
Probiotic OMNi-BiOTiC® 10 AAD Reduces Cyclophosphamide-Induced Inflammation and Adipose Tissue Wasting in Mice


 , ,
, ,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Inflammatory Response
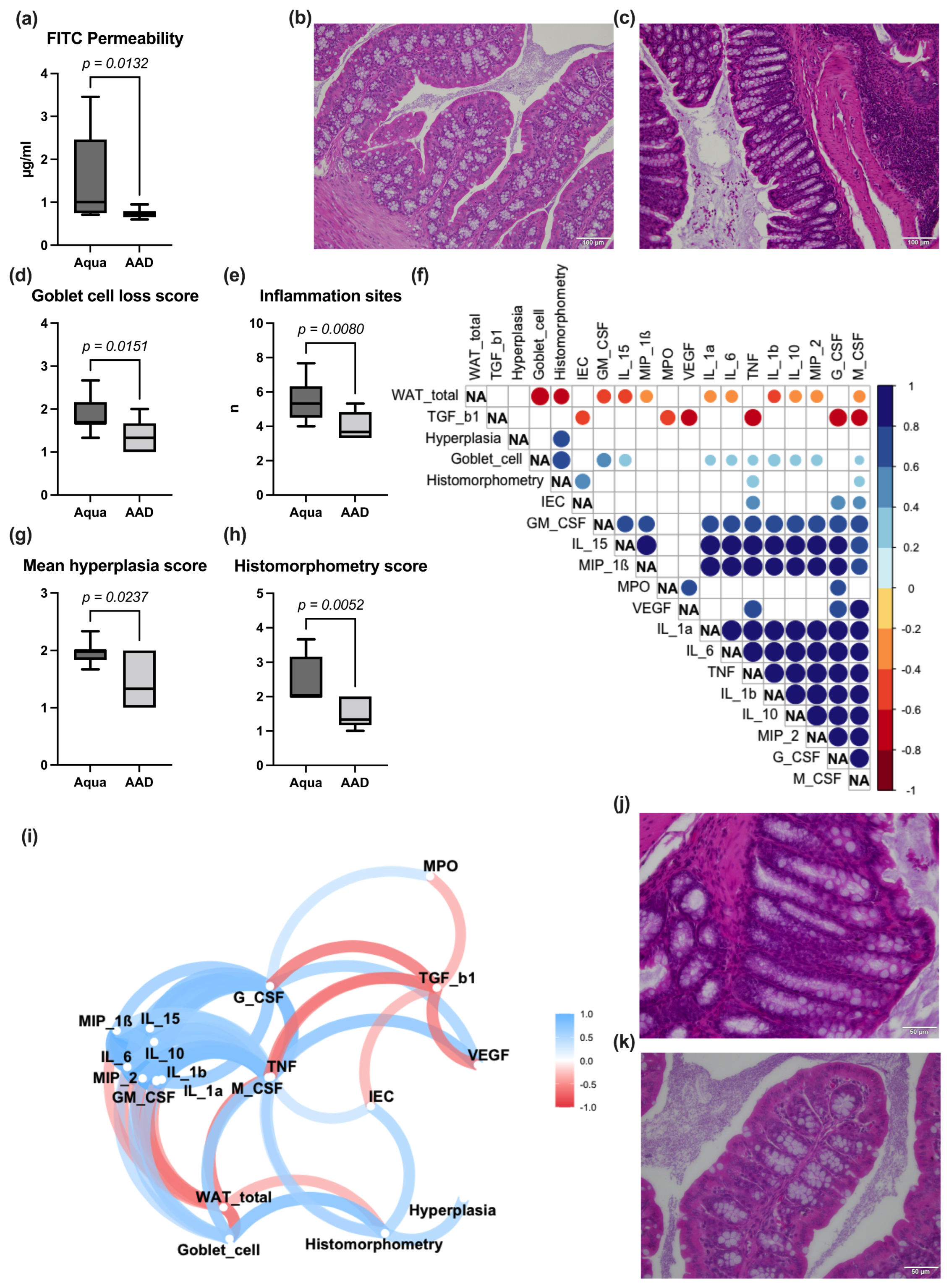
2.2. Gut Permeability Assay and Bowel Wall Integrity
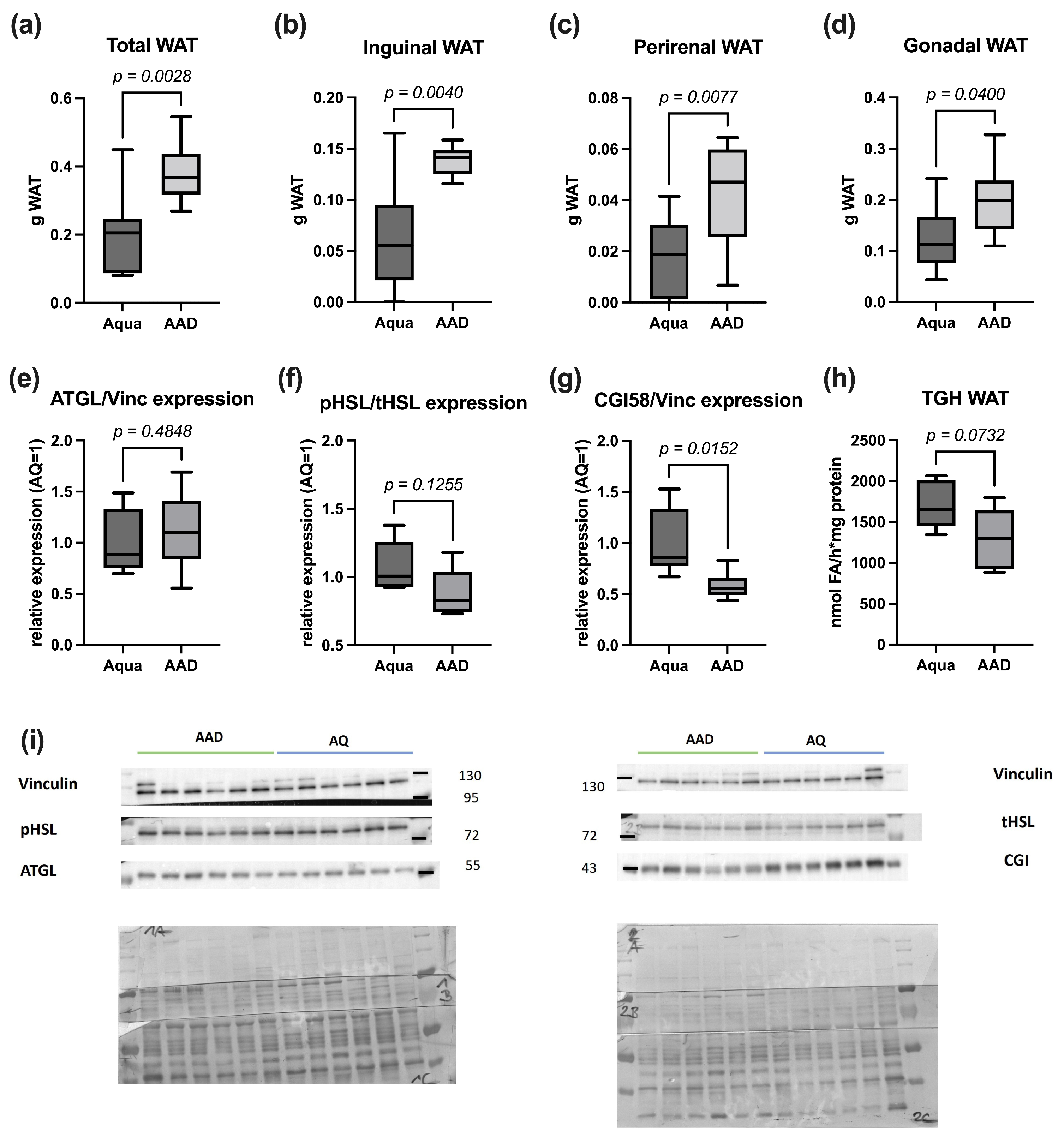
2.3. Lipolysis Markers
2.4. Bowel Wall Inflammation
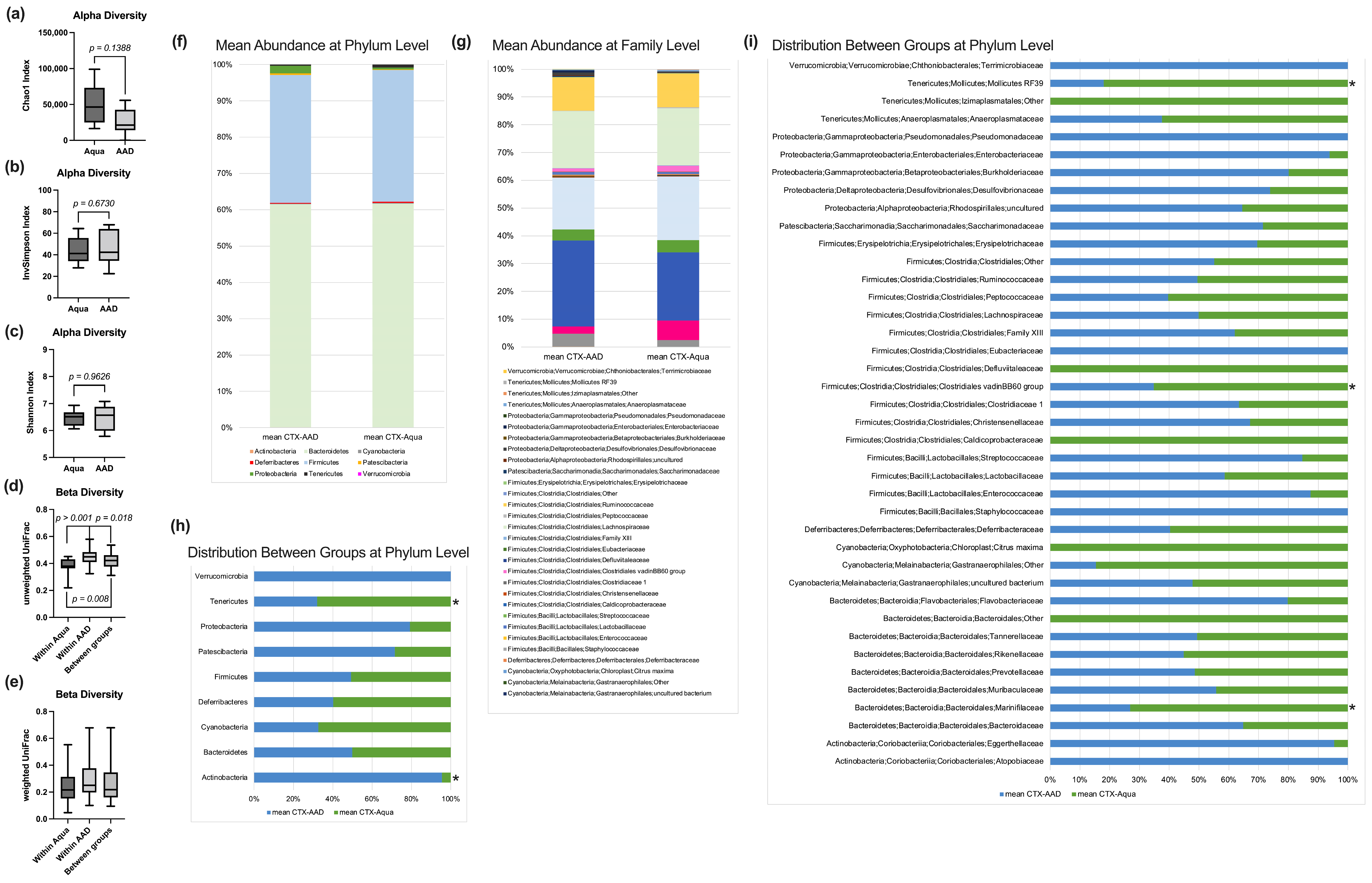
2.5. Fecal Microbiome Analysis
2.6. Fecal Volatile Organic Compounds
2.7. Statistical Analysis
3. Results
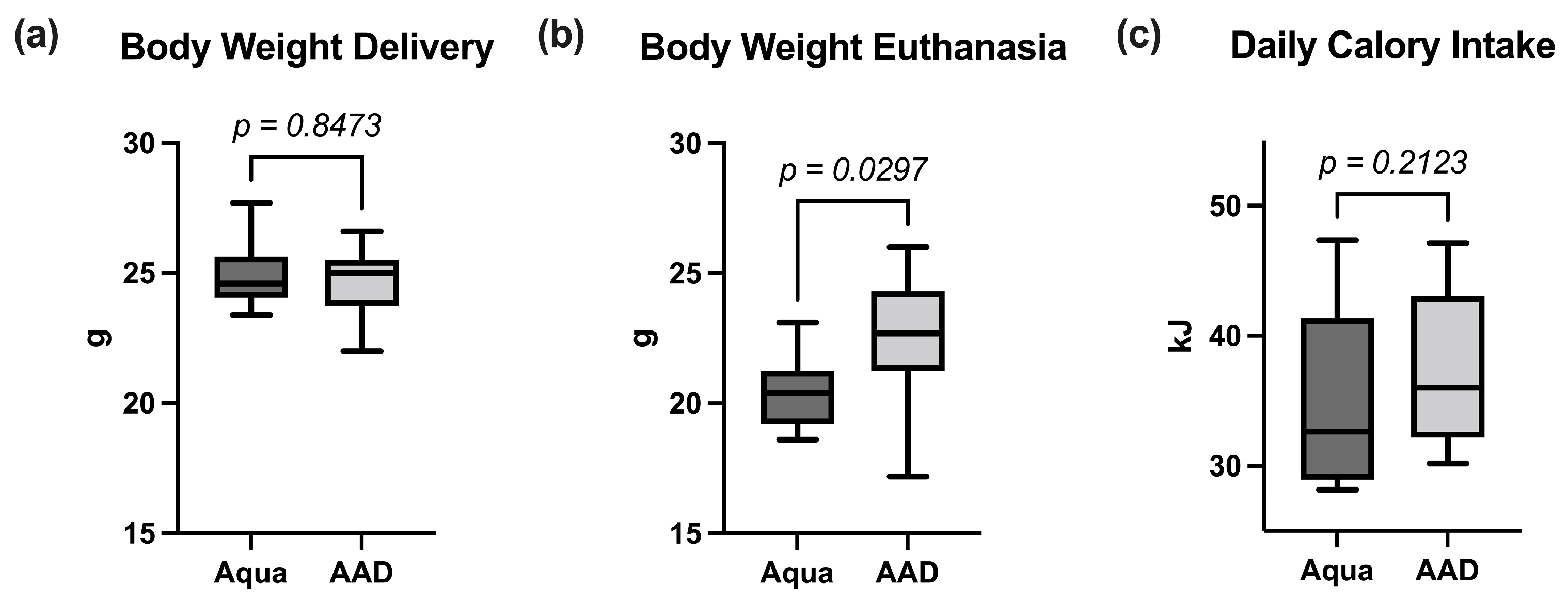
3.1. Inflammatory Response
3.2. Lipolysis Markers
3.3. Gut Permeability Assay, Bowel Wall Integrity, and Inflammation
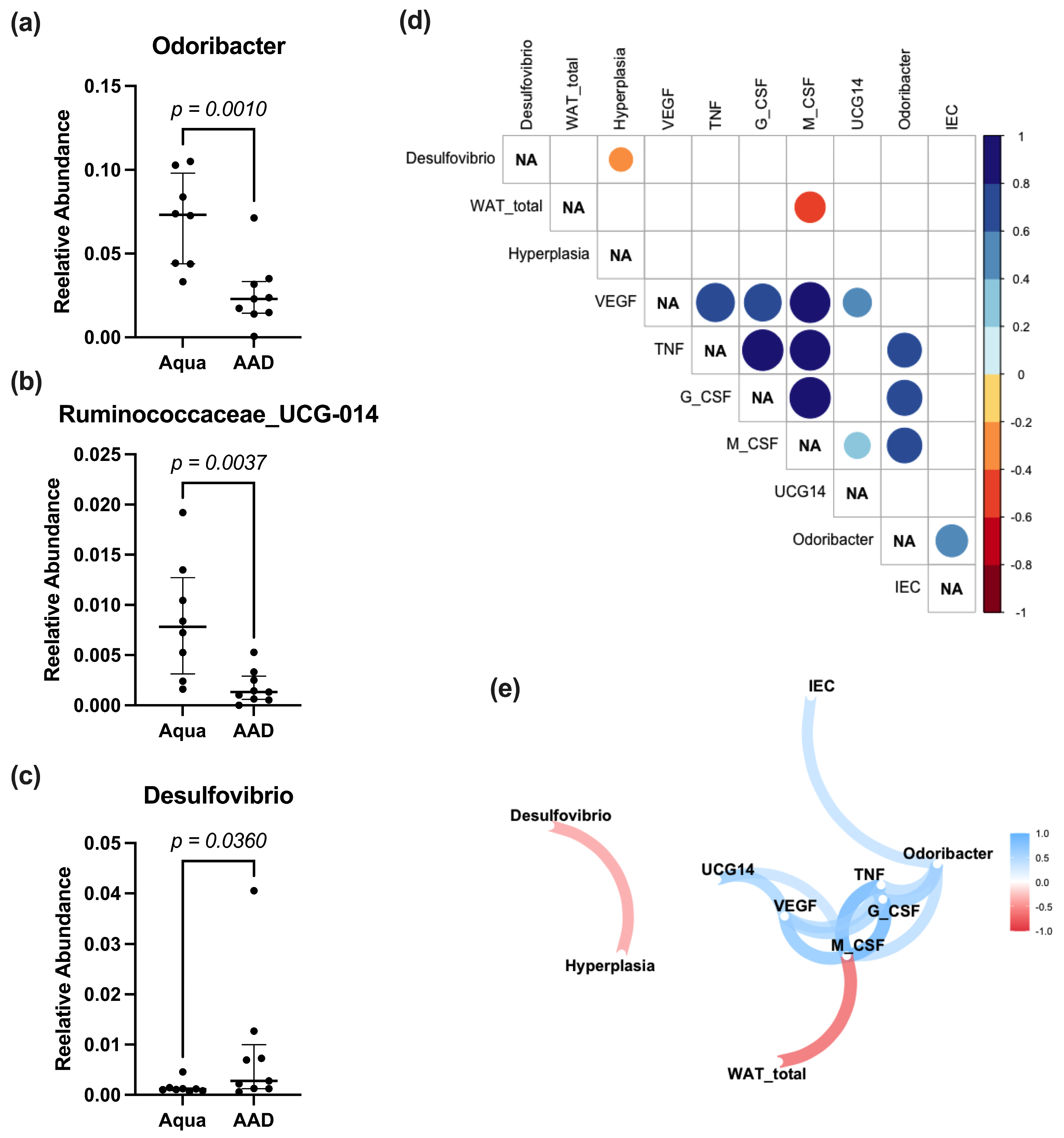
3.4. Fecal Microbiome Analysis
3.5. Fecal Volatile Organic Compound Profile
4. Discussion
Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davila, M.; Bresalier, R.S. Gastrointestinal complications of oncologic therapy. Nat. Clin. Pract. Gastroenterol. Hepatol. 2008, 5, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.L.; Geier, M.S.; Yazbeck, R.; Torres, D.M.; Butler, R.N.; Howarth, G.S. Lactobacillus fermentum BR11 and fructo-oligosaccharide partially reduce jejunal inflammation in a model of intestinal mucositis in rats. Nutr. Cancer 2008, 60, 757–767. [Google Scholar] [CrossRef]
- Ciorba, M.A.; Hallemeier, C.L.; Stenson, W.F.; Parikh, P.J. Probiotics to prevent gastrointestinal toxicity from cancer therapy: An interpretive review and call to action. Curr. Opin. Support. Palliat. Care 2015, 9, 157–162. [Google Scholar] [CrossRef]
- Sonis, S.T. The pathobiology of mucositis. Nat. Rev. Cancer 2004, 4, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Keefe, D.M. Gastrointestinal mucositis: A new biological model. Support. Care Cancer 2004, 12, 6–9. [Google Scholar] [CrossRef]
- Miknevicius, P.; Zulpaite, R.; Leber, B.; Strupas, K.; Stiegler, P.; Schemmer, P. The Impact of Probiotics on Intestinal Mucositis during Chemotherapy for Colorectal Cancer: A Comprehensive Review of Animal Studies. Int. J. Mol. Sci. 2021, 22, 9347. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.J.; Keefe, D.M.; Clarke, J.M.; Regester, G.O.; Thompson, F.M.; Goland, G.J.; Edwards, B.G.; Cummins, A.G. The effect of keratinocyte growth factor on tumour growth and small intestinal mucositis after chemotherapy in the rat with breast cancer. Cancer Chemother. Pharmacol. 2002, 50, 53–58. [Google Scholar] [CrossRef]
- Gibson, R.J.; Keefe, D.M.; Thompson, F.M.; Clarke, J.M.; Goland, G.J.; Cummins, A.G. Effect of interleukin-11 on ameliorating intestinal damage after methotrexate treatment of breast cancer in rats. Dig. Dis. Sci. 2002, 47, 2751–2757. [Google Scholar] [CrossRef]
- Elting, L.S.; Cooksley, C.; Chambers, M.; Cantor, S.B.; Manzullo, E.; Rubenstein, E.B. The burdens of cancer therapy. Clinical and economic outcomes of chemotherapy-induced mucositis. Cancer 2003, 98, 1531–1539. [Google Scholar] [CrossRef]
- Stringer, A.M.; Gibson, R.J.; Bowen, J.M.; Keefe, D.M. Chemotherapy-induced modifications to gastrointestinal microflora: Evidence and implications of change. Curr. Drug Metab. 2009, 10, 79–83. [Google Scholar] [CrossRef]
- van Vliet, M.J.; Harmsen, H.J.; de Bont, E.S.; Tissing, W.J. The role of intestinal microbiota in the development and severity of chemotherapy-induced mucositis. PLoS Pathog. 2010, 6, e1000879. [Google Scholar] [CrossRef] [PubMed]
- Bron, P.A.; Kleerebezem, M.; Brummer, R.J.; Cani, P.D.; Mercenier, A.; MacDonald, T.T.; Garcia-Rodenas, C.L.; Wells, J.M. Can probiotics modulate human disease by impacting intestinal barrier function? Br. J. Nutr. 2017, 117, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, K.X.; Qu, J.M.; Wang, X.D. The changes induced by cyclophosphamide in intestinal barrier and microflora in mice. Eur. J. Pharmacol. 2013, 714, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Badgeley, A.; Anwar, H.; Modi, K.; Murphy, P.; Lakshmikuttyamma, A. Effect of probiotics and gut microbiota on anti-cancer drugs: Mechanistic perspectives. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188494. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Otte, J.M.; Podolsky, D.K. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G613–G626. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; Gonzalez, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Toumi, R.; Abdelouhab, K.; Rafa, H.; Soufli, I.; Raissi-Kerboua, D.; Djeraba, Z.; Touil-Boukoffa, C. Beneficial role of the probiotic mixture Ultrabiotique on maintaining the integrity of intestinal mucosal barrier in DSS-induced experimental colitis. Immunopharmacol. Immunotoxicol. 2013, 35, 403–409. [Google Scholar] [CrossRef]
- Viaud, S.; Daillere, R.; Boneca, I.G.; Lepage, P.; Langella, P.; Chamaillard, M.; Pittet, M.J.; Ghiringhelli, F.; Trinchieri, G.; Goldszmid, R.; et al. Gut microbiome and anticancer immune response: Really hot Sh*t! Cell Death Differ. 2015, 22, 199–214. [Google Scholar] [CrossRef]
- Nami, Y.; Haghshenas, B.; Haghshenas, M.; Abdullah, N.; Yari Khosroushahi, A. The Prophylactic Effect of Probiotic Enterococcus lactis IW5 against Different Human Cancer Cells. Front. Microbiol. 2015, 6, 1317. [Google Scholar] [CrossRef]
- Sharaf, L.K.; Sharma, M.; Chandel, D.; Shukla, G. Prophylactic intervention of probiotics (L. acidophilus, L. rhamnosus GG) and celecoxib modulate Bax-mediated apoptosis in 1,2-dimethylhydrazine-induced experimental colon carcinogenesis. BMC Cancer 2018, 18, 1111. [Google Scholar] [CrossRef] [PubMed]
- Castellani, C.; Singer, G.; Kaiser, M.; Obermüller, B.; Warncke, G.; Miekisch, W.; Kolb-Lenz, D.; Summer, G.; Pauer, T.M.; Elhaddad, A.; et al. The Effects of Neuroblastoma and Chemotherapy on Metabolism, Fecal Microbiome, Volatile Organic Compounds and Gut Barrier—A Murine Model of Human Neuroblastoma. Pediatr. Res. 2019, 85, 546–555. [Google Scholar] [CrossRef]
- Schweiger, M.; Eichmann, T.O.; Taschler, U.; Zimmermann, R.; Zechner, R.; Lass, A. Measurement of lipolysis. Methods Enzymol. 2014, 538, 171–193. [Google Scholar] [CrossRef] [PubMed]
- Adelman, D.; Murray, J.; Wu, T.; Mäki, M.; Green, P.; CP, K. Measuring change in small intestinal histology in patients with celiac disease. Am. J. Gastroenterol. 2018, 113, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kuhl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar] [PubMed]
- Klymiuk, I.; Bilgilier, C.; Stadlmann, A.; Thannesberger, J.; Kastner, M.T.; Hogenauer, C.; Puspok, A.; Biowski-Frotz, S.; Schrutka-Kolbl, C.; Thallinger, G.G.; et al. The Human Gastric Microbiome Is Predicated upon Infection with Helicobacter pylori. Front. Microbiol. 2017, 8, 2508. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Gruning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, A.; Trefz, P.; Fischer, S.; Klepik, K.; Walter, G.; Steffens, M.; Ziller, M.; Schubert, J.K.; Reinhold, P.; Kohler, H.; et al. In Vivo Volatile Organic Compound Signatures of Mycobacterium avium subsp. paratuberculosis. PLoS ONE 2015, 10, e0123980. [Google Scholar] [CrossRef] [PubMed]
- Miekisch, W.; Trefz, P.; Bergmann, A.; Schubert, J.K. Microextraction techniques in breath biomarker analysis. Bioanalysis 2014, 6, 1275–1291. [Google Scholar] [CrossRef]
- Kienesberger, B.; Obermuller, B.; Singer, G.; Mittl, B.; Grabherr, R.; Mayrhofer, S.; Heinl, S.; Stadlbauer, V.; Horvath, A.; Miekisch, W.; et al. (S)-Reutericyclin: Susceptibility Testing and In Vivo Effect on Murine Fecal Microbiome and Volatile Organic Compounds. Int. J. Mol. Sci. 2021, 22, 6424. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, T.; Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix; Version 0.92; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Kuhn, M.; Jackson, S.; Cimentada, J. Corrr: Correlations in R; Version 0.4.3; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; D’Agostino McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the {tidyverse}. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Pedersen, T.; Nicolae, B.; Francois, R. Farver: High Performance Colour Space Manipulation; Version 2.1.0; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef]
- Chmielewska, A.; Szajewska, H. Systematic review of randomised controlled trials: Probiotics for functional constipation. World J. Gastroenterol. 2010, 16, 69–75. [Google Scholar] [CrossRef]
- Demers, M.; Dagnault, A.; Desjardins, J. A randomized double-blind controlled trial: Impact of probiotics on diarrhea in patients treated with pelvic radiation. Clin. Nutr. 2014, 33, 761–767. [Google Scholar] [CrossRef]
- Di Gioia, D.; Aloisio, I.; Mazzola, G.; Biavati, B. Bifidobacteria: Their impact on gut microbiota composition and their applications as probiotics in infants. Appl. Microbiol. Biotechnol. 2014, 98, 563–577. [Google Scholar] [CrossRef]
- Hempel, S.; Newberry, S.J.; Maher, A.R.; Wang, Z.; Miles, J.N.; Shanman, R.; Johnsen, B.; Shekelle, P.G. Probiotics for the prevention and treatment of antibiotic-associated diarrhea: A systematic review and meta-analysis. JAMA 2012, 307, 1959–1969. [Google Scholar] [CrossRef]
- Isolauri, E.; Rautava, S.; Salminen, S. Probiotics in the development and treatment of allergic disease. Gastroenterol. Clin. N. Am. 2012, 41, 747–762. [Google Scholar] [CrossRef]
- McFarland, L.V. Meta-analysis of probiotics for the prevention of traveler’s diarrhea. Travel. Med. Infect. Dis. 2007, 5, 97–105. [Google Scholar] [CrossRef]
- Koning, C.J.; Jonkers, D.M.; Stobberingh, E.E.; Mulder, L.; Rombouts, F.M.; Stockbrugger, R.W. The effect of a multispecies probiotic on the intestinal microbiota and bowel movements in healthy volunteers taking the antibiotic amoxycillin. Am. J. Gastroenterol. 2008, 103, 178–189. [Google Scholar] [CrossRef]
- Lang, F.C. Use of a multi-species probiotic for the prevention of antibiotic associated diarrhea. Nutra Foods 2010, 9, 27–31. [Google Scholar]
- van Wietmarschen, H.A.; Busch, M.; van Oostveen, A.; Pot, G.; Jong, M.C. Probiotics use for antibiotic-associated diarrhea: A pragmatic participatory evaluation in nursing homes. BMC Gastroenterol. 2020, 20, 151. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, H.M.; Koning, C.J.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.M.; Stringer, A.M.; Gibson, R.J.; Yeoh, A.S.; Hannam, S.; Keefe, D.M. VSL#3 probiotic treatment reduces chemotherapy-induced diarrhea and weight loss. Cancer Biol. Ther. 2007, 6, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Argiles, J.M.; Busquets, S.; Garcia-Martinez, C.; Lopez-Soriano, F.J. Mediators involved in the cancer anorexia-cachexia syndrome: Past, present, and future. Nutrition 2005, 21, 977–985. [Google Scholar] [CrossRef]
- Bastos, R.W.; Pedroso, S.H.; Vieira, A.T.; Moreira, L.M.; Franca, C.S.; Cartelle, C.T.; Arantes, R.M.; Generoso, S.V.; Cardoso, V.N.; Neves, M.J.; et al. Saccharomyces cerevisiae UFMG A-905 treatment reduces intestinal damage in a murine model of irinotecan-induced mucositis. Benef. Microbes 2016, 7, 549–557. [Google Scholar] [CrossRef]
- Kato, S.; Hamouda, N.; Kano, Y.; Oikawa, Y.; Tanaka, Y.; Matsumoto, K.; Amagase, K.; Shimakawa, M. Probiotic Bifidobacterium bifidum G9-1 attenuates 5-fluorouracil-induced intestinal mucositis in mice via suppression of dysbiosis-related secondary inflammatory responses. Clin. Exp. Pharmacol. Physiol. 2017, 44, 1017–1025. [Google Scholar] [CrossRef]
- Mi, H.; Dong, Y.; Zhang, B.; Wang, H.; Peter, C.C.K.; Gao, P.; Fu, H.; Gao, Y. Bifidobacterium Infantis Ameliorates Chemotherapy-Induced Intestinal Mucositis Via Regulating T Cell Immunity in Colorectal Cancer Rats. Cell Physiol. Biochem. 2017, 42, 2330–2341. [Google Scholar] [CrossRef]
- Tang, Y.; Wu, Y.; Huang, Z.; Dong, W.; Deng, Y.; Wang, F.; Li, M.; Yuan, J. Administration of probiotic mixture DM#1 ameliorated 5-fluorouracil-induced intestinal mucositis and dysbiosis in rats. Nutrition 2017, 33, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Yuan, K.T.; Yu, H.L.; Feng, W.D.; Chong, P.; Yang, T.; Xue, C.L.; Yu, M.; Shi, H.P. Bifidobacterium infantis has a beneficial effect on 5-fluorouracil-induced intestinal mucositis in rats. Benef. Microbes 2015, 6, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Lass, A.; Zimmermann, R.; Oberer, M.; Zechner, R. Lipolysis—A highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Prog. Lipid Res. 2011, 50, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.S.; Jessen, N.; Jorgensen, J.O.; Moller, N.; Lund, S. Dissecting adipose tissue lipolysis: Molecular regulation and implications for metabolic disease. J. Mol. Endocrinol. 2014, 52, R199–R222. [Google Scholar] [CrossRef]
- Li, T.; Guo, W.; Zhou, Z. Adipose Triglyceride Lipase in Hepatic Physiology and Pathophysiology. Biomolecules 2021, 12, 57. [Google Scholar] [CrossRef]
- Buhl, M.; Bosnjak, E.; Vendelbo, M.H.; Gjedsted, J.; Nielsen, R.R.; K.-Hafstrøm, T.; Vestergaard, E.T.; Jessen, N.; Tonnesen, E.; Moller, A.B.; et al. Direct effects of locally administered lipopolysaccharide on glucose, lipid, and protein metabolism in the placebo-controlled, bilaterally infused human leg. J. Clin. Endocrinol. Metab. 2013, 98, 2090–2099. [Google Scholar] [CrossRef]
- Fruhbeck, G.; Mendez-Gimenez, L.; Fernandez-Formoso, J.A.; Fernandez, S.; Rodriguez, A. Regulation of adipocyte lipolysis. Nutr. Res. Rev. 2014, 27, 63–93. [Google Scholar] [CrossRef]
- Huang, L.; Chiang Chiau, J.S.; Cheng, M.L.; Chan, W.T.; Jiang, C.B.; Chang, S.W.; Yeung, C.Y.; Lee, H.C. SCID/NOD mice model for 5-FU induced intestinal mucositis: Safety and effects of probiotics as therapy. Pediatr. Neonatol. 2019, 60, 252–260. [Google Scholar] [CrossRef]
- Yeung, C.Y.; Chan, W.T.; Jiang, C.B.; Cheng, M.L.; Liu, C.Y.; Chang, S.W.; Chiang Chiau, J.S.; Lee, H.C. Correction: Amelioration of Chemotherapy-Induced Intestinal Mucositis by Orally Administered Probiotics in a Mouse Model. PLoS ONE 2015, 10, e0141402. [Google Scholar] [CrossRef]
- Bibiloni, R.; Fedorak, R.N.; Tannock, G.W.; Madsen, K.L.; Gionchetti, P.; Campieri, M.; De Simone, C.; Sartor, R.B. VSL#3 probiotic-mixture induces remission in patients with active ulcerative colitis. Am. J. Gastroenterol. 2005, 100, 1539–1546. [Google Scholar] [CrossRef]
- Shen, J.; Zuo, Z.X.; Mao, A.P. Effect of probiotics on inducing remission and maintaining therapy in ulcerative colitis, Crohn’s disease, and pouchitis: Meta-analysis of randomized controlled trials. Inflamm. Bowel Dis. 2014, 20, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front. Biosci. 2009, 14, 2765–2778. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.A.; Koren, O.; Goodrich, J.K.; Johansson, M.E.; Nalbantoglu, I.; Aitken, J.D.; Su, Y.; Chassaing, B.; Walters, W.A.; Gonzalez, A.; et al. Transient inability to manage proteobacteria promotes chronic gut inflammation in TLR5-deficient mice. Cell Host Microbe 2012, 12, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Boyd, J.H.; Divangahi, M.; Yahiaoui, L.; Gvozdic, D.; Qureshi, S.; Petrof, B.J. Toll-like receptors differentially regulate CC and CXC chemokines in skeletal muscle via NF-kappaB and calcineurin. Infect. Immun. 2006, 74, 6829–6838. [Google Scholar] [CrossRef]
- Dai, C.; Zhao, D.H.; Jiang, M. VSL#3 probiotics regulate the intestinal epithelial barrier in vivo and in vitro via the p38 and ERK signaling pathways. Int. J. Mol. Med. 2012, 29, 202–208. [Google Scholar] [CrossRef]
- Mennigen, R.; Nolte, K.; Rijcken, E.; Utech, M.; Loeffler, B.; Senninger, N.; Bruewer, M. Probiotic mixture VSL#3 protects the epithelial barrier by maintaining tight junction protein expression and preventing apoptosis in a murine model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1140–G1149. [Google Scholar] [CrossRef]
- Touchefeu, Y.; Montassier, E.; Nieman, K.; Gastinne, T.; Potel, G.; Bruley des Varannes, S.; Le Vacon, F.; de La Cochetiere, M.F. Systematic review: The role of the gut microbiota in chemotherapy- or radiation-induced gastrointestinal mucositis—Current evidence and potential clinical applications. Aliment. Pharmacol. Ther. 2014, 40, 409–421. [Google Scholar] [CrossRef]
- Chang, C.W.; Liu, C.Y.; Lee, H.C.; Huang, Y.H.; Li, L.H.; Chiau, J.C.; Wang, T.E.; Chu, C.H.; Shih, S.C.; Tsai, T.H.; et al. Lactobacillus casei Variety rhamnosus Probiotic Preventively Attenuates 5-Fluorouracil/Oxaliplatin-Induced Intestinal Injury in a Syngeneic Colorectal Cancer Model. Front. Microbiol. 2018, 9, 983. [Google Scholar] [CrossRef]
- Yue, Y.; He, Z.; Zhou, Y.; Ross, R.P.; Stanton, C.; Zhao, J.; Zhang, H.; Yang, B.; Chen, W. Lactobacillus plantarum relieves diarrhea caused by enterotoxin-producing Escherichia coli through inflammation modulation and gut microbiota regulation. Food Funct. 2020, 11, 10362–10374. [Google Scholar] [CrossRef]
- Zhou, J.; Li, M.; Chen, Q.; Li, X.; Chen, L.; Dong, Z.; Zhu, W.; Yang, Y.; Liu, Z.; Chen, Q. Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery. Nat. Commun. 2022, 13, 3432. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C.; Mehrotra, S.; Loo, V.G.; Dewar, K.; Manges, A.R. Excretion of Host DNA in Feces Is Associated with Risk of Clostridium difficile Infection. J. Immunol. Res. 2015, 2015, 246203. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.C.; Wang, Y.N.; Yan, P.G.; Li, Y.H.; Wang, H.Y.; Qian, J.M.; Li, J.N. Effect of VSL#3 and S.Boulardii on intestinal microbiota in mice with acute colitis. Zhonghua Yi Xue Za Zhi 2019, 99, 1735–1742. [Google Scholar] [CrossRef] [PubMed]
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef]
- Hall, A.B.; Yassour, M.; Sauk, J.; Garner, A.; Jiang, X.; Arthur, T.; Lagoudas, G.K.; Vatanen, T.; Fornelos, N.; Wilson, R.; et al. A novel Ruminococcus gnavus clade enriched in inflammatory bowel disease patients. Genome Med. 2017, 9, 103. [Google Scholar] [CrossRef]
- Rajilic-Stojanovic, M.; Jonkers, D.M.; Salonen, A.; Hanevik, K.; Raes, J.; Jalanka, J.; de Vos, W.M.; Manichanh, C.; Golic, N.; Enck, P.; et al. Intestinal microbiota and diet in IBS: Causes, consequences, or epiphenomena? Am. J. Gastroenterol. 2015, 110, 278–287. [Google Scholar] [CrossRef]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef]
- Rios, J.L.; Bomhof, M.R.; Reimer, R.A.; Hart, D.A.; Collins, K.H.; Herzog, W. Protective effect of prebiotic and exercise intervention on knee health in a rat model of diet-induced obesity. Sci. Rep. 2019, 9, 3893. [Google Scholar] [CrossRef]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Ahmed, I.; Greenwood, R.; Costello Bde, L.; Ratcliffe, N.M.; Probert, C.S. An investigation of fecal volatile organic metabolites in irritable bowel syndrome. PLoS ONE 2013, 8, e58204. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Param. | Unit | Aqua | 10 AAD | |||
|---|---|---|---|---|---|---|
| Median | 25–75 Perc. | Median | 25–75 Perc. | p-Value | ||
| INF-γ | pg/mL | 13.1 | 3.0–60.3 | 3.0 | 3.0–11.2 | 0.160 |
| IL-1α | pg/mL | 18.1 | 6.3–813.8 | 1.0 | 1.0–8.0 | 0.022 |
| IL-1β | pg/mL | 7.2 | 0.4–765.9 | 0.4 | 0.4–1.7 | 0.021 |
| IL-6 | pg/mL | 862.1 | 263.8–121,076 | 27.5 | 27.5–307.9 | 0.011 |
| IL-10 | pg/mL | 7.9 | 1.9–236.3 | 0.1 | 0.1–1.6 | 0.018 |
| IL-15 | pg/mL | 8.1 | 2.6–30.9 | 0.3 | 0.3–2.7 | 0.018 |
| IL-17 | pg/mL | 17.0 | 1.6–38.6 | 1.6 | 1.6–4.7 | 0.056 |
| MIP-1α | pg/mL | 2.2 | 0.1–167.5 | 0.1 | 0.1–0.1 | 0.076 |
| MIP-1β | pg/mL | 1.4 | 0.9–119.1 | 0.7 | 0.6–3.8 | 0.042 |
| MIP-2 | pg/mL | 99.2 | 32.9–19,710 | 26.0 | 26.0–50.7 | 0.024 |
| MCP-1 | pg/mL | 552.6 | 49.6–21,255 | 51.8 | 28.3–1161 | 0.385 |
| TNF-α | pg/mL | 59.9 | 10.9–276.5 | 1.7 | 1.7–20.8 | 0.024 |
| G-CSF | pg/mL | 3121 | 1421–4864 | 510 | 51.1–1636 | 0.021 |
| GM-CSF | pg/mL | 5.1 | 3.4–48.7 | 3.4 | 3.4–3.4 | 0.029 |
| M-CSF | pg/mL | 1.4 | 0.4–3.8 | 0.3 | 0.2–0.4 | 0.046 |
| MPO | ng/mL | 118.4 | 60.6–283.9 | 48.9 | 29.6–68.7 | 0.040 |
| NE | ng/mL | 30.9 | 6.9–30.9 | 4.3 | 2.2–11.6 | 0.061 |
| LBP | ng/mL | 271.1 | 74.3–2764 | 125.2 | 56.5–344.4 | 0.291 |
| VEGF | ng/mL | 16.2 | 2.2–32.5 | 0.6 | 0.1–4.1 | 0.047 |
| TGF-β1 | pg/mL | 26,143 | 17,586–31,665 | 34,677 | 29,160–44,524 | 0.019 |
| TGF-β2 | pg/mL | 986.4 | 696.2–1232 | 1141 | 911.6–1330 | 0.436 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obermüller, B.; Singer, G.; Kienesberger, B.; Mittl, B.; Stadlbauer, V.; Horvath, A.; Miekisch, W.; Fuchs, P.; Schweiger, M.; Pajed, L.; et al. Probiotic OMNi-BiOTiC® 10 AAD Reduces Cyclophosphamide-Induced Inflammation and Adipose Tissue Wasting in Mice. Nutrients 2023, 15, 3655. https://doi.org/10.3390/nu15163655
Obermüller B, Singer G, Kienesberger B, Mittl B, Stadlbauer V, Horvath A, Miekisch W, Fuchs P, Schweiger M, Pajed L, et al. Probiotic OMNi-BiOTiC® 10 AAD Reduces Cyclophosphamide-Induced Inflammation and Adipose Tissue Wasting in Mice. Nutrients. 2023; 15(16):3655. https://doi.org/10.3390/nu15163655
Chicago/Turabian StyleObermüller, Beate, Georg Singer, Bernhard Kienesberger, Barbara Mittl, Vanessa Stadlbauer, Angela Horvath, Wolfram Miekisch, Patricia Fuchs, Martina Schweiger, Laura Pajed, and et al. 2023. "Probiotic OMNi-BiOTiC® 10 AAD Reduces Cyclophosphamide-Induced Inflammation and Adipose Tissue Wasting in Mice" Nutrients 15, no. 16: 3655. https://doi.org/10.3390/nu15163655
APA StyleObermüller, B., Singer, G., Kienesberger, B., Mittl, B., Stadlbauer, V., Horvath, A., Miekisch, W., Fuchs, P., Schweiger, M., Pajed, L., Till, H., & Castellani, C. (2023). Probiotic OMNi-BiOTiC® 10 AAD Reduces Cyclophosphamide-Induced Inflammation and Adipose Tissue Wasting in Mice. Nutrients, 15(16), 3655. https://doi.org/10.3390/nu15163655