
Enhancing Bioaccessibility of Plant Protein Using Probiotics: An In Vitro Study

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Probiotic Bacteria
2.2. Protein Ingredients
2.3. Protein Digestion in Simulated Human Upper GI Tract
2.4. Analysis of Soluble Protein
2.5. Analysis of Free Amino Nitrogen and Degree of Hydrolysis
2.6. Analysis of Free Amino Acids
2.7. Analysis of Biogenic Amines
2.8. Microbiological Analyses
2.9. Statistical Analysis
3. Results
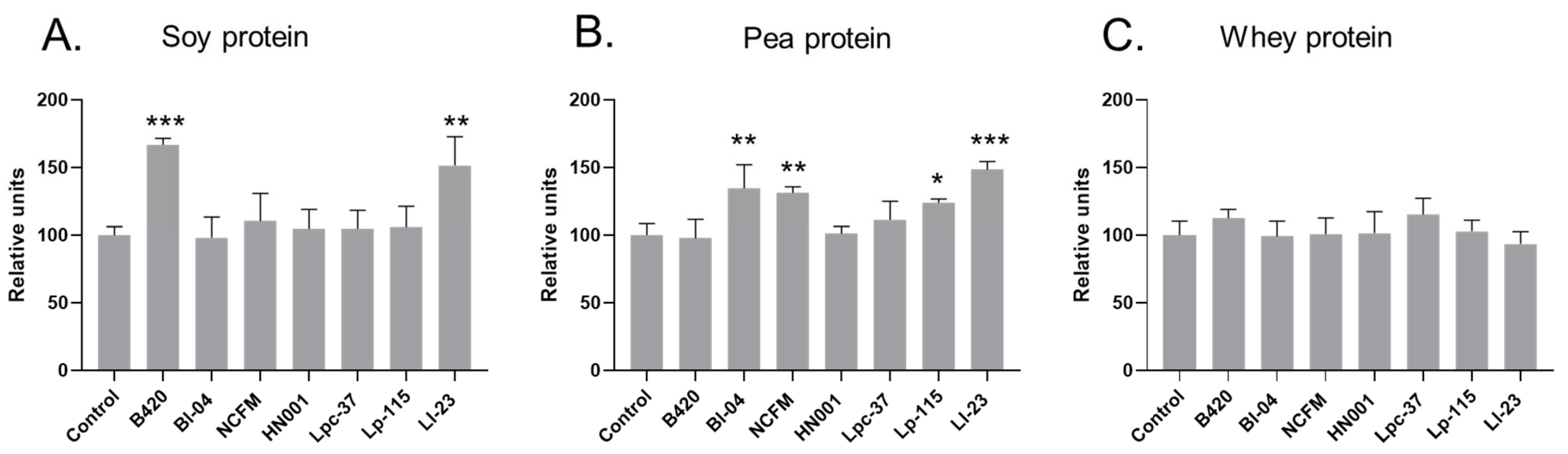
3.1. Effect of Probiotics on Protein Solubility
3.2. Effect of Probiotics on Protein Hydrolysis and Release of Free Amino Nitrogen
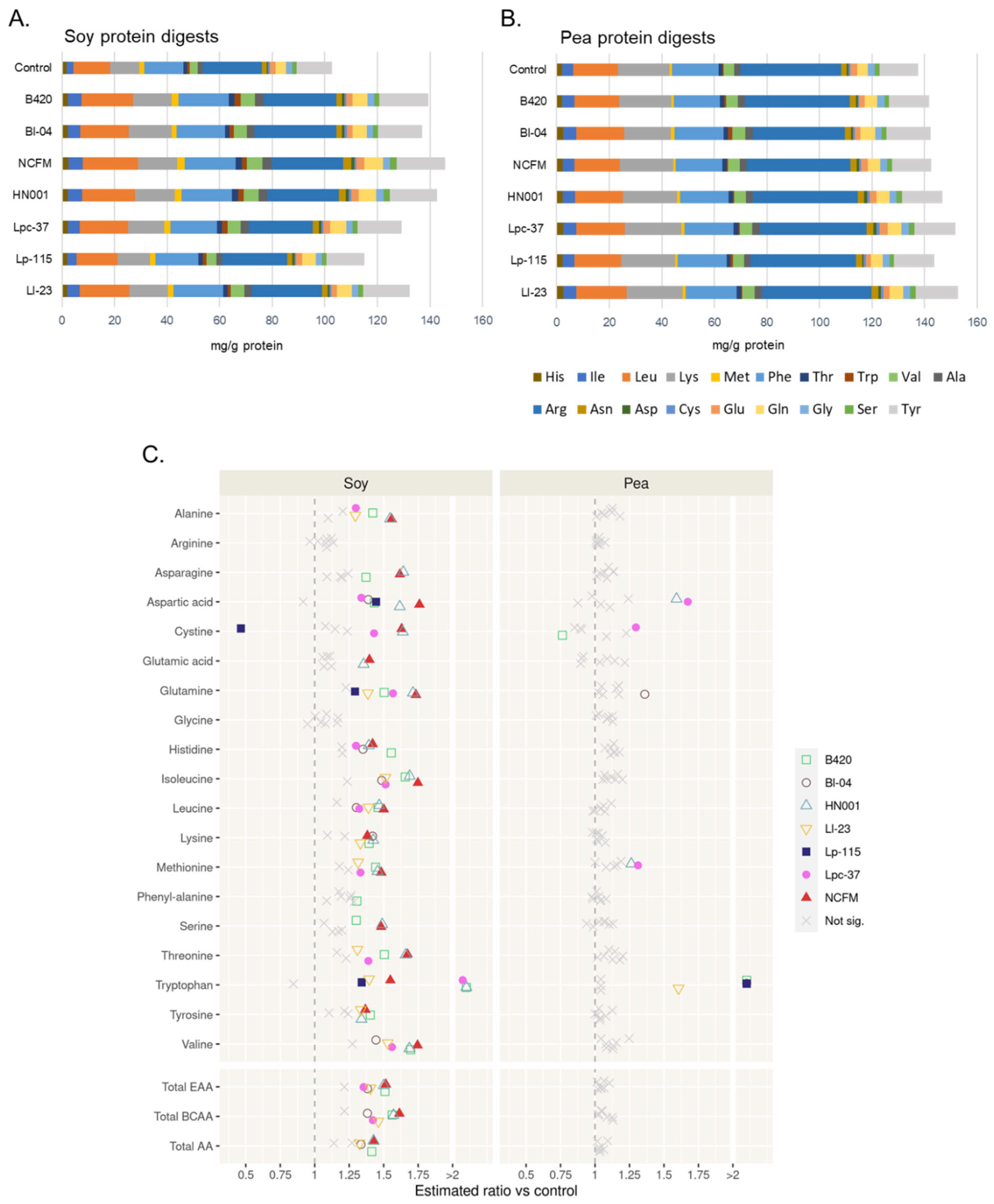
3.3. Effect of Probiotics on Release of Free Amino Acids from Soy and Pea Protein
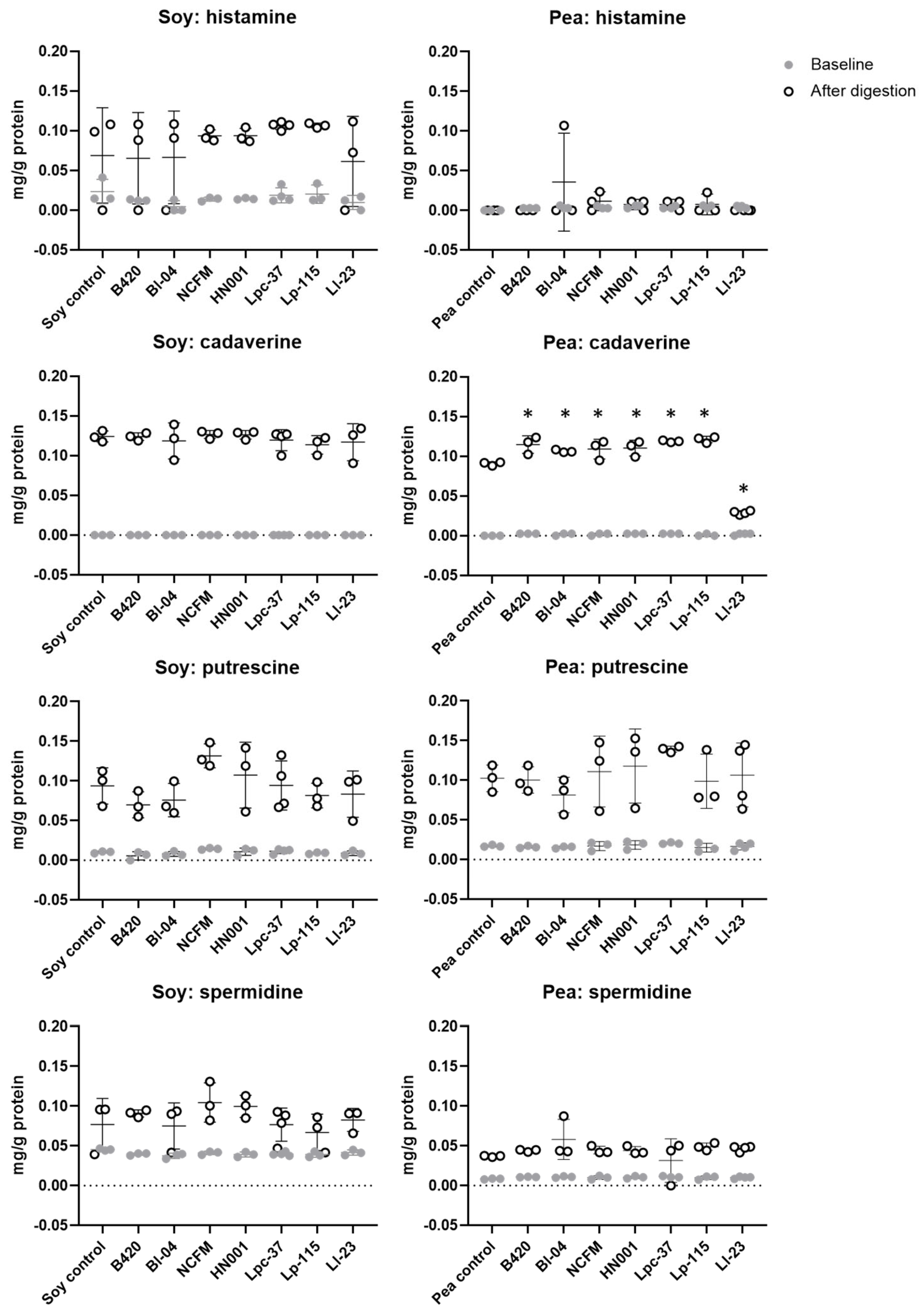
3.4. Effect of Probiotics on Production of Biogenic Amines during Digestion of Soy and Pea Protein
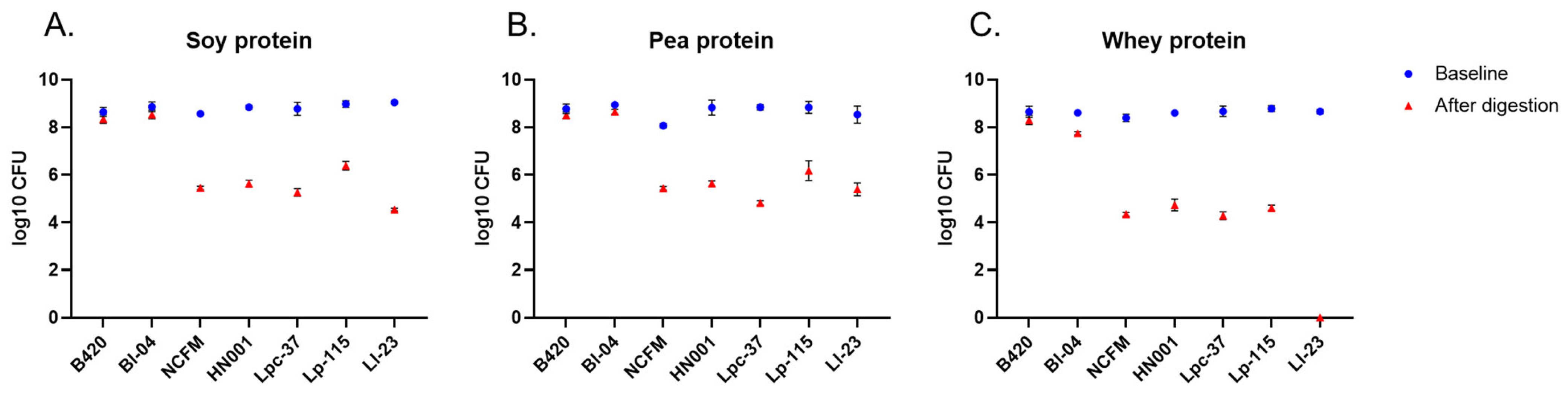
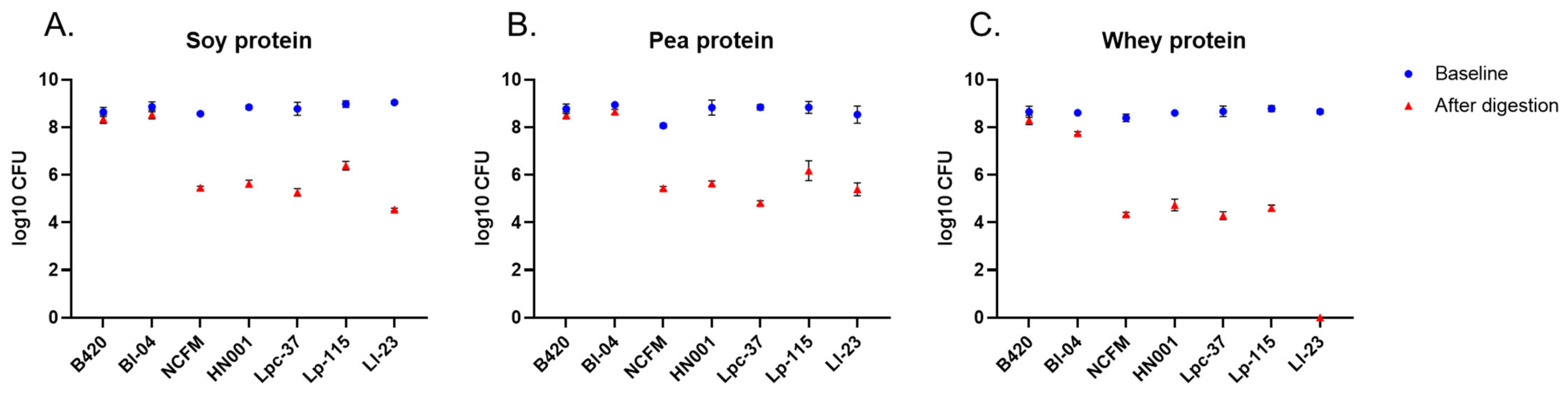
3.5. Probiotic Survival after the Simulated Digestion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alcorta, A.; Porta, A.; Tárrega, A.; Alvarez, M.D.; Vaquero, M.P. Foods for plant-based diets: Challenges and innovations. Foods 2021, 10, 293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, Y.; Li, H.; Liu, X. The potential of proteins, hydrolysates and peptides as growth factors for Lactobacillus and Bifidobacterium: Current research and future perspectives. Food Funct. 2020, 11, 1946–1957. [Google Scholar] [CrossRef]
- Aguirre, L.; Hebert, E.M.; Garro, M.S.; de Giori, G.S. Proteolytic activity of Lactobacillus strains on soybean proteins. LWT-Food Sci. Technol. 2014, 59, 780–785. [Google Scholar] [CrossRef]
- Çabuk, B.; Nosworthy, M.G.; Stone, A.K.; Korber, D.R.; Tanaka, T.; House, J.D.; Nickerson, M.T. Effect of fermentation on the protein digestibility and levels of non-nutritive compounds of pea protein concentrate. Food Technol. Biotechnol. 2018, 56, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Sibakov, N.; Re, M.; Karsma, A.; Laitila, A.; Nordlund, E. Phytic acid reduction by bioprocessing as a tool to improve the in vitro digestibility of faba bean protein. J. Agric. Food Chem. 2018, 66, 10394–10399. [Google Scholar] [CrossRef]
- Manus, J.; Millette, M.; Dridi, C.; Salmieri, S.; Aguilar Uscanga, B.R.; Lacroix, M. Protein quality of a probiotic beverage enriched with pea and rice protein. J. Food Sci. 2021, 86, 3698–3706. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Jäger, R.; Zaragoza, J.; Purpura, M.; Iametti, S.; Marengo, M.; Tinsley, G.M.; Anzalone, A.J.; Oliver, J.M.; Fiore, W.; Biffi, A. Probiotic administration increases amino acid absorption from plant protein: A placebo-controlled, randomized, double-blind, multicenter, crossover study. Probiotics Antimicrob. Proteins 2020, 12, 1330–1339. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H. Influence of probiotics on dietary protein digestion and utilization in the gastrointestinal tract. Curr. Protein Pept. Sci. 2019, 20, 125–131. [Google Scholar] [CrossRef]
- van der Wielen, N.; Moughan, P.J.; Mensink, M. Amino acid absorption in the large intestine of humans and porcine models. J. Nutr. 2017, 147, 1493–1498. [Google Scholar] [CrossRef]
- Lin, R.; Liu, W.; Piao, M.; Zhu, H. A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids 2017, 49, 2083–2090. [Google Scholar] [CrossRef] [PubMed]
- Keller, D.; Van Dinter, R.; Cash, H.; Farmer, S.; Venema, K. Bacillus coagulans GBI-30, 6086 increases plant protein digestion in a dynamic, computer-controlled in vitro model of the small intestine (TIM-1). Benef. Microbes 2017, 8, 491–496. [Google Scholar] [CrossRef]
- Jäger, R.; Purpura, M.; Farmer, S.; Cash, H.A.; Keller, D. Probiotic Bacillus coagulans GBI-30, 6086 improves protein absorption and utilization. Probiotics Antimicrob. Proteins 2018, 10, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Stecker, R.A.; Moon, J.M.; Russo, T.J.; Ratliff, K.M.; Mumford, P.W.; Jäger, R.; Purpura, M.; Kerksick, C.M. Bacillus coagulans GBI-30, 6086 improves amino acid absorption from milk protein. Nutr. Metab. 2020, 17, 93. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wan, Y.; Yin, K.; Wei, Y.; Wang, B.; Yu, X.; Ni, Y.; Zheng, J.; Huang, T.; Song, M. Lower Circulating Branched-Chain Amino Acid Concentrations among Vegetarians are Associated with Changes in Gut Microbial Composition and Function. Mol. Nutr. Food Res. 2019, 63, 1900612. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.L.; Keerati-U-Rai, M.; Yin, H.; Daoust, J.; Nonnotte, E.; Quinquis, L.; St-Denis, T.; Bolster, D.R. Differential responses of blood essential amino acid levels following ingestion of high-quality plant-based protein blends compared to whey protein—A double-blind randomized, cross-over, clinical trial. Nutrients 2019, 11, 2987. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Thakur, N.; Raigond, P.; Singh, Y.; Mishra, T.; Singh, B.; Lal, M.K.; Dutt, S. Recent updates on bioaccessibility of phytonutrients. Trends Food Sci. Technol. 2020, 97, 366–380. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Salli, K.; Hirvonen, J.; Siitonen, J.; Ahonen, I.; Anglenius, H.; Maukonen, J. Selective Utilization of the Human Milk Oligosaccharides 2′-Fucosyllactose, 3-Fucosyllactose, and Difucosyllactose by Various Probiotic and Pathogenic Bacteria. J. Agric. Food Chem. 2021, 69, 170–182. [Google Scholar] [CrossRef]
- Greene, J.; Henderson, J.W., Jr.; Wikswo, J.P. Rapid and precise determination of cellular amino acid flux rates using HPLC with automated derivatization with absorbance detection (application note). Agilent Technologies. 2009, February 6, pp. 1–8. Available online: https://www.agilent.com/Library/applications/5990-3283EN.pdf (accessed on 18 May 2021).
- Saarinen, M. Determination of biogenic amines as dansyl derivatives in intestinal digesta and feces by reversed phase HPLC. Chromatographia 2002, 55, 297–300. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 18 May 2021).
- Venables, W.; Ripley, B. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; D’Agostino McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Egger, L.; Schlegel, P.; Baumann, C.; Stoffers, H.; Guggisberg, D.; Brugger, C.; Durr, D.; Stoll, P.; Vergeres, G.; Portmann, R. Physiological comparability of the harmonized INFOGEST in vitro digestion method to in vivo pig digestion. Food Res. Int. 2017, 102, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, Z.; Li, Y.; Meng, X.; Sui, X.; Qi, B.; Zhou, L. Relationship between surface hydrophobicity and structure of soy protein isolate subjected to different ionic strength. Int. J. Food Prop. 2015, 18, 1059–1074. [Google Scholar] [CrossRef]
- Lu, Z.; He, J.; Zhang, Y.; Bing, D. Composition, physicochemical properties of pea protein and its application in functional foods. Crit. Rev. Food Sci. Nutr. 2020, 60, 2593–2605. [Google Scholar] [CrossRef] [PubMed]
- Gilani, G.S.; Xiao, C.W.; Cockell, K.A. Impact of antinutritional factors in food proteins on the digestibility of protein and the bioavailability of amino acids and on protein quality. Br. J. Nutr. 2012, 108 (Suppl. S2), S315–S332. [Google Scholar] [CrossRef]
- Judkins, T.C.; Archer, D.L.; Kramer, D.C.; Solch, R.J. Probiotics, nutrition, and the small intestine. Curr. Gastroenterol. Rep. 2020, 22, 2. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- Liu, M.; Bayjanov, J.R.; Renckens, B.; Nauta, A.; Siezen, R.J. The proteolytic system of lactic acid bacteria revisited: A genomic comparison. BMC Genom. 2010, 11, 36. [Google Scholar] [CrossRef]
- Altermann, E.; Russell, W.M.; Azcarate-Peril, M.A.; Barrangou, R.; Buck, B.L.; McAuliffe, O.; Souther, N.; Dobson, A.; Duong, T.; Callanan, M. Complete genome sequence of the probiotic lactic acid bacterium Lactobacillus acidophilus NCFM. Proc. Natl. Acad. Sci. USA 2005, 102, 3906–3912. [Google Scholar] [CrossRef]
- Janer, C.; Arigoni, F.; Lee, B.; Peláez, C.; Requena, T. Enzymatic ability of Bifidobacterium animalis subsp. lactis to hydrolyze milk proteins: Identification and characterization of endopeptidase O. Appl. Environ. Microbiol. 2005, 71, 8460–8465. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, S.H.; Crombag, J.J.; Senden, J.M.; Waterval, W.H.; Bierau, J.; Verdijk, L.B.; van Loon, L.J. Protein content and amino acid composition of commercially available plant-based protein isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of biogenic amines on food quality and safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific opinion on risk based control of biogenic amine formation in fermented foods. Efsa J. 2011, 9, 2393. [Google Scholar] [CrossRef]
- Marteau, P.; Minekus, M.; Havenaar, R.; Huis, J. Survival of lactic acid bacteria in a dynamic model of the stomach and small intestine: Validation and the effects of bile. J. Dairy Sci. 1997, 80, 1031–1037. [Google Scholar] [CrossRef]
- Madureira, A.R.; Amorim, M.; Gomes, A.M.; Pintado, M.E.; Malcata, F.X. Protective effect of whey cheese matrix on probiotic strains exposed to simulated gastrointestinal conditions. Food Res. Int. 2011, 44, 465–470. [Google Scholar] [CrossRef]
- Faye, T.; Tamburello, A.; Vegarud, G.E.; Skeie, S. Survival of lactic acid bacteria from fermented milks in an in vitro digestion model exploiting sequential incubation in human gastric and duodenum juice. J. Dairy Sci. 2012, 95, 558–566. [Google Scholar] [CrossRef]
- Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Effect of different matrices on probiotic resistance to in vitro simulated gastrointestinal conditions. Int. J. Dairy Technol. 2015, 68, 595–601. [Google Scholar] [CrossRef]
- Santos-Hernández, M.; Alfieri, F.; Gallo, V.; Miralles, B.; Masi, P.; Romano, A.; Ferranti, P.; Recio, I. Compared digestibility of plant protein isolates by using the INFOGEST digestion protocol. Food Res. Int. 2020, 137, 109708. [Google Scholar] [CrossRef]
- Ariëns, R.M.; Bastiaan-Net, S.; Van de Berg-Somhorst, D.B.; El Bachrioui, K.; Boudewijn, A.; van den Dool, R.T.; de Jong, G.A.; Wichers, H.J.; Mes, J.J. Comparing nutritional and digestibility aspects of sustainable proteins using the INFOGEST digestion protocol. J. Funct. Foods 2021, 87, 104748. [Google Scholar] [CrossRef]
- Ren, C.; Xiong, W.; Peng, D.; He, Y.; Zhou, P.; Li, J.; Li, B. Effects of thermal sterilization on soy protein isolate/polyphenol complexes: Aspects of structure, in vitro digestibility and antioxidant activity. Food Res. Int. 2018, 112, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.S.; Hettiarachchy, N.S.; Kumar, T.K.S.; Carbonero, F.; Martin, E.M.; Benamara, M. Physicochemical properties of soy protein hydrolysate and its formulation and stability with encapsulated probiotic under in vitro gastrointestinal environment. J. Food Sci. 2020, 85, 3543–3551. [Google Scholar] [CrossRef] [PubMed]
- Egger, L.; Menard, O.; Baumann, C.; Duerr, D.; Schlegel, P.; Stoll, P.; Vergeres, G.; Dupont, D.; Portmann, R. Digestion of milk proteins: Comparing static and dynamic in vitro digestion systems with in vivo data. Food Res. Int. 2019, 118, 32–39. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Media |
|---|---|---|
| Bifidobacterium animalis subsp. lactis | B420 | Bifidobacterium medium 58 |
| Bifidobacterium animalis subsp. lactis | Bl-04 | Bifidobacterium medium 58 |
| Lactobacillus acidophilus | NCFM | MRS |
| Lacticaseibacillus rhamnosus | HN001 | MRS |
| Lacticaseibacillus paracasei subsp. paracasei | Lpc-37 | MRS |
| Lactiplantibacillus plantarum | Lp-115 | MRS |
| Lactococcus lactis subsp. lactis | Ll-23 | MRS |
| Treatment | FAN, Baseline (mg/g Protein) | FAN, after Digestion (mg/g Protein) | DH (%) |
|---|---|---|---|
| Soy protein | |||
| Control | 6.52 (0.30) | 23.4 (3.0) † | 25.4 (3.2) |
| B420 | 7.31 (0.53) | 27.8 (0.84) †* | 30.2 (0.9) |
| Bl-04 | 7.82 (0.17) | 31.1 (2.5) †*** | 33.8 (2.7) ** |
| NCFM | 8.34 (0.43) | 30.8 (1.4) †*** | 33.4 (1.5) * |
| HN001 | 7.31 (0.44) | 30.6 (2.7) †*** | 33.2 (2.9) * |
| Lpc-37 | 7.12 (0.31) | 28.1 (2.7) †** | 30.5 (2.9) |
| Lp-115 | 7.31 (0.18) | 28.7 (3.4) †** | 31.1 (3.7) |
| Ll-23 | 6.72 (0.16) | 25.4 (2.7) † | 27.5 (2.9) |
| Pea protein | |||
| Control | 3.23 (0.29) | 26.1 (2.75) † | 28.4 (3.0) |
| B420 | 4.21 (0.24) | 29.4 (2.02) †* | 32.0 (2.2) |
| Bl-04 | 4.10 (0.68) | 27.8 (1.68) † | 30.2 (1.8) |
| NCFM | 3.77 (0.66) | 30.2 (1.01) †** | 32.8 (1.1) |
| HN001 | 3.54 (1.16) | 26.2 (3.03) † | 28.5 (3.3) |
| Lpc-37 | 4.21 (0.66) | 24.9 (0.73) † | 27.0 (0.8) |
| Lp-115 | 3.13 (0.49) | 27.0 (1.43) † | 29.3 (1.6) |
| Ll-23 | 3.31 (0.57) | 25.7 (0.86) † | 28.0 (0.9) |
| Whey protein | |||
| Control | 9.6 (0.26) | 37.6 (5.43) † | 33.6 (4.8) |
| B420 | 12.6 (1.50) | 42.1 (1.64) † | 37.6 (1.5) |
| Bl-04 | 13.5 (0.73) | 41.2 (3.66) † | 36.8 (3.2) |
| NCFM | 12.1 (1.84) | 35.2 (2.75) † | 31.4 (2.5) |
| HN001 | 14.4 (3.84) | 35.0 (2.14) † | 31.3 (1.9) |
| Lpc-37 | 13.5 (1.84) | 45.1 (2.79) †* | 40.2 (2.5) |
| Lp-115 | 13.1 (1.78) | 38.6 (4.52) † | 34.5 (4.0) |
| Ll-23 | 12.1 (1.44) | 39.0 (4.25) † | 34.8 (3.8) |
| Soy Protein | Pea Protein | ||||||
|---|---|---|---|---|---|---|---|
| Probiotic | Survival a | Protein Solubility b | Protein Hydrolysis (FAN) b | Free Amino Acids b | Protein Solubility b | Protein Hydrolysis (FAN) b | Free Amino Acids b |
| B. animalis subsp. lactis B420 | ↔ | ↑↑ | ↑ | ↑↑ | ↔ | ↑ | |
| B. animalis subsp. lactis Bl-04 | ↔ | ↔ | ↑↑ | ↑ | ↑↑ | ↔ | Gln↑ |
| L. acidophilus NCFM | ↓ | ↔ | ↑↑ | ↑↑ | ↑↑ | ↑↑ | |
| L. rhamnosus HN001 | ↓ | ↔ | ↑↑ | ↑↑ | ↔ | ↔ | Asp↑↑, Met↑ |
| L. paracasei subsp. paracasei Lpc-37 | ↓ | ↔ | ↑↑ | ↑ c | ↔ | ↔ | Asp↑↑, Cys↑, Met↑↑ |
| L. plantarum Lp-115 | ↓ | ↔ | ↑↑ | ↔ | ↑ | ↔ | |
| L. lactis subsp. lactis Ll-23 | ↓ | ↑↑ | ↔ | ↑ | ↑↑ | ↔ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marttinen, M.; Anjum, M.; Saarinen, M.T.; Ahonen, I.; Lehtinen, M.J.; Nurminen, P.; Laitila, A. Enhancing Bioaccessibility of Plant Protein Using Probiotics: An In Vitro Study. Nutrients 2023, 15, 3905. https://doi.org/10.3390/nu15183905
Marttinen M, Anjum M, Saarinen MT, Ahonen I, Lehtinen MJ, Nurminen P, Laitila A. Enhancing Bioaccessibility of Plant Protein Using Probiotics: An In Vitro Study. Nutrients. 2023; 15(18):3905. https://doi.org/10.3390/nu15183905
Chicago/Turabian StyleMarttinen, Maija, Mehreen Anjum, Markku T. Saarinen, Ilmari Ahonen, Markus J. Lehtinen, Päivi Nurminen, and Arja Laitila. 2023. "Enhancing Bioaccessibility of Plant Protein Using Probiotics: An In Vitro Study" Nutrients 15, no. 18: 3905. https://doi.org/10.3390/nu15183905
APA StyleMarttinen, M., Anjum, M., Saarinen, M. T., Ahonen, I., Lehtinen, M. J., Nurminen, P., & Laitila, A. (2023). Enhancing Bioaccessibility of Plant Protein Using Probiotics: An In Vitro Study. Nutrients, 15(18), 3905. https://doi.org/10.3390/nu15183905