mechanism from Paeonia ostii ‘Feng Dan’seed protein. J. Food Sci. 2022, 87, 4892–4904. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, Y.Y.; Yu, T.T.; He, J.T.; Cui, J.; Wang, J.N.; Cheng, X.N.; Fan, J.F. Oat globulin peptides regulate antidiabetic drug targets and glucose transporters in Caco-2 cells. J. Funct. Foods 2018, 42, 12–20. [Google Scholar] [CrossRef]
- Cox, J. Prediction of peptide mass spectral libraries with machine learning. Nat. Biotechnol. 2023, 41, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Neagu, A.N.; Jayathirtha, M.; Baxter, E.; Donnelly, M.; Petre, B.A.; Darie, C.C. Applications of tandem mass spectrometry (MS/MS) in protein analysis for biomedical research. Molecules 2022, 27, 2411. [Google Scholar] [CrossRef]
- Qiu, X.I.; Ruterbories, K.J.; Ji, Q.C.; Jenkins, G.J. Signature peptide selection workflow for biomarker quantification using LC–MS-based targeted proteomics. Bioanalysis 2023, 15, 295–300. [Google Scholar] [CrossRef]
- Yan, Y.Q.; Wang, L.; Wang, H. Functional peptides from one-bead one-compound high-throughput screening technique. Chem. Res. Chin. Univ. 2023, 39, 83–91. [Google Scholar] [CrossRef]
- Guo, H.; Hao, Y.; Yang, X.; Ren, G.; Richel, A. Exploration on bioactive properties of quinoa protein hydrolysate and peptides: A review. Crit. Rev. Food Sci. 2023, 63, 2896–2909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.L.; Liu, L.L.; Gao, L.; Zhao, Y.H.; Shi, N.; Wang, H.B.; Xu, H.Y. Bioinformatics and computer simulation approaches to the discovery and analysis of bioactive peptides. Curr. Pharm. Biotechnol. 2022, 23, 1541–1555. [Google Scholar]
- Peredo-Lovillo, A.; Hernández-Mendoza, A.; Vallejo-Cordoba, B.; Romero-Luna, H.E. Conventional and in silico approaches to select promising food-derived bioactive peptides: A review. Food Chem. X 2022, 13, 100183. [Google Scholar] [CrossRef] [PubMed]
- Mudgil, P.; Kamal, H.; Kilari, B.P.; Salim, M.A.S.M.; Gan, C.Y.; Maqsood, S. Simulated gastrointestinal digestion of camel and bovine casein hydrolysates: Identification and characterization of novel anti-diabetic bioactive peptides. Food Chem. 2021, 353, 129374. [Google Scholar] [CrossRef] [PubMed]
- Baba, W.N.; Mudgil, P.; Kamal, H.; Kilari, B.P.; Gan, C.Y.; Maqsood, S. Identification and characterization of novel α-amylase and α-glucosidase inhibitory peptides from camel whey proteins. J. Dairy Sci. 2021, 104, 1364–1377. [Google Scholar] [CrossRef]
- Yu, Z.P.; Yin, Y.G.; Zhao, W.Z.; Yu, Y.D.; Liu, B.Q.; Liu, J.B.; Chen, F. Novel peptides derived from egg white protein inhibiting alpha-glucosidase. Food Chem. 2011, 129, 1376–1382. [Google Scholar] [CrossRef]
- Yu, Z.P.; Yin, Y.G.; Zhao, W.Z.; Liu, J.B.; Chen, F. Anti-diabetic activity peptides from albumin against α-glucosidase and α-amylase. Food Chem. 2012, 135, 2078–2085. [Google Scholar] [CrossRef]
- Zambrowicz, A.; Eckert, E.; Pokora, M.; Bobak, L.; Dabrowska, A.; Szoltysik, M.; Trzoszka, T.; Chrzanowska, J. Antioxidant and antidiabetic activities of peptides isolated from a hydrolysate of an egg-yolk protein by-product prepared with a proteinase from Asian pumpkin (Cucurbita ficifolia). Rsc. Adv. 2015, 5, 10460–10467. [Google Scholar] [CrossRef]
- Martini, S.; Solieri, L.; Cattivelli, A.; Pizzamiglio, V.; Tagliazucchi, D. An integrated peptidomics and in silico approach to identify novel anti-diabetic peptides in Parmigiano-Reggiano cheese. Biology 2021, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.Q.; Zhao, Q.; Wang, D.D.; Fan, Y.Z.; Shi, Y.A.; Huang, A.X. Novel ACE inhibitory, antioxidant and α-glucosidase inhibitory peptides identified from fermented rubing cheese through peptidomic and molecular docking. LWT-Food Sci. Technol. 2022, 159, 113196. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, H.S.; Choi, J.W.; Ra, K.S.; Kim, J.M.; Suh, H.J. Novel tripeptides with α-glucosidase inhibitory activity isolated from silk cocoon hydrolysate. J. Agric. Food Chem. 2011, 59, 11522–11525. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, N.; Wang, W.; Wang, J.H.; Zhu, Z.Y.; Lia, X. Molecular mechanisms of novel peptides from silkworm pupae that inhibit α-glucosidase. Peptides 2016, 76, 45–50. [Google Scholar] [CrossRef]
- Xie, F.; Wang, S.Y.; Zhang, L.; Wu, J.H.; Wang, Z.W. Investigating inhibitory activity of novel synthetic sericin peptide on α-D-glucosidase: Kinetics and interaction mechanism study using a docking simulation. J. Sci. Food Agric. 2018, 98, 1502–1510. [Google Scholar] [CrossRef] [PubMed]
- Ramadhan, A.H.; Nawas, T.; Zhang, X.W.; Pembe, W.M.; Xia, W.S.; Xu, Y.S. Purification and identification of a novel antidiabetic peptide from Chinese giant salamander (Andrias davidianus) protein hydrolysate against α-amylase and α-glucosidase. Int. J. Food Prop. 2017, 20, S3360–S3372. [Google Scholar] [CrossRef]
- Zhou, M.; Ren, G.Y.; Zhang, B.; Ma, F.L.; Fan, J.L.; Qiu, Z.J. Screening and identification of a novel antidiabetic peptide from collagen hydrolysates of Chinese giant salamander skin: Network pharmacology, inhibition kinetics and protection of IR-HepG2 cells. Food Funct. 2022, 13, 3329–3342. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, E.; Karaś, M.; Baraniak, B.; Jakubczyk, A. Evaluation of ACE, α-glucosidase, and lipase inhibitory activities of peptides obtained by in vitro digestion of selected species of edible insects. Eur. Food Res. Technol. 2020, 246, 1361–1369. [Google Scholar] [CrossRef]
- Matsui, T.; Oki, T.; Osajima, Y. Isolation and identification of peptidic α-glucosidase inhibitors derived from sardine muscle hydrolysate. Z. Naturforschung C 1999, 54, 259–263. [Google Scholar] [CrossRef]
- Huang, T.H.; Liu, P.Y.; Lin, Y.L.; Tsai, J.S. Hypoglycemic peptide-enriched hydrolysates of corbicula fluminea and chlorella sorokiniana possess synergistic hypoglycemic activity through inhibiting α-glucosidase and dipeptidyl peptidase-4 activity. J. Sci. Food Agric. 2022, 102, 716–723. [Google Scholar] [CrossRef]
- Wang, R.C.; Zhao, H.X.; Pan, X.X.; Orfila, C.; Lu, W.H.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of α-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Deng, Y.J.; Xie, P.J.; Liu, L.J.; Zhang, C.H.; Cheng, J.; Zhang, Y.; Liu, Y.; Huang, L.X.; Jiang, J.X. Novel bioactive peptides from ginkgo biloba seed protein and evaluation of their α-glucosidase inhibition activity. Food Chem. 2023, 404, 134481. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.Z.; Yan, H.; He, R.H.; Ma, Y.K. Purification and a molecular docking study of α-glucosidase-inhibitory peptides from a soybean protein hydrolysate with ultrasonic pretreatment. Eur. Food Res. Technol. 2018, 244, 1995–2005. [Google Scholar] [CrossRef]
- Mousavi, B.; Azizi, M.H.; Abbasi, S. Antidiabetic bio-peptides of soft and hard wheat glutens. Food Chem. Mol. Sci. 2022, 4, 100104. [Google Scholar]
- Zhao, B.L.; Su, K.Y.; Mao, X.L.; Zhang, X.W. Separation and identification of enzyme inhibition peptides from dark tea protein. Bioorg. Chem. 2020, 99, 103772. [Google Scholar] [CrossRef] [PubMed]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Hernández-Ledesma, B. Release of dipeptidyl peptidase IV, α-amylase and α-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa Willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef]
- Ujiroghene, O.J.; Liu, L.; Zhang, S.W.; Lu, J.; Pang, X.Y.; Lv, J.P. α-Glucosidase and ACE dual inhibitory protein hydrolysates and peptide fractions of sprouted quinoa yoghurt beverages inoculated with Lactobacillus casei. Food Chem. 2019, 299, 124985. [Google Scholar]
- Fu, Y.X.; Liu, Z.Y.; Wang, H.; Zhang, F.; Guo, S.; Shen, Q. Comparison of the generation of α-glucosidase inhibitory peptides derived from prolamins of raw and cooked foxtail millet: In vitro activity, de novo sequencing, and in silico docking. Food Chem. 2023, 411, 135378. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liang, K.; Jin, Y.Q.; Zhang, M.M.; Chen, Y.; Wu, H.; Lai, F.R. Identification and characterization of two novel α-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Liu, W.; Li, H.; Wen, Y.; Liu, Y.; Wang, J.; Sun, B. Molecular mechanism for the α-glucosidase inhibitory effect of wheat germ peptides. J. Agric. Food Chem. 2021, 69, 15231–15239. [Google Scholar] [CrossRef]
- Hu, S.F.; Fan, X.D.; Qi, P.; Zhang, X.W. Identification of anti-diabetes peptides from spirulina platensis. J. Funct. Foods 2019, 56, 333–341. [Google Scholar] [CrossRef]
- Wang, J.; Wu, T.; Fang, L.; Liu, C.L.; Liu, X.T.; Li, H.M.; Shi, J.H.; Li, M.H.; Min, W.H. Anti-diabetic effect by walnut (Juglans mandshurica Maxim.)-derived peptide LPLLR through inhibiting α-glucosidase and α-amylase, and alleviating insulin resistance of hepatic HepG2 cells. J. Funct. Foods 2020, 69, 103944. [Google Scholar] [CrossRef]
- Yang, X.Y.; Wang, D.; Dai, Y.Y.; Zhao, L.P.; Wang, W.T.; Ding, X.Z. Identification and molecular binding mechanism of novel α-glucosidase inhibitory peptides from hot-pressed peanut meal protein hydrolysates. Foods 2023, 12, 663. [Google Scholar] [CrossRef] [PubMed]
- Mojica, L.; De Mejía, E.G. Optimization of enzymatic production of anti-diabetic peptides from black bean (Phaseolus vulgaris L.) proteins, their characterization and biological potential. Food Funct. 2016, 7, 713–727. [Google Scholar] [CrossRef]
- Quintero-Soto, M.F.; Chávez-Ontiveros, J.; Garzón-Tiznado, J.A.; Salazar-Salas, N.Y.; Pineda-Hidalgo, K.V.; Delgado-Vargas, F.; Lopez-Valenzuela, J.A. Characterization of peptides with antioxidant activity and antidiabetic potential obtained from chickpea (Cicer arietinum L.) protein hydrolyzates. J. Food Sci. 2021, 86, 2962–2977. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.P.; Wu, F.H.; He, Z.P.; Fang, X.Z.; Liu, X.Q. Optimization and molecular mechanism of novel α-glucosidase inhibitory peptides derived from camellia seed cake through enzymatic hydrolysis. Foods 2023, 12, 393. [Google Scholar] [CrossRef]
- Kang, M.G.; Yi, S.H.; Lee, J.S. Production and characterization of a new α-glucosidase inhibitory peptide from aspergillus oryzae N159-1. Mycobiology 2013, 41, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Lozano-González, M.; Ovalle-Magallanes, B.; Rangel-Grimaldo, M.; De la Torre-Zavala, S.; Noriega, L.G.; Tovar-Palacio, C.; Tovar, A.R.; Mata, R. Antidiabetic in vitro and in vivo evaluation of cyclodipeptides isolated from Pseudomonas fluorescens IB-MR-66e. New J. Chem. 2019, 43, 7756–7762. [Google Scholar] [CrossRef]
- Qiu, L.Y.; Deng, Z.Y.; Zhao, C.D.; Xiao, T.; Weng, C.; Li, J.; Zheng, L.F. Nutritional composition and proteomic analysis of soft-shelled turtle (Pelodiscus sinensis) egg and identification of oligopeptides with alpha-glucosidase inhibitory activity. Food Res. Int. 2021, 145, 110414. [Google Scholar] [CrossRef]
- Li, N.; Qiu, L.Y.; Song, H.L.; Xiao, T.; Luo, T.; Deng, Z.Y.; Zheng, L.F. Inhibition mechanism of oligopeptides from soft-shelled turtle egg against α-glucosidase and their gastrointestinal digestive properties. J. Food Biochem. 2022, 46, e14328. [Google Scholar] [CrossRef]
- Wang, X.F.; Fan, Y.Z.; Xu, F.R.; Xie, J.; Gao, X.Y.; Li, L.F.; Tian, Y.; Sheng, J. Characterization of the structure, stability, and activity of hypoglycemic peptides from Moringa oleifera seed protein hydrolysates. Food Funct. 2022, 13, 3481–3494. [Google Scholar] [CrossRef] [PubMed]
- Fadimu, G.J.; Farahnaky, A.; Gill, H.; Olalere, O.A.; Gan, C.Y.; Truong, T. In-silico analysis and antidiabetic effect of α-amylase and α-glucosidase inhibitory peptides from lupin protein hydrolysate: Enzyme-peptide interaction study using molecular docking approach. Foods 2022, 11, 3375. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Gao, T.; Hou, Y.K.; Li, D.; Fu, L. Identification and characterization of two novel α-glucosidase inhibitory peptides from almond (Armeniaca sibirica) oil manufacture residue. LWT Food Sci. Technol. 2020, 134, 11021. [Google Scholar] [CrossRef]
- Pritchard, S.R.; Phillips, M.; Kailasapathy, K. Identification of bioactive peptides in commercial Cheddar cheese. Food Res. Int. 2010, 43, 1545–1548. [Google Scholar] [CrossRef]
- Fang, Y.W.; Wang, S.Y.; Wu, J.H.; Zhang, L.; Wang, Z.W.; Gan, L.; He, J.J.; Shi, H.M.; Hou, J.L. The kinetics and mechanism of α-glucosidase inhibition by F5-SP, a novel compound derived from sericin peptides. Food Funct. 2017, 8, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Le, T.K.D.; Danova, A.; Aree, T.; Duong, T.H.; Koketsu, M.; Ninomiya, M.; Sawada, Y.; Kamsri, P.; Pungpo, P.; Chavasiri, W. α-Glucosidase inhibitors from the stems of knema globularia. J. Nat. Prod. 2022, 85, 776–786. [Google Scholar] [CrossRef]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2022, 21, 1049–1079. [Google Scholar] [CrossRef]
- Teng, H.; Chen, L. α-Glucosidase and α-amylase inhibitors from seed oil: A review of liposoluble substance to treat diabetes. Crit. Rev. Food Sci. 2017, 57, 3438–3448. [Google Scholar] [CrossRef] [PubMed]
- Famuwagun, A.A.; Alashi, A.M.; Gbadamosi, O.S.; Taiwo, K.A.; Oyedele, D.; Adebooye, O.C.; Aluko, R.E. Antioxidant and enzymes inhibitory properties of amaranth leaf protein hydrolyzates and ultrafiltration peptide fractions. J. Food Biochem. 2021, 45, e13396. [Google Scholar] [CrossRef]
- Chen, N.; Yang, H.; Sun, Y.; Niu, J.; Liu, S.Y. Purification and identification of antioxidant peptides from walnut (Juglans regia L.) protein hydrolysates. Peptides 2012, 38, 344–349. [Google Scholar] [CrossRef]
- Dong, Z.Y.; Li, M.Y.; Tian, G.; Zhang, T.H.; Ren, H.; Quek, S.Y. Effects of ultrasonic pretreatment on the structure and functionality of chicken bone protein prepared by enzymatic method. Food Chem. 2019, 299, 125103. [Google Scholar] [CrossRef]
- Nourmohammadi, E.; SadeghiMahoonak, A.; Alami, M.; Ghorbani, M. Amino acid composition and antioxidative properties of hydrolysed pumpkin (Cucurbita pepo L.) oil cake protein. Int. J. Food Prop. 2017, 20, 3244–3255. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant and angiotensin I converting enzyme (ACE) inhibitory activities of date seed protein hydrolysates prepared using alcalase, flavourzyme and thermolysin. J. Funct. Foods 2015, 18, 1125–1137. [Google Scholar] [CrossRef]
- Arise, R.O.; Idi, J.J.; Mic-Braimoh, I.M.; Korode, E.; Ahmed, R.N.; Osemwegie, O. In vitro Angiotesin-1-converting enzyme, α-amylase and α-glucosidase inhibitory and antioxidant activities of Luffa cylindrical (L.) M. Roem seed protein hydrolysate. Heliyon 2019, 5, e01634. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, S.; Moslehishad, M.; Salami, M. Antioxidant and alpha-glucosidase enzyme inhibitory properties of hydrolyzed protein and bioactive peptides of quinoa. Int. J. Biol. Macromol. 2022, 213, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Fan, H.B.; Chalamaiah, M.; Wu, J.P. Preparation of low-molecular-w4eight, collagen hydrolysates (peptides): Current progress, challenges, and future perspectives. Food Chem. 2019, 301, 125222. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, Y.Y.; Gan, C.Y. Enzyme-assisted extraction and identification of antioxidative and α-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2016, 190, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Oseguera-Toledo, M.E.; de Mejia, E.G.; Amaya-Llano, S.L. Hard-to-cook bean (Phaseolus vulgaris L.) proteins hydrolyzed by alcalase and bromelain produced bioactive peptide fractions that inhibit targets of type-2 diabetes and oxidative stress. Food Res. Int. 2015, 76, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Mejía, E.; Batista, K.A.; Fernández, J.J.A.; Fernandes, K.F. Antihyperglycemic and hypoglycemic activity of naturally occurring peptides and protein hydrolysates from easy-to-cook and hard-to-cook beans (Phaseolus vulgaris L.). Food Res. Int. 2019, 121, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Crespo, I.S.; Molina, H.L.; Chel-Guerrero, L.; Ortiz-Andrade, R.; Betancur-Ancona, D. Inhibitory effect of peptide fractions derivatives from chia (salvia hispanica) hydrolysis against α-amylase and α-glucosidase enzymes. Nutr. Hosp. 2018, 35, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Uraipong, C.; Zhao, J. In vitro digestion of rice bran proteins produces peptides with potent inhibitory effects on α-glucosidase and angiotensin I converting enzyme. J. Sci. Food Agric. 2018, 98, 758–766. [Google Scholar] [CrossRef]
- Chaipoot, S.; Punfa, W.; Ounjaijean, S.; Phongphisutthinant, R.; Kulprachakarn, K.; Parklak, W.; Phaworn, L.; Rotphet, P.; Boonyapranai, K. Antioxidant, anti-diabetic, anti-obesity, and antihypertensive properties of protein hydrolysate and peptide fractions from black sesame cake. Molecules 2022, 28, 211. [Google Scholar] [CrossRef]
- Agrawal, H.; Joshi, R.; Gupta, M. Purification, identification and characterization of two novel antioxidant peptides from finger millet (Eleusine coracana) protein hydrolysate. Food Res. Int. 2019, 120, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wei, G.Q.; Li, K.L.; Duan, S.H.; Ye, R.; Huang, A.X. Identification and molecular docking of novel α-glucosidase inhibitory peptides from hydrolysates of Binglangjiang buffalo casein. LWT-Food Sci. Technol. 2022, 156, 113062. [Google Scholar] [CrossRef]
- Dakhili, S.; Abdolalizadeh, L.; Hosseini, S.M.; Shojaee-Aliabadi, S.; Mirmoghtadaie, L. Quinoa protein: Composition, structure and functional properties. Food Chem. 2019, 299, 125161. [Google Scholar] [CrossRef] [PubMed]
- Maestri, E.; Pavlicevic, M.; Montorsi, M.; Marmiroli, N. Meta-Analysis for correlating structure of bioactive peptides in foods of animal origin with regard to effect and stability. Compr. Rev. Food Sci. Food Saf. 2019, 18, 3–30. [Google Scholar] [CrossRef]
- Sadri, H.; Larki, N.N.; Kolahian, S. Hypoglycemic and hypolipidemic effects of leucine, zinc, and chromium, alone and in combination, in rats with type 2 diabetes. Biol. Trace Elem. Res. 2017, 180, 246–254. [Google Scholar] [CrossRef]
- Mollica, A.; Zengin, G.; Durdagi, S.; Salmas, R.E.; Macedonio, G.; Stefanucci, A.; Dimmito, M.P.; Novellino, E. Combinatorial peptide library screening for discovery of diverse α-glucosidase inhibitors using molecular dynamics simulations and binary QSAR models. J. Biomol. Struct. Dyn. 2019, 37, 726–740. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Mosoni, L.; Berlett, B.S.; Stadtman, E.R. Methionine residues as endogenous antioxidants in proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 15036–15040. [Google Scholar] [CrossRef]
- Wu, G.Y.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.L.; McKnight, J.R.; Satterfield, M.C.; et al. Proline and hydroxyproline metabolism: Implications for animal and human nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef]
- Huebner, J.; Muecke, R. Arginine-supplemented enteral nutrition. J. Cancer Res. Clin. 2014, 140, 681–683. [Google Scholar] [CrossRef] [PubMed]
- Chartrand, D.; Da Silva, M.S.; Julien, P.; Rudkowska, I. Influence of amino acids in dairy products on glucose homeostasis: The clinical evidence. Can. J. Diabetes 2017, 41, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.; Gaspar, A.R.M. Rational in silico design of novel α-glucosidase inhibitory peptides and in vitro evaluation of promising candidates. Biomed. Pharmacother. 2018, 107, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Kaur, A. Antidiabetic potential of a peptide isolated from an endophytic Aspergillus awamori. J. Appl. Microbiol. 2016, 120, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Pino, F.; Guadix, A.; Guadix, E.M. Identification of novel dipeptidyl peptidase IV and α-glucosidase inhibitory peptides from Tenebrio molitor. Food Funct. 2021, 12, 873–880. [Google Scholar] [CrossRef]
- Karimi, A.; Azizi, M.H.; Gavlighi, H.A. Fractionation of hydrolysate from corn germ protein by ultrafiltration: In vitro antidiabetic and antioxidant activity. Food Sci. Nutr. 2020, 8, 2395–2405. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, Y.N.; Kim, J.K.; Park, H.Y.; Song, B.S. Viscothionin purified from mistletoe (Viscum album var. coloratum Ohwi) induces insulin secretion from pancreatic beta cells. J. Ethnopharmacol. 2019, 234, 172–179. [Google Scholar] [CrossRef]
- Sun, W.J.; Wu, C.E.; Fan, G.J.; Hao, G.; Shi, H.J.; Zhang, C. Preparation of a functional beverage with α-glucosidase inhibitory peptides obtained from ginkgo seeds. J. Food Sci. Technol. 2021, 58, 4495–4503. [Google Scholar] [CrossRef]
- Tyagi, P.; Pechenov, S.; Subramony, J.A. Oral peptide delivery: Translational challenges due to physiological effects. J. Control Release 2018, 287, 167–176. [Google Scholar] [CrossRef]
- Megías, C.; del Mar Yust, M.; Pedroche, J.; Lquari, H.; Girón-Calle, J.; Alaiz, M.; Millán, F.; Vioque, J. Purification of an ACE inhibitory peptide after hydrolysis of sunflower (Helianthus annuus L.) protein isolates. J. Agric. Food Chem. 2004, 52, 1928–1932. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ding, X. Characterization of inhibition and stability of soy-protein-derived angiotensin I-converting enzyme inhibitory peptides. Food Res. Int. 2002, 35, 367–375. [Google Scholar] [CrossRef]
- You, L.J.; Zhao, M.; Regenstein, J.M.; Ren, J.Y. Changes in the antioxidant activity of loach (Misgurnus anguillicaudatus) protein hydrolysates during a simulated gastrointestinal digestion. Food Chem. 2010, 120, 810–816. [Google Scholar] [CrossRef]
- Ahmed, T.; Sun, X.H.; Udenigwe, C.C. Role of structural properties of bioactive peptides in their stability during simulated gastrointestinal digestion: A systematic review. Trends Food Sci. Technol. 2022, 120, 265–273. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
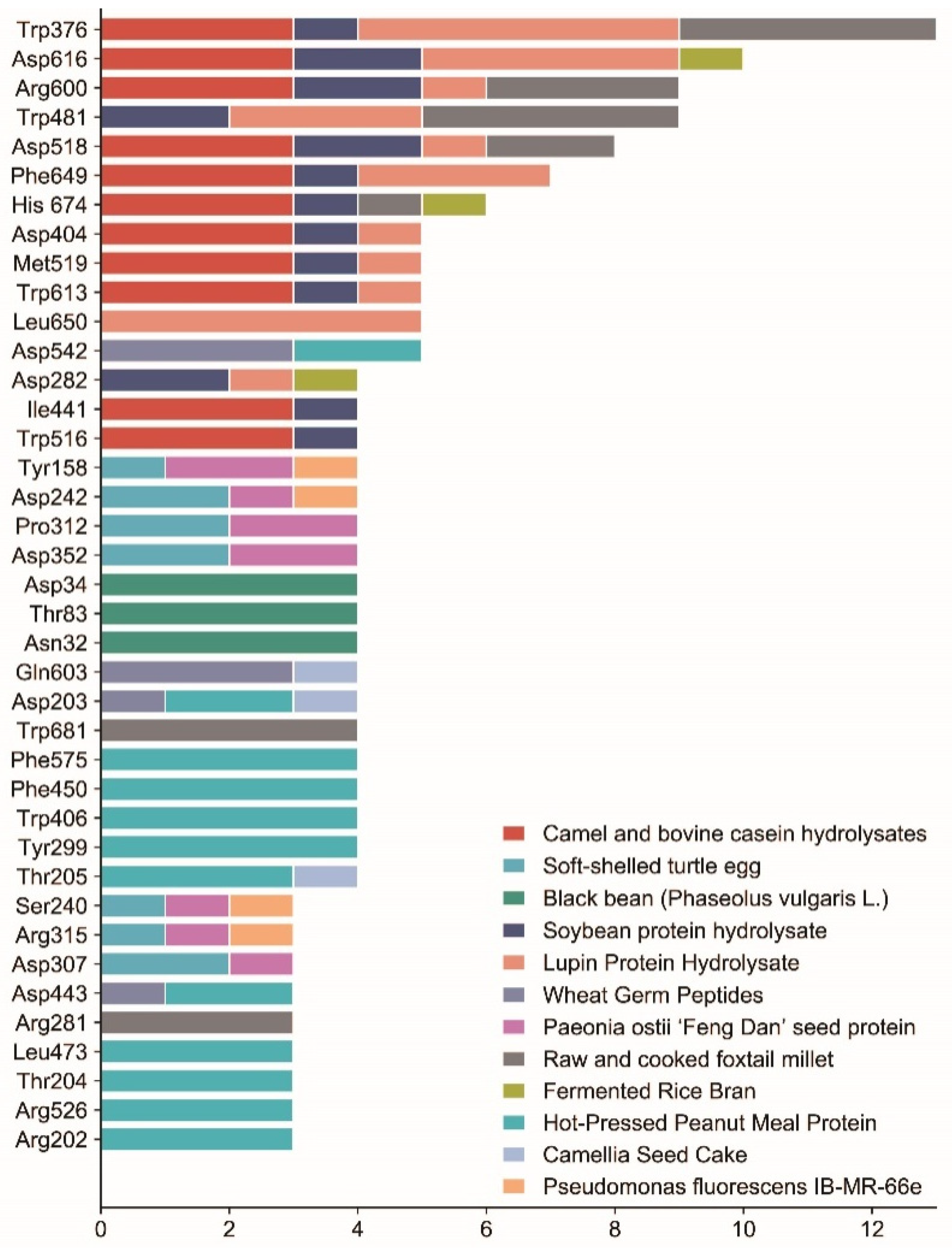
| Source | Amino Acid Sequence | Interaction Mechanism | Inhibition (IC50) | Bound Residues of α-Glucosidase | References | |
|---|---|---|---|---|---|---|
| Model | Interaction | |||||
| Camel and bovine casein hydrolysates | LPTGWLM, MFE, GPAHCLL | Hydrogen bond | Trp376, Asp404, Ile441, Trp516, Asp518, Met519, Arg600, Trp613, Asp616, Phe649, His674; | [37] | ||
| Camel whey proteins | CCGM, MFE | [38] | ||||
| Egg white protein | RVPSLM | 23.07 μmol/L | [39] | |||
| TPSPR | 40.02 μmol/L | |||||
| Albumin | KLPGF | 59.5 ± 5.7 μmol/L | [40] | |||
| NVLQPS | 100.0 ± 5.7 μmol/L | |||||
| Egg yolk protein hydrolysate | VTGRFAGHPAAQ | 365.4 μg/mL | [27] | |||
| LAPSLPGKPKPD | 1065.6 mM | [41] | ||||
| Parmigiano-Reggiano cheese | PFP | 8.6 mmol/L | [42] | |||
| Fermented rubing cheese | QPHQPLPP | Hydrogen bonds | 889 μmol/L | Arg428, Trp710, Asp568, Glu771, Asn448; | [43] | |
| TPVVVPPF | 4330 μmol/L | Glu361, Glu443, Arg428; | ||||
| Silk cocoon hydrolysate | GEY | 2.7 mg/mL | [44] | |||
| GYG | 1.5 mg/mL | |||||
| Silkworm pupae | QPGR | Hydrogen bonds | 65.8 μmol/L | [45] | ||
| SQSPA | 20 μmol/L | |||||
| QPPT | 560 μmol/L | |||||
| NSPR | 205 μmol/L | |||||
| Sericin peptides | SEDSSEVDIDLGNLG | Noncompetitive | Hydrogen bonds; Van der Waals; Hydrophobic interaction | 2.9 ± 0.1 μmol/L | [46] | |
| Chinese giant salamander (Andrias davidianus) protein hydrolysate | CSSV | 206 μg/mL | [47] | |||
| YSFR | 162 μg/mL | |||||
| SAAP | 66.9 μg/mL | |||||
| PGGP | 63.5 μg/mL | |||||
| LGGGN | 42.93 μg/mL | |||||
| Chinese giant salamander skin | GPPGPA | Competitive | Hydrogen bonds; Hydrophobic interaction | Hydrogen bonds: Asn53, Gln59, Trp80, Ala36, Ser59, Asp103, Gln102, Arg110, Arg188; Hydrophobic interaction: Ser56, Ala58, Asn199, Ser216, Leu78, Gln79, Phe225, Gln218, Ser18, Gln62, His66, Gln37, Tyr109, Leu393, Glu384, Lys106, Trp390, Ala211, Asp189, Tyr240, Ile185; | [48] | |
| Edible insects | FDPFPK | 5.95 μg/mL | [49] | |||
| Sardine muscle hydrolyzate | VW | 22.6 mM | [50] | |||
| WYPL | 3.7 mM | |||||
| Corbicula fluminea and Chlorella sorokiniana | VKP, VKK, VW, WV, IW, LW | [51] | ||||
| Soy protein | LLPLPVLK | 237.43 ± 0.52 μmol/L | [52] | |||
| SWLRL | 182.05 ± 0.74 μmol/L | |||||
| WLRL | 165.29 ± 0.74 μmol/L | |||||
| Ginkgo biloba seed protein | LSMSFPPF | Hydrogen bonds; Van der Waals; Hydrophobic interaction | 454.33 ± 32.45 μmol/L | Hydrogen bonds: Ala518, Pro512, His515, Phe516, Phe534, Ser92, Gly94; Hydrophobic interaction: Ala518, Ala533, Met535, Phe516, Phe534, Ala532, Val513, Ala451, Pro442; Van der Waals: Ala454, Phe455, Ala514, Asp440, Thr517, Thr519, Leu525, Asn447, Arg520, Leu93, His91, Lys89; | [53] | |
| VPKIPPP | 943.82 ± 73.1 μmol/L | Hydrogen bonds: Pro230, Asn301, Asp333; Hydrophobic interaction: Ala343, Val335, Arg340, Met302, Ala232, Pro230, Phe397, Val334, Phe297; Van der Waals: Glu377, Thr339, Leu227, Leu300, Pro602, Gly228, Arg400, Tyr389, Glu396, Asn233, Ala229, Glu231; | ||||
| MPGPPSD | 1446.81 ± 66.98 μmol/L | Hydrogen bonds: Gly94, Asn4, Asp48, Arg457, Pro442, Asp440; Hydrophobic interaction: Phe463, Arg457, Lys96, Trp7; Van der Waals: Asp441, Asn447, Met6, Asp464, Lys483, Leu95, Val47, Asn46, Ala43, Arg456, His459, Pro460, Gly11; | ||||
| Soybean protein hydrolysate | GSR | Noncompetitive | Hydrogen bonds; Van der Waals; Anion-π interactions | 20.4 μmol/L | Hydrogen bonds: Asp518, Asp616, Asp282; Van der Waals: Tyr292, Trp481, Asp518, Met519, Arg600, Trp613, Asp616; | [54] |
| EAK | 520.2 μmol/L | Hydrogen bonds: His674, Asp518, Arg600, Asp616, Asp282; Van der Waals: Trp376, Asp404, Ile441, Trp481, Trp516, Asp518, Phe649, His674; Anion-π interactions: Trp376, Trp481, Trp516, Phe649; | ||||
| Soft and hard wheat glutens | WGLYH | Hydrogen bonds; Electrostatic interaction | [55] | |||
| Paeonia ostia ‘Feng Dan’ seed protein | YFFM | Mixed-type | Hydrogen bonds; π-π stacking interactions | Hydrogen bonds: Tyr158, Asp352, Glu411, Arg315, Pro312; π-π stacking interactions: His280, Phe303; | [28] | |
| FFFM | Hydrogen bonds | 245.46 ± 44.01 μmol/L | Hydrogen bonds: Asp307, Gly309, Thr310, Pro312, Leu313; | |||
| YYFM | Hydrogen bonds | 306.71 ± 48.17 μmol/L | Hydrogen bonds: Tyr158, Ser240, Asp242, Asp352; | |||
| Dark tea protein | TAELLPR | 0.43 ± 0.03 mg/mL | [56] | |||
| CGKKFVR | 0.52 ± 0.09 mg/mL | |||||
| AVPANLVDLNVPALLK | 1.03 ± 0.13 mg/mL | |||||
| VVDLVFFAAAK | 0.04 ± 0.04 mg/mL | Pro395, Leu393, Val269, Glu271, Arg257, Phe297, Met281, Leu278, Asp322; | ||||
| Quinoa (Chenopodium quinoa Willd.) | IQEGGLT | Hydrophobic interaction | [57] | |||
| Sprouted quinoa yoghurt beverages | LAHMIVAGA | Noncompetitive | Hydrogen bonds | 10.9 mg/mL | [58] | |
| VAHPVF | 9.0 mg/mL | |||||
| Raw and cooked foxtail millet | AMFLPGA, TMMMLLP, FFLPQ, FMLPQ | Hydrogen bonds; Hydrophobic interaction | Hydrogen bonds: Arg600, Arg281, Ala284, Asp518, Ser676, Gly651, Trp481, His674; Hydrophobic interaction: Trp376, Trp481, Trp681; | [59] | ||
| Fermented rice bran | GLLGY | Noncompetitive | Hydrogen bonds | Asp282, Ser523, Asp616, His674; | [25] | |
| Hemp (Cannabis sativa L.) seed protein | LR, PLMLP | Hydrophobic interaction | [60] | |||
| The roots of Gypsophila oldhamiana | Cyclo-(GFDFILP) | 305 μmol/L | [26] | |||
| Wheat germ peptides | LDLQR | Hydrogen bonds | 8.59 mM | Asp443, Arg334, Gln603, Asp203, Asp542; | [61] | |
| AGGFR | 8.66 mM | Asp542, Gln603 | ||||
| LDNFR | 9.21 mM | |||||
| Spirulina platensis | GVPMPNK | 151.5 μg/mL | [62] | |||
| RNPFVFAPTLLTVAAR | 164.5 μg/mL | |||||
| LRSELAAWSR | 134.2 μg/mL | Thr168, Leu144, Ile146, Gly149, Arg10, Trp8; | ||||
| Walnut (Juglans mandshurica maxim.) | LPLLR | [63] | ||||
| Hot-pressed peanut meal protein | FYNPAAGR, PGVLPVAS, FFVPPSQQ, FSYNPQAG | Hydrogen bonds; Hydrophobic interaction; Salt bridges; π-stacking | Hydrogen bonds: Arg202, Asp203, Thr205, Asn209, Asp327, Asp443, Asp474, Arg526, Asp542; Hydrophobic interaction: Thr204, Tyr299, Trp406, Phe450, Leu473, Lys480, Phe575; Salt bridges: Asp327, Asp443, Asp542, Arg202; | [64] | ||
| Black bean (Phaseolus vulgaris L.) | TTGGKGGK | Hydrogen bonds; Hydrophobic interaction; Polar interactions | [65] | |||
| AKSPLF | Hydrogen bonds: Asp34, Thr83, Asp89; Polar interactions: Asp34, Thr83, Asp89, Asn32; | |||||
| QTPF | Hydrogen bonds: Asn32, Asp34, Thr83; Polar interactions: Asn32, Asp34, Trp36, Thr83;Hydrophobic interaction: Pro82; | |||||
| FEELN | Hydrogen bonds: Asp34, Thr83; Polar interactions: Asn32, Asp34, Thr83; | |||||
| LSKSVL | Hydrogen bonds: Asp34, Thr83, Asp89, Asn32; Polar interactions: Asp34, Thr83, Asp89, Asn32; | |||||
| Chickpea (Cicerarietinum L.) protein hydrolyzates | FGKG | Competitive | Hydrophobic interaction | Trp59, Leu162, Ala198, His305; | [66] | |
| Camellia seed cake | LLVLYYEY | Noncompetitive | Hydrogen bonds | 0.33 mM | Arg730, Gly732, Arg653, Glu661; | [67] |
| LLLLPSYSEF | Mixed-type | 1.11 mM | Asp203, Thr205, Tyr605, Gln603; | |||
| Aspergillus oryzae | PFP | Mixed-type | 3.1 mg/mL | [68] | ||
| Pseudomonas fluorescens IB-MR-66e | Cyclo(L-Pro-L-Leu) | Hydrophobic interaction; Hydrophilic interaction; Hydrogen bonds | Hydrogen bonds: Ser157; Hydrophobic interaction: Lys156, Tyr158, Asp242, Val232, Asp233, Phe314, Arg315, Tyr316, Asn415; Hydrophilic interaction: Ser157, Ser240, Ser241; | [69] | ||
| Soft-shelled turtle egg | ARDASVLK | Hydrogen bonds; Hydrophobic interaction | 195 μmol/L | Tyr158, Ser240, Asp242, Glu277, Thr310, Pro312, Arg315, Asp352, Glu411; | [70,71] | |
| HNKPEVEVR | 56 μmol/L | Lys156, Ser157, Asp242, Asp307, Pro312; | ||||
| SGTLLHK | 289 μmol/L | Asp215, Glu277, Asp307, Asp352; | ||||
| Moringa oleifera seed protein | KETTTIVR | Hydrogen bonds; Hydrophobic interaction | 109.65 μmol/L | Hydrogen bonds: Arg429, Asp379, Val380, Asp333, Leu300, Leu227, Lys398; Hydrophobic interaction: Ala378, Gly399, Arg400, Phe397, Val334, Tyr389, Asn301, Ala229, Met302, Val335, Glu231, Pro230, Pro395, Glu377; | [72] | |
| Lupin protein hydrolysate | SPRRF, FE, RR, RPR, LRP, PPGIP | Hydrogen bonds; Hydrophobic interaction | Hydrogen bonds: Asp616, Asp518, Arg411; Hydrophobic interaction: Trp376, Phe649, Asp404, Asp616, Ser676, Leu650, Leu678, Trp481, Asp282, Phe525, Phe649, Trp613, Leu677, Ser379, Asn417, Leu405, Asp419, Met408, Ser410, Arg600, Met519; | [73] | ||
| Almond (Armeniaca sibirica) oil manufacture residue | WH | 16.99 ± 0.05 μmol/Lol/L | [74] | |||
| WS | 44.63 ± 0.03 μmol/Lol/L | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, H.; Xie, T.; Wu, Q.; Hu, Z.; Luo, Y.; Luo, F. Alpha-Glucosidase Inhibitory Peptides: Sources, Preparations, Identifications, and Action Mechanisms. Nutrients 2023, 15, 4267. https://doi.org/10.3390/nu15194267
Lu H, Xie T, Wu Q, Hu Z, Luo Y, Luo F. Alpha-Glucosidase Inhibitory Peptides: Sources, Preparations, Identifications, and Action Mechanisms. Nutrients. 2023; 15(19):4267. https://doi.org/10.3390/nu15194267
Chicago/Turabian StyleLu, Han, Tiantian Xie, Qi Wu, Zuomin Hu, Yi Luo, and Feijun Luo. 2023. "Alpha-Glucosidase Inhibitory Peptides: Sources, Preparations, Identifications, and Action Mechanisms" Nutrients 15, no. 19: 4267. https://doi.org/10.3390/nu15194267
APA StyleLu, H., Xie, T., Wu, Q., Hu, Z., Luo, Y., & Luo, F. (2023). Alpha-Glucosidase Inhibitory Peptides: Sources, Preparations, Identifications, and Action Mechanisms. Nutrients, 15(19), 4267. https://doi.org/10.3390/nu15194267