
Effect of Ornithine α-Ketoglutarate on Intestinal Microbiota and Serum Inflammatory Cytokines in Dextran Sulfate Sodium Induced Colitis

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. OKG Treatment
2.3. OKG Treatment in DSS-Induced Colitis
2.4. Serum Biochemical Parameters
2.5. Amino Acids
2.6. Clinical Disease Activity Index
2.7. Colonic HE Staining
2.8. Inflammatory Cytokines in Serum
2.9. Bacterial Profiling
2.10. Statistical Analysis
3. Results
3.1. OKG Treatment Improves Growth Performance in Healthy Mice
3.2. OKG Ameliorates the Body Weight, Colon Length, and DAI of DSS-Induced Colitis in Mice
3.3. OKG Alleviates the Pathological Changes in DSS-Induced Colitis
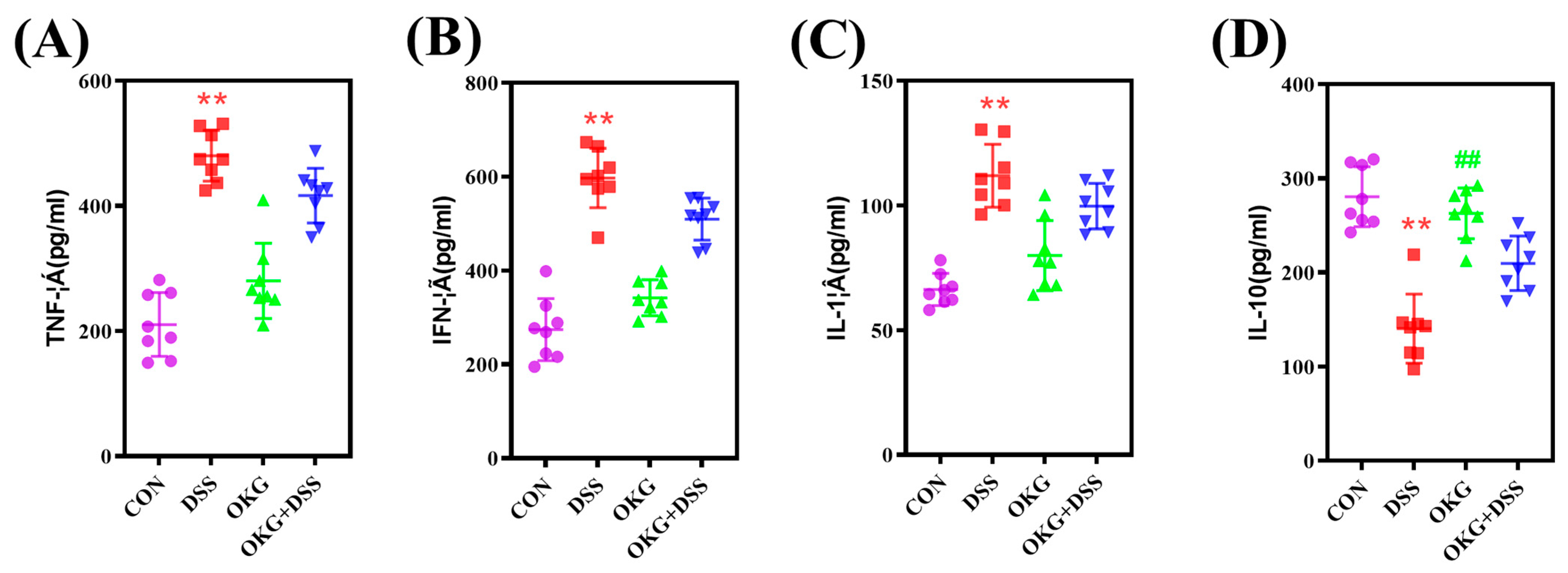
3.4. OKG Affects the Inflammatory Cytokines in DSS-Induced Colitis
3.5. OKG Affects the Gut Microbiota of Subjects with DSS-Induced Colitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chu, H.; Tao, X.; Sun, Z.; Hao, W.; Wei, X. Galactooligosaccharides protects against DSS-induced murine colitis through regulating intestinal flora and inhibiting NF-kappaB pathway. Life Sci. 2020, 242, 117220. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Anwaar, S.; Li, L.; Yin, Z.; Ye, Z.; Huang, Z. A new target for the treatment of inflammatory bowel disease: Interleukin-37. Int. Immunopharmacol. 2020, 83, 106391. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Yu, S.; Shi, C.; Gu, J.; Shao, Y.; Chen, Q.; Li, Y.; Mezzenga, R. Amyloid-Polyphenol Hybrid Nanofilaments Mitigate Colitis and Regulate Gut Microbial Dysbiosis. ACS Nano 2020, 14, 2760–2776. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Wang, K.; Li, Z.; Tao, F.; Xu, Y.; Lan, J.; Chen, G.; Yang, C. Bacillus amyloliquefaciens Ameliorates Dextran Sulfate Sodium-Induced Colitis by Improving Gut Microbial Dysbiosis in Mice Model. Front. Microbiol. 2018, 9, 3260. [Google Scholar] [CrossRef]
- Hernandez-Chirlaque, C.; Aranda, C.J.; Ocon, B.; Capitan-Canadas, F.; Ortega-Gonzalez, M.; Carrero, J.J.; Suarez, M.D.; Zarzuelo, A.; Sanchez de Medina, F.; Martinez-Augustin, O. Germ-free and Antibiotic-treated Mice are Highly Susceptible to Epithelial Injury in DSS Colitis. J. Crohn’s Colitis 2016, 10, 1324–1335. [Google Scholar] [CrossRef]
- Mahajan, R.; Singh, A.; Kedia, S.; Kaur, K.; Midha, V.; Sahu, P.; Mehta, V.; Singh, D.; Bansal, N.; Dharni, K. Maintaining infliximab induced clinical remission with azathioprine and 5-aminosalicylates in acute severe steroid-refractory ulcerative colitis has lower cost and high efficacy (MIRACLE): A multicenter study. Intest. Res. 2022, 20, 64. [Google Scholar] [CrossRef]
- Kane, W.J.; Berry, P.S. Perioperative Assessment and Optimization in Major Colorectal Surgery: Medication Management. Clin. Colon Rectal Surg. 2023, 36, 210–217. [Google Scholar] [CrossRef]
- Feng, H.; Li, Z.; Qi, C.; Wang, X.; Qiao, B.; Wang, K.; Tang, X.; Liu, A. Effect of In Ovo Ornithine-á-Ketoglutarate Feeding on Early Body Weight and Pectoral Muscle Development of Chicks. Indian. J. Anim. Res. 2019, 53, 1679–1683. [Google Scholar] [CrossRef]
- Moinard, C.; Chauveau, B.; Walrand, S.; Felgines, C.; Chassagne, J.; Caldefie, F.; Cynober, L.A.; Vasson, M.-P. Phagocyte functions in stressed rats: Comparison of modulation by glutamine, arginine and ornithine 2-oxoglutarate. Clin. Sci. 1999, 97, 59. [Google Scholar] [CrossRef]
- Schuster, H.; Blanc, M.C.; Genthon, C.; Therond, P.; Bonnefont-Rousselot, D.; Le Tourneau, A.; De Bandt, J.P.; Cynober, L. Does dietary ornithine alpha-ketoglutarate supplementation protect the liver against ischemia-reperfusion injury? Clin. Nutr. 2005, 24, 375–384. [Google Scholar] [CrossRef]
- Jeevanandam, M.; Petersen, S.R. Substrate fuel kinetics in enterally fed trauma patients supplemented with ornithine alpha ketoglutarate. Clin. Nutr. 1999, 18, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, E.S.; Rabelo, C.M.; Prado Neto, A.X.; Garcia, J.H.; Guimaraes, S.B.; Vasconcelos, P.R. Effect of short-term ornithine alpha-ketoglutarate pretreatment on intestinal ischemia-reperfusion in rats. Acta Cir. Bras. 2011, 26 (Suppl. S1), 2–7. [Google Scholar] [CrossRef]
- Canbay, A.; Sowa, J.P. L-Ornithine L-Aspartate (LOLA) as a Novel Approach for Therapy of Non-alcoholic Fatty Liver Disease. Drugs 2019, 79, 39–44. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, Y.; Li, J.; Qi, M.; Tan, B. Serum biochemical parameters and amino acids metabolism are altered in piglets by early-weaning and proline and putrescine supplementations. Anim. Nutr. 2021, 7, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Pruss, K.M.; Enam, F.; Battaglioli, E.; DeFeo, M.; Diaz, O.R.; Higginbottom, S.K.; Fischer, C.R.; Hryckowian, A.J.; Van Treuren, W.; Dodd, D. Oxidative ornithine metabolism supports non-inflammatory C. difficile colonization. Nat. Metab. 2022, 4, 19–28. [Google Scholar] [CrossRef]
- He, L.; Huang, N.; Li, H.; Tian, J.; Zhou, X.; Li, T.; Yao, K.; Wu, G.; Yin, Y. AMPK/α-ketoglutarate axis regulates intestinal water and ion homeostasis in young pigs. J. Agric. Food Chem. 2017, 65, 2287–2298. [Google Scholar] [CrossRef]
- Li, Y.; Wang, P.; Yin, J.; Jin, S.; Su, W.; Tian, J.; Li, T.; Yao, K. Effects of ornithine α-ketoglutarate on growth performance and gut microbiota in a chronic oxidative stress pig model induced by D-galactose. Food Funct. 2020, 11, 472–482. [Google Scholar] [CrossRef]
- Li, Y.; Bao, X.; Yang, F.; Tian, J.; Su, W.; Yin, J.; Yao, K.; Li, T.; Yin, Y. Ornithine α-Ketoglutarate Alleviates Inflammation via Regulating Ileal Mucosa Microbiota and Metabolites in Enterotoxigenic Escherichia coli-Infected Pigs. Front. Nutr. 2022, 9, 862498. [Google Scholar] [CrossRef] [PubMed]
- Guzik, A.C.; Southern, L.L.; Matthews, J.O.; Bidner, T.D.; Ladner, J.P. Ornithine alpha-ketoglutarate and creatine effects on growth and plasma metabolites of nursery pigs. J. Anim. Sci. 2000, 78, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Han, H.; Chen, S.; Gao, J.; Liu, G.; Wu, X.; Deng, J.P.; Yu, Q.F.; Huang, X.G.; et al. Melatonin reprogramming of gut microbiota improves lipid dysmetabolism in high-fat diet-fed mice. J. Pineal Res. 2018, 65, e12524. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.; Yin, Y.; Zhang, S.; Li, X.; Sun, Q.; Wan, D. Effects of Zinc Oxide/Zeolite on Intestinal Morphology, Intestinal Microflora, and Diarrhea Rates in Weaned Piglets. Biol. Trace Element Res. 2020, 199, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, H.; Yin, J.; Zheng, J.; Zhu, X.; Li, T.; Yin, Y. Effects of glutamate and aspartate on growth performance, serum amino acids, and amino acid transporters in piglets. Food Agric. Immunol. 2018, 29, 675–687. [Google Scholar] [CrossRef]
- Yin, J.; Han, H.; Li, Y.; Liu, Z.; Zhao, Y.; Fang, R.; Huang, X.; Zheng, J.; Ren, W.; Wu, F. Lysine Restriction Affects Feed Intake and Amino Acid Metabolism via Gut Microbiome in Piglets. Cell. Physiol. Biochem. 2017, 44, 1749. [Google Scholar] [CrossRef] [PubMed]
- Ghattamaneni, N.K.R.; Panchal, S.K.; Brown, L. An improved rat model for chronic inflammatory bowel disease. Pharmacol. Rep. 2019, 71, 149–155. [Google Scholar] [CrossRef]
- Ding, S.; Ma, Y.; Liu, G.; Yan, W.; Jiang, H.; Fang, J. Lactobacillus brevis Alleviates DSS-Induced Colitis by Reprograming Intestinal Microbiota and Influencing Serum Metabolome in Murine Model. Front. Physiol. 2019, 10, 1152. [Google Scholar] [CrossRef]
- Peng, K.Y.; Gu, J.F.; Su, S.L.; Zhu, Y.; Guo, J.M.; Qian, D.W.; Duan, J.A. Salvia miltiorrhiza stems and leaves total phenolic acids combination with tanshinone protect against DSS-induced ulcerative colitis through inhibiting TLR4/PI3K/AKT/mTOR signaling pathway in mice. J. Ethnopharmacol. 2020, 264, 113052. [Google Scholar] [CrossRef]
- Yin, J.; Li, Y.; Han, H.; Liu, Z.J.; Zeng, X.F.; Li, T.; Yin, Y. Long-Term Effects of Lysine Concentration on Growth Performance, Intestinal Microbiome, and Metabolic Profiles in a Pig model. Food Funct. 2018, 9, 4153–4163. [Google Scholar] [CrossRef]
- Li, Y.; Han, H.; Yin, J.; He, X.; Tang, Z.; Li, T.; Yao, K.; Yin, Y. D- and L-Aspartate regulates growth performance, inflammation and intestinal microbial community in young pigs. Food Funct. 2019, 10, 1028–1037. [Google Scholar] [CrossRef]
- Manuel, M. A new semi-subterranean diving beetle of the Hydroporus normandi-complex from south-eastern France, with notes on other taxa of the complex (Coleoptera: Dytiscidae). Zootaxa 2013, 3652, 453–474. [Google Scholar] [CrossRef]
- Allouchi, H.; Ceolin, R.; Berthon, L.; Tombret, F.; Rietveld, I.B. Characterization of molecular associations involving L-ornithine and alpha-ketoglutaric acid: Crystal structure of L-ornithinium alpha-ketoglutarate. Ann. Pharm. Fr. 2014, 72, 238–243. [Google Scholar] [CrossRef]
- Pernet, P.; Coudray-Lucas, C.; Schneid, C.; Jardel, A.; Cynober, L. Dose dependency of the effect of ornithine α-ketoglutarate on tissue glutamine concentrations and hypercatabolic response in endotoxaemic rats. Br. J. Nutr. 2007, 92, 627. [Google Scholar] [CrossRef] [PubMed]
- Albaugh, V.L.; Mukherjee, K.; Barbul, A. Proline Precursors and Collagen Synthesis: Biochemical Challenges of Nutrient Supplementation and Wound Healing. J. Nutr. 2017, 147, 2011–2017. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes Filho, A.; Cunha, R.M.; Vasconcelos, P.R.; Guimaraes, S.B. Glutamine and ornithine alpha-ketoglutarate supplementation on malate dehydrogenases expression in hepatectomized rats. Acta Cir. Bras. 2014, 29, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Cynober, L.; Lasnier, E.; Le Boucher, J.; Jardel, A.; Coudray-Lucas, C. Effect of ornithine alpha-ketoglutarate on glutamine pools in burn injury: Evidence of component interaction. Intensive Care Med. 2007, 33, 538–541. [Google Scholar] [CrossRef] [PubMed]
- Jeevanandam, M.; Holaday, N.J.; Petersen, S.R. Ornithine-alpha-ketoglutarate (OKG) supplementation is more effective than its component salts in traumatized rats. J. Nutr. 1996, 126, 2141–2150. [Google Scholar] [CrossRef]
- Schneid, C.; De Bandt, J.P.; Cynober, L.; Torres, E.; Reach, G.; Darquy, S. In vivo induction of insulin secretion by ornithine alpha-ketoglutarate: Involvement of nitric oxide and glutamine. Metabolism 2003, 52, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Soendergaard, C.; Young, J.A.; Kopchick, J.J. Growth Hormone Resistance-Special Focus on Inflammatory Bowel Disease. Int. J. Mol. Sci. 2017, 18, 1019. [Google Scholar] [CrossRef]
- Loi, C.; Nakib, S.; Neveux, N.; Arnaud-Battandier, F.; Cynober, L. Ornithine alpha-ketoglutarate metabolism in the healthy rat in the postabsorptive state. Metabolism 2005, 54, 1108–1114. [Google Scholar] [CrossRef]
- Rauch, F.; Middelmann, B.; Cagnoli, M.; Keller, K.M.; Schönau, E. Comparison of total alkaline phosphatase and three assays for bone-specific alkaline phosphatase in childhood and adolescence. Acta Paediatr. 2010, 86, 583–587. [Google Scholar] [CrossRef]
- Prvulovic, D.; Kosarcic, S.; Popovic, M.; Dimitrijevic, D.; Grubor-Lajsic, G. The Influence of Hydrated Aluminosilicate on Biochemical and Haematological Blood Parameters, Growth Performance and Carcass Traits of Pigs. J. Anim. Vet. Adv. 2012, 11, 134–140. [Google Scholar] [CrossRef]
- Mirabile, A.; Rivoltini, L.; Daveri, E.; Vernieri, C.; Mele, R.; Porcu, L.; Lazzari, C.; Bulotta, A.; Vigano, M.G.; Cascinu, S.; et al. Metabolism and Immune Modulation in Patients with Solid Tumors: Systematic Review of Preclinical and Clinical Evidence. Cancers 2020, 12, 1153. [Google Scholar] [CrossRef] [PubMed]
- Tatara, M.R.; Sliwa, E.; Krupski, W.; Brodzki, A.; Pasternak, K. Ornithine alpha-ketoglutarate increases mineralization and mechanical properties of tibia in turkeys. Bone 2006, 39, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Donsbough, A.L.; Powell, S.; Waguespack, A.; Bidner, T.D.; Southern, L.L. Uric acid, urea, and ammonia concentrations in serum and uric acid concentration in excreta as indicators of amino acid utilization in diets for broilers. Poult. Sci. 2010, 89, 287–294. [Google Scholar] [CrossRef]
- Lopez-Colom, P.; Castillejos, L.; Barba-Vidal, E.; Zhu, Y.; Puyalto, M.; Jose Mallo, J.; Martin-Orue, S.M. Response of gastrointestinal fermentative activity and colonic microbiota to protected sodium butyrate and protected sodium heptanoate in weaned piglets challenged with ETEC F4(+). Arch. Anim. Nutr. 2019, 73, 339–359. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; He, Z.; Zhou, Y.; Ross, R.P.; Stanton, C.; Zhao, J.; Zhang, H.; Yang, B.; Chen, W. Lactobacillus plantarum relieves diarrhea caused by enterotoxin-producing Escherichia coli through inflammation modulation and gut microbiota regulation. Food Funct. 2020, 11, 10362–10374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Jiao, H.; Wang, C.; Lin, Y.; You, S. Chlorogenic Acid Ameliorates Colitis and Alters Colonic Microbiota in a Mouse Model of Dextran Sulfate Sodium-Induced Colitis. Front. Physiol. 2019, 10, 325. [Google Scholar] [CrossRef]
- Le Bricon, T.; Cynober, L.; Baracos, V. Ornithine α-ketoglutarate limits muscle protein breakdown without stimulating tumor growth in rats bearing Yoshida ascites hepatoma. Metabolism 1994, 43, 899–905. [Google Scholar] [CrossRef]
- Zhai, Z.; Zhang, F.; Cao, R.; Ni, X.; Xin, Z.; Deng, J.; Wu, G.; Ren, W.; Yin, Y.; Deng, B. Cecropin A Alleviates Inflammation Through Modulating the Gut Microbiota of C57BL/6 Mice With DSS-Induced IBD. Front. Microbiol. 2019, 10, 1595. [Google Scholar] [CrossRef]
- Sangaraju, R.; Nalban, N.; Alavala, S.; Rajendran, V.; Jerald, M.K.; Sistla, R. Protective effect of galangin against dextran sulfate sodium (DSS)-induced ulcerative colitis in Balb/c mice. Inflamm. Res. 2019, 68, 691–704. [Google Scholar] [CrossRef]
- Vallabhapurapu, S.; Karin, M. Regulation and Function of NF-kappa B Transcription Factors in the Immune System. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef]
- Stecher, B.; Hardt, W.D. The role of microbiota in infectious disease. Trends Microbiol. 2008, 16, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Li, H.H.; Li, Y.P.; Zhu, Q.; Qiao, J.Y.; Wang, W.J. Dietary supplementation with Clostridium butyricum helps to improve the intestinal barrier function of weaned piglets challenged with enterotoxigenic Escherichia coli K88. J. Appl. Microbiol. 2018, 125, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Li, Y.; Fang, J.; Liu, G.; Yin, J.; Li, T.; Yin, Y. Gut Microbiota and Type 1 Diabetes. Int. J. Mol. Sci. 2018, 19, 995. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Luo, J.; Zhang, C.; Liu, L.; Ou, S.; Zhang, G.; Peng, X. Alliin protects against inflammatory bowel disease by preserving the gene expression in colonic epithelial cells rather than altering gut microbiota. J. Funct. Foods 2019, 59, 309–318. [Google Scholar] [CrossRef]
- Schlegel, L.; Coudray-Lucas, C.; Barbut, F.d.r.; Le Boucher, J.; Jardel, A.; Zarrabian, S.; Cynober, L. Bacterial dissemination and metabolic changes in rats induced by endotoxemia following intestinal E. coli overgrowth are reduced by ornithine α-ketoglutarate administration. J. Nutr. 2000, 130, 2897–2902. [Google Scholar] [CrossRef]
- Moayyedi, P.; Surette, M.G.; Kim, P.T.; Libertucci, J.; Wolfe, M.; Onischi, C.; Armstrong, D.; Marshall, J.K.; Kassam, Z.; Reinisch, W.; et al. Fecal Microbiota Transplantation Induces Remission in Patients With Active Ulcerative Colitis in a Randomized Controlled Trial. Gastroenterology 2015, 149, 102–109.e106. [Google Scholar] [CrossRef]
- Goethel, A.; Croitoru, K.; Philpott, D.J. The interplay between microbes and the immune response in inflammatory bowel disease. J. Physiol. 2018, 596, 3869–3882. [Google Scholar] [CrossRef]
- Mizoguchi, E.; Dziarski, R.; Park, S.Y.; Kashyap, D.R.; Dowd, S.E.; Gupta, D. Pglyrp-Regulated Gut Microflora Prevotella falsenii, Parabacteroides distasonis and Bacteroides eggerthii Enhance and Alistipes finegoldii Attenuates Colitis in Mice. PLoS ONE 2016, 11, e0146162. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature Graded | Description | Grade |
|---|---|---|
| Inflammation | None | 0 |
| Slight | 1 | |
| Moderate | 2 | |
| Severe | 3 | |
| Extent | None | 0 |
| Mucosa | 1 | |
| Submucosa | 2 | |
| Transmural | 3 | |
| Percentage of involvement (%) | 1–25 | 1 |
| 26–50 | 2 | |
| 51–75 | 3 | |
| 76–100 | 4 | |
| Crypt damage | None | 0 |
| Submucosa | 1 | |
| Basal one-third lost | 2 | |
| Basal two-thirds lost | 3 | |
| Only surface epithelium intact | 4 | |
| All crypts and epithelia are destroyed | 5 | |
| Ulceration | No ulceration | 0 |
| Mild ulceration | 1 | |
| Moderate ulceration | 2 | |
| Extensive ulceration | 3 |
| Item | Control | LOKG | MOKG | HOKG | p-Value |
|---|---|---|---|---|---|
| TP | 27.85 ± 0.90 | 28.31 ± 0.51 | 27.84 ± 0.61 | 29.05 ± 1.12 | 0.699 |
| ALB | 18.45 ± 0.70 | 18.75 ± 0.37 | 18.10 ± 0.48 | 18.73 ± 0.83 | 0.871 |
| BUN | 3.36 ± 0.09 a | 2.81 ± 0.18 b | 2.77 ± 0.09 b | 3.53 ± 0.14 a | 0.000 |
| UA | 1.17 ± 0.04 b | 1.23 ± 0.04 ab | 1.30 ± 0.05 ab | 1.43 ± 0.09 a | 0.048 |
| GLU | 1.38 ± 0.10 b | 1.55 ± 0.09 b | 2.32 ± 0.19 a | 2.10 ± 0.13 a | 0.000 |
| NH3L | 273.39 ± 9.44 ab | 259.88 ± 9.01 ab | 243.01 ± 4.83 b | 282.20 ± 16.2 a | 0.041 |
| ALT | 25.45 ± 1.88 a | 25.23 ± 1.25 a | 23.73 ± 0.83 a | 16.56 ± 1.47 b | 0.000 |
| AST | 254.40 ± 13.85 a | 198.67 ± 14.17 ab | 281.40 ± 18.59 a | 64.80 ± 8.91 b | 0.000 |
| ALP | 48.50 ± 3.01 b | 59.60 ± 3.67 a | 64.63 ± 2.85 a | 54.00 ± 0.26 ab | 0.002 |
| Item | Control | LOKG | MOKG | HOKG | p-Value |
|---|---|---|---|---|---|
| Asp | 7.21 ± 0.27 | 7.15 ± 0.18 | 7.47 ± 0.22 | 7.62 ± 0.30 | 0.509 |
| Thr | 21.02 ± 1.33 | 20.78 ± 0.39 | 21.27 ± 0.44 | 20.22 ± 1.04 | 0.874 |
| Ser | 9.54 ± 0.46 | 9.19 ± 0.24 | 8.92 ± 0.41 | 9.48 ± 0.63 | 0.758 |
| Glu | 20.10 ± 0.88 | 19.02 ± 0.48 | 18.88 ± 0.67 | 21.81 ± 1.64 | 0.159 |
| Gly | 8.94 ± 0.27 | 9.34 ± 0.29 | 9.33 ± 0.21 | 8.88 ± 0.48 | 0.663 |
| Ala | 15.29 ± 1.17 | 13.74 ± 0.49 | 14.15 ± 0.81 | 13.70 ± 0.43 | 0.470 |
| Cys | 12.17 ± 0.70 b | 12.66 ± 0.37 b | 13.23 ± 0.20 b | 16.84 ± 0.37 a | 0.000 |
| Val | 9.87 ± 0.95 | 8.09 ± 0.35 | 8.08 ± 0.05 | 9.56 ± 1.12 | 0.271 |
| Met | 9.52 ± 0.79 | 8.04 ± 0.22 | 9.23 ± 0.35 | 9.63 ± 0.45 | 0.159 |
| Ile | 3.36 ± 0.32 a | 2.37 ± 0.05 b | 2.39 ± 0.12 b | 2.96 ± 0.31 ab | 0.028 |
| Leu | 6.27 ± 0.46 | 5.25 ± 0.12 | 7.88 ± 1.86 | 6.86 ± 1.27 | 0.427 |
| Tyr | 4.03 ± 0.67 ab | 2.89 ± 0.26 b | 7.28 ± 1.71 a | 4.42 ± 1.48 ab | 0.039 |
| Phe | 3.41 ± 0.16 b | 4.39 ± 0.32 b | 7.31 ± 0.62 a | 6.84 ± 0.63 a | 0.000 |
| Lys | 25.53 ± 1.43 | 26.72 ± 0.81 | 27.03 ± 1.84 | 26.76 ± 1.42 | 0.889 |
| His | 4.59 ± 0.76 b | 4.86 ± 0.66 b | 6.15 ± 0.99 ab | 7.70 ± 1.17 a | 0.046 |
| Arg | 10.82 ± 0.74 | 10.42 ± 0.38 | 10.40 ± 0.37 | 11.73 ± 0.70 | 0.365 |
| Pro | 12.69 ± 0.67 | 10.54 ± 0.45 | 12.17 ± 0.90 | 10.53 ± 0.86 | 0.095 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Tian, J.; Su, W.; Yang, F.; Yin, J.; Jiang, Q.; Li, Y.; Yao, K.; Li, T.; Yin, Y. Effect of Ornithine α-Ketoglutarate on Intestinal Microbiota and Serum Inflammatory Cytokines in Dextran Sulfate Sodium Induced Colitis. Nutrients 2023, 15, 2476. https://doi.org/10.3390/nu15112476
Wang T, Tian J, Su W, Yang F, Yin J, Jiang Q, Li Y, Yao K, Li T, Yin Y. Effect of Ornithine α-Ketoglutarate on Intestinal Microbiota and Serum Inflammatory Cytokines in Dextran Sulfate Sodium Induced Colitis. Nutrients. 2023; 15(11):2476. https://doi.org/10.3390/nu15112476
Chicago/Turabian StyleWang, Tao, Junquan Tian, Wenxuan Su, Fan Yang, Jie Yin, Qian Jiang, Yuying Li, Kang Yao, Tiejun Li, and Yulong Yin. 2023. "Effect of Ornithine α-Ketoglutarate on Intestinal Microbiota and Serum Inflammatory Cytokines in Dextran Sulfate Sodium Induced Colitis" Nutrients 15, no. 11: 2476. https://doi.org/10.3390/nu15112476
APA StyleWang, T., Tian, J., Su, W., Yang, F., Yin, J., Jiang, Q., Li, Y., Yao, K., Li, T., & Yin, Y. (2023). Effect of Ornithine α-Ketoglutarate on Intestinal Microbiota and Serum Inflammatory Cytokines in Dextran Sulfate Sodium Induced Colitis. Nutrients, 15(11), 2476. https://doi.org/10.3390/nu15112476