
Gut Microbiota and Critically Ill Patients: Immunity and Its Modulation via Probiotics and Immunonutrition

 ,
,  ,
,  ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Methods
3. Gut Dysbiosis as the Origin of Systemic Inflammatory Response to Acute Critical Damages
4. Gut Microbiota and Immune System Disequilibrium in Critical Patient
5. Gut Dysbiosis Modulation through Probiotics and Immunonutrition
5.1. Probiotics and Their Derivatives
5.1.1. Probiotics Definition and Effects
5.1.2. Probiotics and Diarrhea
5.1.3. Fecal Microbiota Transplantation and Critically Ill Patient
5.1.4. Probiotics and Infections
5.1.5. Limitations to Probiotics’ Use in ICU Patients
5.2. Promising Effectiveness of Immunonutrition
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef]
- Cunningham, R.M.; Walton, M.A.; Carter, P.M. The Major Causes of Death in Children and Adolescents in the United States. N. Engl. J. Med. 2018, 379, 2468–2475. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Ojima, M.; Ogura, H. Gut Microbiota and Probiotics/Synbiotics for Modulation of Immunity in Critically Ill Patients. Nutrients 2021, 13, 2439. [Google Scholar] [CrossRef] [PubMed]
- Stoecklein, V.M.; Osuka, A.; Lederer, J.A. Trauma equals danger-Damage control by the immune system. J. Leukoc. Biol. 2012, 92, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, M.D.; Moore, F.A. Persistent Inflammation, Immunosuppression, and Catabolism: Evolution of Multiple Organ Dysfunction. Surg. Infect. 2016, 17, 167–172. [Google Scholar] [CrossRef] [PubMed]
- De Rose, D.U.; Pace, P.G.; Ceccherini-Silberstein, F.; Dotta, A.; Andreoni, M.; Sarmati, L.; Iannetta, M. T Lymphocyte Subset Counts and Interferon-Gamma Production in Adults and Children with COVID-19: A Narrative Review. J. Pers. Med. 2023, 13, 755. [Google Scholar] [CrossRef]
- Otani, S.; Coopersmith, C.M. Gut integrity in critical illness. J. Intensiv. Care 2019, 7, 17. [Google Scholar] [CrossRef]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 372, 71. [Google Scholar]
- Shanahan, F.; Ghosh, T.S.; O’Toole, P.W. The Healthy Microbiome-What Is the Definition of a Healthy Gut Microbiome? Gastroenterology 2021, 160, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Scarpellini, E.; Luzza, F. The role of gut microbiota in gastrointestinal diseases: The heart of the matter. Minerva Gastroenterol. 2021, 67, 312–313. [Google Scholar] [CrossRef]
- Scarpellini, E.; Basilico, M.; Rinninella, E.; Carbone, F.; Schol, J.; Rasetti, C.; Abenavoli, L.; Santori, P. Probiotics and gut health. Minerva Gastroenterol. 2021, 67, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Yokoyama, Y.; Nishio, H.; Ebata, T.; Sugawara, G.; Asahara, T.; Nomoto, K.; Nagino, M. Intraoperative Bacterial Translocation Detected by Bacterium-Specific Ribosomal RNA-Targeted Reverse-Transcriptase Polymerase Chain Reaction for the Mesenteric Lymph Node Strongly Predicts Postoperative Infectious Complications After Major Hepatectomy for Biliary Malignancies. Ann. Surg. 2010, 252, 1013–1019. [Google Scholar]
- Iapichino, G.; Callegari, M.L.; Marzorati, S.; Cigada, M.; Corbella, D.; Ferrari, S.; Morelli, L. Impact of antibiotics on the gut microbiota of critically ill patients. J. Med. Microbiol. 2008, 57, 1007–1014. [Google Scholar] [CrossRef]
- Zaborin, A.; Smith, D.; Garfield, K.; Quensen, J.; Shakhsheer, B.; Kade, M.; Tirrell, M.; Tiedje, J.; Gilbert, J.A.; Zaborina, O.; et al. Membership and behavior of ultra-low-diversity pathogen communities present in the gut of humans during prolonged critical illness. mBio 2014, 5, e01361-14. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- Morowitz, M.J.; Carlisle, E.M.; Alverdy, J.C. Contributions of Intestinal Bacteria to Nutrition and Metabolism in the Critically Ill. Surg. Clin. N. Am. 2011, 91, 771–785. [Google Scholar] [CrossRef]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D.; et al. Host-derived nitrate boosts growth of E. coli in the inflamed gut. Science 2013, 339, 708–711. [Google Scholar] [CrossRef]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H.; et al. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology 2014, 147, 1055–1063.e8. [Google Scholar] [CrossRef]
- Lupp, C.; Robertson, M.L.; Wickham, M.E.; Sekirov, I.; Champion, O.L.; Gaynor, E.C.; Finlay, B.B. Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of Enterobacteriaceae. Cell Host Microbe 2007, 2, 204. [Google Scholar] [CrossRef]
- Grootjans, J.; Lenaerts, K.; Derikx, J.P.; Matthijsen, R.A.; de Bruïne, A.P.; van Bijnen, A.A.; van Dam, R.M.; Dejong, C.H.; Buurman, W.A. Human intestinal ischemia-reperfusion-induced inflammation characterized: Experiences from a new translational model. Am. J. Pathol. 2010, 176, 2283–2291. [Google Scholar] [CrossRef]
- Shimizu, K.; Ogura, H.; Hamasaki, T.; Goto, M.; Tasaki, O.; Asahara, T.; Nomoto, K.; Morotomi, M.; Matsushima, A.; Kuwagata, Y.; et al. Altered Gut Flora Are Associated with Septic Complications and Death in Critically Ill Patients with Systemic Inflammatory Response Syndrome. Dig. Dis. Sci. 2010, 56, 1171–1177. [Google Scholar] [CrossRef]
- Marascio, N.; Scarlata, G.G.M.; Romeo, F.; Cicino, C.; Trecarichi, E.M.; Quirino, A.; Torti, C.; Matera, G.; Russo, A. The Role of Gut Microbiota in the Clinical Outcome of Septic Patients: State of the Art and Future Perspectives. Int. J. Mol. Sci. 2023, 24, 9307. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, Ethnicity or Subsistence-Specific Variations in Human Microbiome Composition and Diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [PubMed]
- Ojima, M.; Motooka, D.; Shimizu, K.; Gotoh, K.; Shintani, A.; Yoshiya, K.; Nakamura, S.; Ogura, H.; Iida, T.; Shimazu, T. Metagenomic Analysis Reveals Dynamic Changes of Whole Gut Microbiota in the Acute Phase of Intensive Care Unit Patients. Dig. Dis. Sci. 2016, 61, 1628–1634. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa Takashi, M.; Asahara, T.; Henzan, N.; Murakami, H.; Yamamoto, H.; Mukai, N.; Minami, Y.; Sugano, M.; Kubota, N.; Uegaki, S.; et al. Dramatic Changes of the Gut Flora Immediately After Severe and Sudden Insults. Dig. Dis. Sci. 2011, 56, 2361–2365. [Google Scholar] [CrossRef]
- Ojima, M.; Shimizu, K.; Motooka, D.; Ishihara, T.; Nakamura, S.; Shintani, A.; Ogura, H.; Iida, T.; Yoshiya, K.; Shimazu, T. Gut Dysbiosis Associated with Antibiotics and Disease Severity and Its Relation to Mortality in Critically Ill Patients. Dig. Dis. Sci. 2021, 37, 2420–2432. [Google Scholar] [CrossRef]
- Moore, E.E.; Moore, F.A.; Franciose, R.J.; Kim, F.J.; Biffl, W.L.; Banerjee, A. The Postischemic Gut Serves as a Priming Bed for Circulating Neutrophils that Provoke Multiple Organ Failure. J. Trauma Inj. Infect. Crit. Care 1994, 37, 881–887. [Google Scholar] [CrossRef]
- Eaves-Pyles, T.; Alexander, J.W. Rapid and Prolonged Impairment of Gut Barrier Function after Thermal Injury in Mice. Shock 1998, 9, 95–100. [Google Scholar] [CrossRef]
- Magnotti, L.J.; Upperman, J.S.; Xu, D.-Z.; Lu, Q.; Deitch, E.A. Gut-Derived Mesenteric Lymph but not Portal Blood Increases Endothelial Cell Permeability and Promotes Lung Injury After Hemorrhagic Shock. Ann. Surg. 1998, 228, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Yoseph, B.P.; Klingensmith, N.J.; Liang, Z.; Breed, E.; Burd, E.M.; Mittal, R.; Dominguez, J.A.; Petrie, B.; Ford, M.L.; Coopersmith, C.M. Mechanisms of Intestinal Barrier Dysfunction in Sepsis. Shock 2016, 46, 52–59. [Google Scholar] [CrossRef]
- O’Boyle, C.J.; MacFie, J.; Mitchell, C.J.; Johnstone, D.; Sagar, P.M.; Sedman, P.C. Microbiology of bacterial translocation in humans. Gut 1998, 42, 29–35. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Vollaard, E.J.; Clasener, H.A. Colonization resistance. Antimicrob. Agents Chemother. 1994, 38, 409–414. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nat. Cell Biol. 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Tanoue, T.; Morita, S.; Plichta, D.R.; Skelly, A.N.; Suda, W.; Sugiura, Y.; Narushima, S.; Vlamakis, H.; Motoo, I.; Sugita, K.; et al. A defined commensal consortium elicits CD8 T cells and anti-cancer immunity. Nature 2019, 565, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nat. Cell Biol. 2016, 535, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Sepsis-induced immunosuppression: From cellular dysfunctions to immunotherapy. Nat. Rev. Immunol. 2013, 13, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Peled, J.U.; Gomes, A.L.C.; Devlin, S.M.; Littmann, E.R.; Taur, Y.; Sung, A.D.; Weber, D.; Hashimoto, D.; Slingerland, A.E.; Slingerland, J.B.; et al. Microbiota as Predictor of Mortality in Allogeneic Hematopoietic-Cell Transplantation. N. Engl. J. Med. 2020, 382, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Tan, C.; Zhu, J.; Zeng, X.; Gao, X.; Wu, Q.; Chen, Q.; Wang, H.; Zhou, H.; He, Y.; et al. Dysbiosis of the intestinal microbiota in neurocritically ill patients and the risk for death. Crit Care 2019, 23, 195. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Guarner, F.; Guerrant, R.; Holt, P.; Quigley, E.M.M.; Sartor, R.B.; Sherman, P.; Mayer, E.A. An update on the use and investigation of probiotics in health and disease. Gut 2013, 62, 787–796. [Google Scholar] [CrossRef]
- Shimizu, K.; Ogura, H.; Asahara, T.; Nomoto, K.; Morotomi, M.; Tasaki, O.; Matsushima, A.; Kuwagata, Y.; Shimazu, T.; Sugimoto, H. Probiotic/Synbiotic Therapy for Treating Critically Ill Patients from a Gut Microbiota Perspective. Dig. Dis. Sci. 2012, 58, 23–32. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Genet. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Scarpellini, E.; Fagoonee, S.; Rinninella, E.; Rasetti, C.; Aquila, I.; Larussa, T.; Ricci, P.; Luzza, F.; Abenavoli, L. Gut Microbiota and Liver Interaction through Immune System Cross-Talk: A Comprehensive Review at the Time of the SARS-CoV-2 Pandemic. J. Clin. Med. 2020, 9, 2488. [Google Scholar] [CrossRef] [PubMed]
- Schlee, M.; Wehkamp, J.; Altenhoefer, A.; Oelschlaeger, T.A.; Stange, E.F.; Fellermann, K. Induction of human beta-defensin 2 by the probiotic Escherichia coli Nissle 1917 is mediated through flagellin. Infect. Immun. 2007, 75, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.B.; Davis, K.M.; Lysenko, E.S.; Zhou, A.Y.; Yu, Y.; Weiser, J.N. Recognition of peptidoglycan from the microbiota by Nod1 enhances systemic innate immunity. Nat. Med. 2010, 16, 228–231. [Google Scholar] [CrossRef]
- Asahara, T.; Shimizu, K.; Nomoto, K.; Hamabata, T.; Ozawa, A.; Takeda, Y. Probiotic Bifidobacteria Protect Mice from Lethal Infection with Shiga Toxin-Producing Escherichia coli O157:H7. Infect. Immun. 2004, 72, 2240–2247. [Google Scholar] [CrossRef] [PubMed]
- Yasui, H.; Kiyoshima, J.; Hori, T. Reduction of Influenza Virus Titer and Protection against Influenza Virus Infection in Infant Mice Fed Lactobacillus casei Shirota. Clin. Vaccine Immunol. 2004, 11, 675–679. [Google Scholar] [CrossRef]
- Hagihara, M.; Kuroki, Y.; Ariyoshi, T.; Higashi, S.; Fukuda, K.; Yamashita, R.; Matsumoto, A.; Mori, T.; Mimura, K.; Yamaguchi, N.; et al. Clostridium butyricum Modulates the Microbiome to Protect Intestinal Barrier Function in Mice with Antibiotic-Induced Dysbiosis. iScience 2020, 23, 100772. [Google Scholar] [CrossRef]
- Khailova, L.; Baird, C.H.; Rush, A.A.; McNamee, E.N.; Wischmeyer, P.E. Lactobacillus rhamnosus GG improves outcome in experimental pseudomonas aeruginosa pneumonia: Potential role of regulatory T cells. Shock 2013, 40, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Tirlapur, N.; Puthucheary, Z.A.; Cooper, J.A.; Sanders, J.; Coen, P.G.; Moonesinghe, S.R.; Wilson, A.P.; Mythen, M.G.; Montgomery, H.E. Diarrhoea in the critically ill is common, associated with poor outcome and rarely due to Clostridium difficile. Sci. Rep. 2016, 6, 24691. [Google Scholar] [CrossRef]
- Hickson, M.; D’Souza, A.L.; Muthu, N.; Rogers, T.; Want, S.; Rajkumar, C.; Bulpitt, C.J. Use of probiotic Lactobacillus preparation to prevent diarrhoea associated with antibiotics: Randomised double blind placebo controlled trial. BMJ 2007, 335, 80. [Google Scholar] [CrossRef]
- D’Souza, A.L.; Rajkumar, C.; Cooke, J.; Bulpitt, C.J. Probiotics in prevention of antibiotic associated diarrhoea: Meta-analysis. BMJ 2002, 324, 1361. [Google Scholar] [CrossRef] [PubMed]
- Bleichner, G.; Blehaut, H.; Mentec, H.; Moyse, D. Saccharomyces boulardii prevents diarrhea in critically ill tube-fed patients. Intensiv. Care Med. 1997, 23, 517–523. [Google Scholar] [CrossRef]
- Alagna, L.; Haak, B.W.; Gori, A. Fecal microbiota transplantation in the ICU: Perspectives on future implementations. Intensiv. Care Med. 2019, 45, 998–1001. [Google Scholar] [CrossRef]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; E Coffin, S.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, 987–994. [Google Scholar] [CrossRef]
- Wurm, P.; Spindelboeck, W.; Krause, R.; Plank, J.; Fuchs, G.; Bashir, M.; Petritsch, W.; Halwachs, B.; Langner, C.; Hogenauer, C.; et al. Antibiotic-Associated Apoptotic Enterocolitis in the Absence of a Defined Pathogen: The Role of Intestinal Microbiota Depletion. Crit. Care Med. 2017, 45, e600–e606. [Google Scholar] [CrossRef]
- Cibulková, I.; Řehořová, V.; Hajer, J.; Duška, F. Fecal Microbial Transplantation in Critically Ill Patients-Structured Review and Perspectives. Biomolecules 2021, 11, 1459. [Google Scholar] [CrossRef]
- Cheng, Y.W.; Phelps, E.; Nemes, S.; Rogers, N.; Sagi, S.; Bohm, M.; El-Halabi, M.; Allegretti, J.R.; Kassam, Z.; Xu, H.; et al. Fecal Microbiota Transplant Decreases Mortality in Patients with Refractory Severe or Fulminant Clostridioides difficile Infection. Clin. Gastroenterol. Hepatol. 2020, 18, 2234–2243.e1. [Google Scholar] [CrossRef]
- Alukal, J.; Dutta, S.K.; Surapaneni, B.K.; Le, M.; Tabbaa, O.; Phillips, L.; Mattar, M.C. Safety and Efficacy of Fecal Microbiota Transplant in 9 Critically Ill Patients with Severe and Complicated Clostridium difficile Infection with Impending Colectomy. J. Dig. Dis. 2019, 20, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Paramsothy, S.; Paramsothy, R.; Rubin, D.T.; Kamm, M.A.; Kaakoush, N.O.; Mitchell, H.M.; Castaño-Rodríguez, N. Faecal Microbiota Transplantation for Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis. J. Crohn’s Colitis 2017, 11, 1180–1199. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Yang, J.; Wang, J.; Yang, Y.; Huang, J.; Gong, H.; Cui, H.; Chen, D. Successful Treatment with Fecal Microbiota Transplantation in Patients with Multiple Organ Dysfunction Syndrome and Diarrhea Following Severe Sepsis. Crit. Care 2016, 20, 332. [Google Scholar] [CrossRef]
- Dai, M.; Liu, Y.; Chen, W.; Buch, H.; Shan, Y.; Chang, L.; Bai, Y.; Shen, C.; Zhang, X.; Huo, Y.; et al. Rescue Fecal Microbiota Transplantation for Antibiotic-Associated Diarrhea in Critically Ill Patients. Crit. Care 2019, 23, 284. [Google Scholar] [CrossRef]
- Kelly, C.R.; Ihunnah, C.; Fischer, M.; Khoruts, A.; Surawicz, C.; Afzali, A.; Aroniadis, O.; Barto, A.; Borody, T.; Giovanelli, A.; et al. Fecal Microbiota Transplant for Treatment of Clostridium difficile Infection in Immunocompromised Patients. Am. J. Gastroenterol. 2014, 109, 1065–1071. [Google Scholar] [CrossRef]
- Spindelboeck, W.; Schulz, E.; Uhl, B.; Kashofer, K.; Aigelsreiter, A.; Zinke-Cerwenka, W.; Mulabecirovic, A.; Kump, P.K.; Halwachs, B.; Gorkiewicz, G.; et al. Repeated Fecal Microbiota Transplantations Attenuate Diarrhea and Lead to Sustained Changes in the Fecal Microbiota in Acute, Refractory Gastrointestinal Graft-versus-Host-Disease. Haematologica 2017, 102, e210–e213. [Google Scholar] [CrossRef] [PubMed]
- Bilinski, J.; Grzesiowski, P.; Sorensen, N.; Madry, K.; Muszynski, J.; Robak, K.; Wroblewska, M.; Dzieciatkowski, T.; Dulny, G.; Dwilewicz-Trojaczek, J.; et al. Fecal Microbiota Transplantation in Patients with Blood Disorders Inhibits Gut Colonization with Antibiotic-Resistant Bacteria: Results of a Prospective, Single-Center Study. Clin. Infect. Dis. 2017, 65, 364–370. [Google Scholar] [CrossRef]
- Huttner, B.D.; de Lastours, V.; Wassenberg, M.; Maharshak, N.; Mauris, A.; Galperine, T.; Zanichelli, V.; Kapel, N.; Bellanger, A.; Olearo, F.; et al. A 5-Day Course of Oral Antibiotics Followed by Faecal Transplantation to Eradicate Carriage of Multidrug-Resistant Enterobacteriaceae: A Randomized Clinical Trial. Clin. Microbiol. Infect 2019, 25, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.D.; Liu, M.M.; Liang, X.; Hu, N.; Huang, W. Effects of perioperative supplementation with pro-/synbiotics on clinical outcomes in surgical patients: A meta-analysis with trial sequential analysis of randomized controlled trials. Clin. Nutr. 2018, 37, 505–515. [Google Scholar] [CrossRef]
- Gu, W.J.; Deng, T.; Gong, Y.Z.; Jing, R.; Liu, J.C. The effects of probiotics in early enteral nutrition on the outcomes of trauma: A meta-analysis of randomized controlled trials. JPEN J. Parenter. Enteral Nutr. 2013, 37, 310–317. [Google Scholar] [CrossRef]
- Sugawara, G.; Nagino, M.; Nishio, H.; Ebata, T.; Takagi, K.; Asahara, T.; Nomoto, K.; Nimura, Y. Perioperative synbiotic treatment to prevent postoperative infectious complications in biliary cancer surgery: A randomized controlled trial. Ann. Surg. 2006, 244, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Yamada, T.; Ogura, H.; Mohri, T.; Kiguchi, T.; Fujimi, S.; Asahara, T.; Ojima, M.; Ikeda, M.; Shimazu, T. Synbiotics modulate gut microbiota and reduce enteritis and ventilator-associated pneumonia in patients with sepsis: A randomized controlled trial. Crit. Care 2018, 22, 239. [Google Scholar] [CrossRef]
- Batra, P.; Soni, K.D.; Mathur, P. Efficacy of probiotics in the prevention of VAP in critically ill ICU patients: An updated systematic review and meta-analysis of randomized control trials. J. Intensive Care 2020, 8, 81. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Zhang, F.; Lui, G.C.; Yeoh, Y.K.; Li, A.Y.; Zhan, H.; Wan, Y.; Chung, A.C.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955.e8. [Google Scholar] [CrossRef]
- Ceccarelli, G.; Borrazzo, C.; Pinacchio, C.; Santinelli, L.; Innocenti, G.P.; Cavallari, E.N.; Celani, L.; Marazzato, M.; Alessandri, F.; Ruberto, F.; et al. Oral Bacteriotherapy in Patients With COVID-19: A Retrospective Cohort Study. Front. Nutr. 2021, 7, 613928. [Google Scholar] [CrossRef] [PubMed]
- Fruhwald, S.; Holzer, P.; Metzler, H. Intestinal motility disturbances in intensive care patients pathogenesis and clinical impact. Intensiv. Care Med. 2006, 33, 36–44. [Google Scholar] [CrossRef]
- Montejo, J.C. Enteral nutrition-related gastrointestinal complications in critically ill patients: A multicenter study. The Nutritional and Metabolic Working Group of the Spanish Society of Intensive Care Medicine and Coronary Units. Crit. Care Med. 1999, 27, 1447–1453. [Google Scholar] [CrossRef]
- Wang, X.; Gong, Z.; Wu, K.; Wang, B.; Yuang, Y. Gastrointestinal dysmotility in patients with acute pancreatitis. J. Gastroenterol. Hepatol. 2003, 18, 57–62. [Google Scholar] [CrossRef]
- Oláh, A.; Belágyi, T.; Issekutz, Á.; Gamal, M.E.; Bengmark, S. Randomized clinical trial of specific lactobacillus and fibre supplement to early enteral nutrition in patients with acute pancreatitis. BJS 2002, 89, 1103–1107. [Google Scholar] [CrossRef]
- Besselink, M.G.; van Santvoort, H.C.; Buskens, E.; Boermeester, M.A.; van Goor, H.; Timmerman, H.M.; Nieuwenhuijs, V.B.; Bollen, T.L.; van Ramshorst, B.; Witteman, B.J.; et al. Probiotic prophylaxis in predicted severe acute pancreatitis: A randomised, double-blind, placebo-controlled trial. Lancet 2008, 371, 651–659. [Google Scholar] [CrossRef]
- Derbyshire, E.; Delange, J. COVID-19: Is there a role for immuno-nutrition; particularly in the over 65s? BMJ Nutr. Prev. Health 2020, 3, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Jovic, T.H.; Ali, S.R.; Ibrahim, N.; Jessop, Z.M.; Tarassoli, S.P.; Dobbs, T.D.; Holford, P.; Thornton, C.A.; Whitaker, I.S. Could Vitamins Help in the Fight Against COVID-19? Nutrients 2020, 12, 2550. [Google Scholar] [CrossRef] [PubMed]
- Grimble, R.F. Basics in clinical nutrition: Immuno-nutrition—Nutrients which influence immunity: Effect and mechanism of action. Eur. E-J. Clin. Nutr. Metab. 2009, 4, e10–e13. [Google Scholar] [CrossRef]
- Matuszczyk, M.; Meglicka, M.; Landowski, P.; Czkwianianc, E.; Sordyl, B.; Szymańska, E.; Kierkuś, J. Oral exclusive enteral nutrition for induction of clinical remission, mucosal healing, and improvement of nutritional status and growth velocity in children with active Crohn’s disease—A prospective multicentre trial. Prz. Gastroenterol. 2021, 16, 346–351. [Google Scholar] [CrossRef]
- Grimble, R.F. Immunonutrition. Curr. Opin. Gastroenterol. 2005, 21, 216–222. [Google Scholar] [CrossRef]
- Cerantola, Y.; Hübner, M.; Grass, F.; Demartines, N.; Schäfer, M. Immunonutrition in gastrointestinal surgery. J. Br. Surg. 2011, 98, 37–48. [Google Scholar] [CrossRef]
- Mizock, B.A. Immunonutrition and critical illness: An update. Nutrition 2010, 26, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Marik, P.E.; Zaloga, G.P. Immunonutrition in critically ill patients: A systematic review and analysis of the literature. Intensive Care Med. 2008, 34, 1980–1990. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology 2013, 145, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Millard, A.L.; Mertes, P.M.; Ittelet, D.; Villard, F.; Jeannesson, P.; Bernard, J. Butyrate affects differentiation, maturation and function of human monocyte-derived dendritic cells and macrophages. Clin. Exp. Immunol. 2002, 130, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Byndloss, M.X.; Olsan, E.E.; Rivera-Chávez, F.; Tiffany, C.R.; Cevallos, S.A.; Lokken, K.L.; Torres, T.P.; Byndloss, A.J.; Faber, F.; Gao, Y.; et al. Microbiota-activated PPAR-c signaling inhibits dysbiotic Enterobacteriaceae expansion. Science 2017, 357, 570–575. [Google Scholar] [CrossRef]
- Bae, M.; Kim, H. Mini-Review on the Roles of Vitamin C, Vitamin D, and Selenium in the Immune System against COVID-19. Molecules 2020, 25, 5346. [Google Scholar] [CrossRef] [PubMed]
- Alizadehsani, R.; Sani, Z.A.; Behjati, M.; Roshanzamir, Z.; Hussain, S.; Abedini, N.; Hasanzadeh, F.; Khosravi, A.; Shoeibi, A.; Roshanzamir, M.; et al. Risk factors prediction, clinical outcomes, and mortality in COVID-19 patients. J. Med. Virol. 2021, 93, 2307–2320. [Google Scholar] [CrossRef]
- Ahmadi, S.; Firoozi, D.; Dehghani, M.; Zare, M.; Mehrabi, Z.; Ghaseminasab-Parizi, M.; Masoumi, S.J. Evaluation of Nutritional Status of Intensive Care Unit COVID-19 Patients Based on the Nutritional Risk Screening 2002 Score. Int. J. Clin. Pract. 2022, 2022, 2448161. [Google Scholar] [CrossRef]
- Ting, T.H.Y.; Lo, T.H.M.; Lo, W.W.T.; Ding, Q.; Yuk, D.K.L.; Hui, E.; Tang, M.W.S. Inadequate energy and protein intake; underweight and malnutrition are associated with in-hospital mortality among COVID-19 rehabilitation patients during the omicron outbreak in Hong Kong. Aging Med. 2022, 5, 204–210. [Google Scholar] [CrossRef]
- Yu, L.; Abd Ghani, M.K.; Aghemo, A.; Barh, D.; Bassetti, M.; Catena, F.; Gallo, G.; Gholamrezanezhad, A.; Kamal, M.A.; Lal, A.; et al. SARS-CoV-2 Infection, Inflammation, Immunonutrition, and Pathogenesis of COVID-19. Curr. Med. Chem. 2023, 30, 4390–4408. [Google Scholar] [CrossRef]
- Pimentel, R.F.W.; Silva, A.P.; Santana, A.I.C.; Silva, D.S.E.; Ramos, M.S.; Souza, M.C.; Suen, V.M.M.; Maduro, I.P.N.N.; Ribas Filho, D.; D’Oliveira Júnior, A.; et al. Effect of immuno-nutrition on serum levels of C-reactive protein and lymphocytes in patients with COVID-19: A ran-domized; controlled; double-blind clinical trial. Nutr. Hos. 2022, 39, 20–26. [Google Scholar]
- Scarcella, M.; Scarpellini, E.; Ascani, A.; Commissari, R.; Scorcella, C.; Zanetti, M.; Parisi, A.; Monti, R.; Milic, N.; Donati, A.; et al. Effect of Whey Proteins on Malnutrition and Extubating Time of Critically Ill COVID-19 Patients. Nutrients 2022, 14, 437. [Google Scholar] [CrossRef] [PubMed]
- Scarcella, M.; Scarpellini, E.; Piergallini, S.; Rinninella, E.; Routhiaux, K.; Rasetti, C.; Abenavoli, L.; De Robertis, E.; Manzi, P.; Commissari, R.; et al. Effect of Immuno-Nutrition on Malnutrition, Inflammatory Response and Clinical Course of Semi-Critically Ill COVID-19 Patients: A Pilot Perspective Study. Nutrients 2023, 15, 1250. [Google Scholar] [CrossRef] [PubMed]


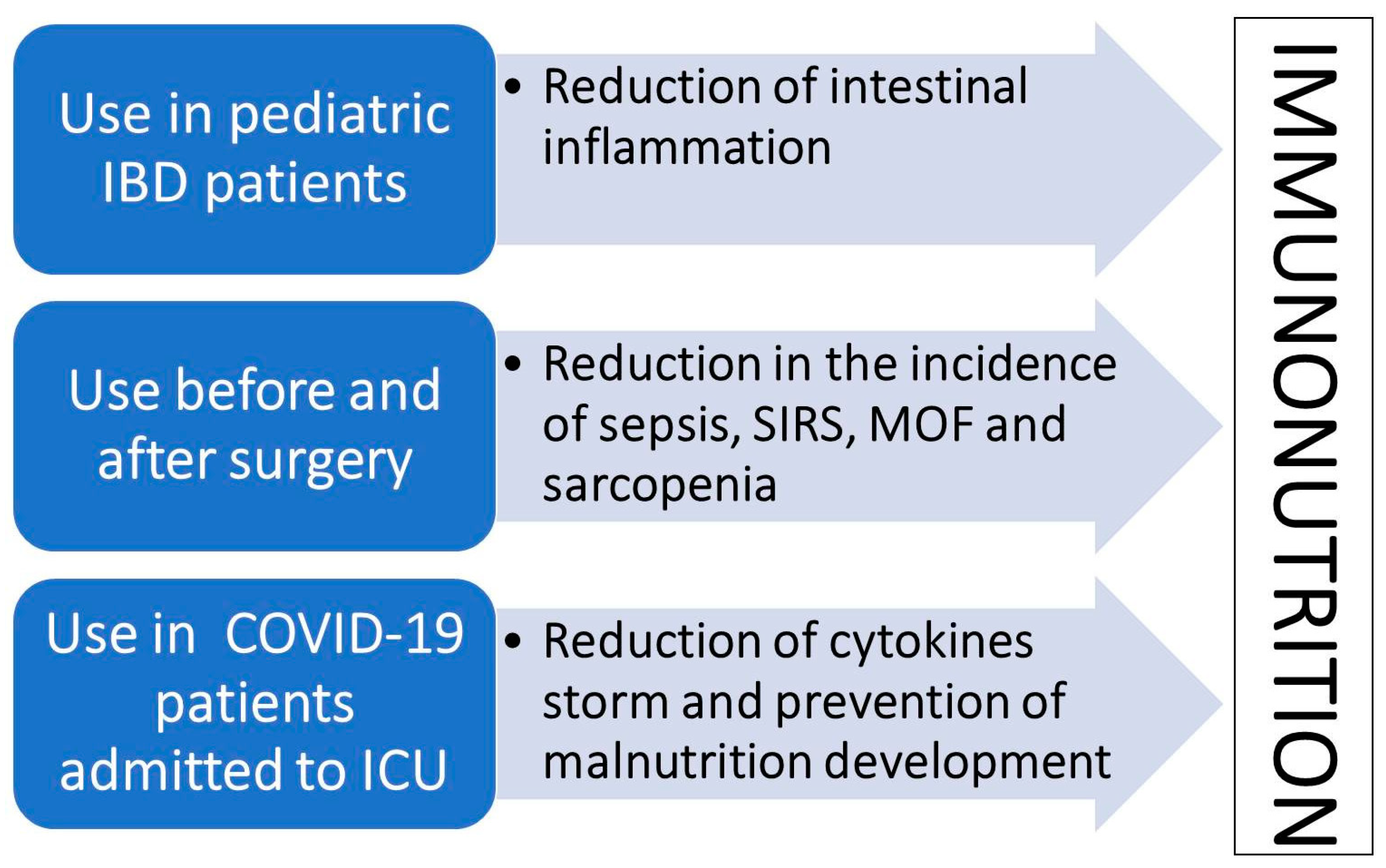{kind=link}
| Non-ICU Patients | ICU Patients |
|---|---|
| Phyla: Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria, Verucomicrobia, Fusobacteria [11,12,13] Genera: Clostridium, Lactobacillus, Enterococcus, Streptococcus, Bacteroides, Prevotella, Pseudomonas, Acinetobacter, Corynebacterium, Actinomyces, Bifidobacterium, Akkermansia, Fusobacterium [11,12,13] | ↑↑ Phyla: Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria [25,26,27] ↑↑ Genera: Clostridium, Lactobacillus, Enterococcus, Streptococcus, Bacteroides, Prevotella, Pseudomonas, Acinetobacter, Corynebacterium, Actinomyces, Bifidobacterium [25,26,27] ↓↓ Phyla: Verucomicrobia, Fusobacteria [25,26,27] ↓↓ Genera: Akkermansia, Fusobacterium, Faecalibacterium [25,26,27] |
| References | Patients | Probiotics | Outcome |
|---|---|---|---|
| Hickson et al. [45] | 135 hospital patients | L. casei, L. bulgaricus, and Streptococcus thermophiles | Reduction in the incidence of diarrhea of 12% vs. 34% from the control group |
| D’Souza et al. [46] | Immunocompromised patients from nine different randomized trials | Saccharomyces boulardii, L. rhamnosus GG, B. longum | Prevention of diarrhea outbreaks |
| Bleichner et al. [47] | 128 ICU patients | S. boulardii | Reduction in the incidence of diarrhea only in the group treated with probiotics |
| Sugawara et al. [53] | 81 patients subjected to hepatectomy | Lactobacillus casei, Bifidobacterium breve | Significantly increase of natural killer cell activity, lymphocyte counts, and reduction of pro-inflammatory cytokines levels after hepatectomy vs. control group |
| Batra et al. [55] | 1103 ICU patients from nine different randomized trials | Lactobacillus spp., Pediococcus spp., Leuconostoc spp., Bifidobacterium spp., Bacillus subtilis, Streptococcus spp., Ergyphilus spp., Bifidus spp., Saccharomyces spp., Enterococcus spp. | Significant reduction of VAP incidence and mortality |
| Ceccarelli G. et al. [57] | 200 patients with severe COVID-19 pneumonia | Streptococcus thermophilus, Bifidobacterium lactis, Lactobacillus acidophilus, Lactobacillus helveticus, Lactobacillus paracasei, Lactobacillus plantarum, and Lactobacillus brevis | Significant association with reduced risk of death |
| Olah et al. [61] | 45 patients with acute pancreatitis | Lactobacillus plantarum | Significant reduction of infectious complications |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abenavoli, L.; Scarpellini, E.; Paravati, M.R.; Scarlata, G.G.M.; Boccuto, L.; Tilocca, B.; Roncada, P.; Luzza, F. Gut Microbiota and Critically Ill Patients: Immunity and Its Modulation via Probiotics and Immunonutrition. Nutrients 2023, 15, 3569. https://doi.org/10.3390/nu15163569
Abenavoli L, Scarpellini E, Paravati MR, Scarlata GGM, Boccuto L, Tilocca B, Roncada P, Luzza F. Gut Microbiota and Critically Ill Patients: Immunity and Its Modulation via Probiotics and Immunonutrition. Nutrients. 2023; 15(16):3569. https://doi.org/10.3390/nu15163569
Chicago/Turabian StyleAbenavoli, Ludovico, Emidio Scarpellini, Maria Rosaria Paravati, Giuseppe Guido Maria Scarlata, Luigi Boccuto, Bruno Tilocca, Paola Roncada, and Francesco Luzza. 2023. "Gut Microbiota and Critically Ill Patients: Immunity and Its Modulation via Probiotics and Immunonutrition" Nutrients 15, no. 16: 3569. https://doi.org/10.3390/nu15163569
APA StyleAbenavoli, L., Scarpellini, E., Paravati, M. R., Scarlata, G. G. M., Boccuto, L., Tilocca, B., Roncada, P., & Luzza, F. (2023). Gut Microbiota and Critically Ill Patients: Immunity and Its Modulation via Probiotics and Immunonutrition. Nutrients, 15(16), 3569. https://doi.org/10.3390/nu15163569