
Association of NAFLD with FGF21 Polygenic Hazard Score, and Its Interaction with Protein Intake Level in Korean Adults


Abstract
1. Introduction
2. Materials and Methods
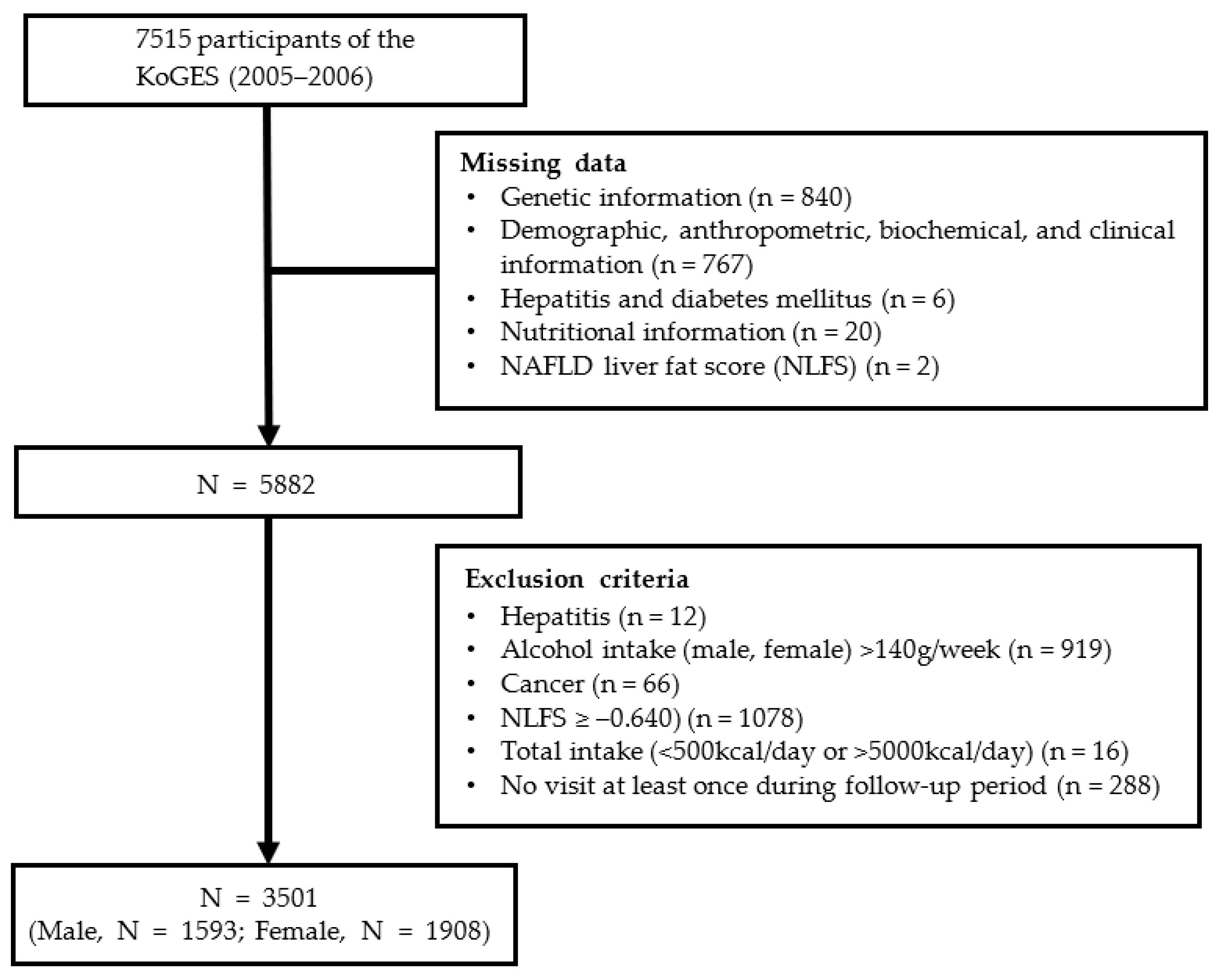
2.1. Study Participants
2.2. Demographic, Anthropometric, and Biochemical Data of the Study Population
2.3. Dietary Assessment
2.4. NAFLD Diagnosis Using the NLFS
2.5. Quality Control, Genotyping, and Genetic-Variant Selection
2.6. PHS Development and Calculation for NAFLD
2.7. Expression Quantitative Trait Loci (eQTL) Analysis of Eight SNPs Using Genotype-Tissue Expression (GTEx)
2.8. Statistical Analysis
3. Results
3.1. Baseline Characteristics and Nutritional Intake
3.2. SNP Selection for PHS
3.3. Association between the PHS and NAFLD Incidence
3.4. Association between the PHS and NAFLD Incidence by Protein Intake
3.5. Potential Effects of Genetic Variants on Gene Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Am. J. Gastroenterol. 2012, 107, 811–826. [Google Scholar] [CrossRef] [PubMed]
- Le, M.H.; Yeo, Y.H.; Li, X.; Li, J.; Zou, B.; Wu, Y.; Ye, Q.; Huang, D.Q.; Zhao, C.; Zhang, J.; et al. 2019 Global NAFLD Prevalence: A Systematic Review and Meta-analysis. Clin. Gastroenterol. Hepatol. 2022, 20, 2809–2817.e28. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Friedman, S.L.; Shulman, G.I. Mechanisms and disease consequences of nonalcoholic fatty liver disease. Cell 2021, 184, 2537–2564. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Effenberger, M. From NAFLD to MAFLD: When pathophysiology succeeds. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 387–388. [Google Scholar] [CrossRef]
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wai-Sun Wong, V.; Dufour, J.F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef]
- Ye, Q.; Zou, B.; Yeo, Y.H.; Li, J.; Huang, D.Q.; Wu, Y.; Yang, H.; Liu, C.; Kam, L.Y.; Tan, X.X.E.; et al. Global prevalence, incidence, and outcomes of non-obese or lean non-alcoholic fatty liver disease: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2020, 5, 739–752. [Google Scholar] [CrossRef]
- Eslam, M.; Valenti, L.; Romeo, S. Genetics and epigenetics of NAFLD and NASH: Clinical impact. J. Hepatol. 2018, 68, 268–279. [Google Scholar] [CrossRef]
- Romeo, S.; Kozlitina, J.; Xing, C.; Pertsemlidis, A.; Cox, D.; Pennacchio, L.A.; Boerwinkle, E.; Cohen, J.C.; Hobbs, H.H. Genetic variation in PNPLA3 confers susceptibility to nonalcoholic fatty liver disease. Nat. Genet. 2008, 40, 1461–1465. [Google Scholar] [CrossRef]
- Sliz, E.; Sebert, S.; Würtz, P.; Kangas, A.J.; Soininen, P.; Lehtimäki, T.; Kähönen, M.; Viikari, J.; Männikkö, M.; Ala-Korpela, M.; et al. NAFLD risk alleles in PNPLA3, TM6SF2, GCKR and LYPLAL1 show divergent metabolic effects. Hum. Mol. Genet. 2018, 27, 2214–2223. [Google Scholar] [CrossRef]
- Kozlitina, J. Genetic Risk Factors and Disease Modifiers of Nonalcoholic Steatohepatitis. Gastroenterol. Clin. N. Am. 2020, 49, 25–44. [Google Scholar] [CrossRef]
- Trépo, E.; Valenti, L. Update on NAFLD genetics: From new variants to the clinic. J. Hepatol. 2020, 72, 1196–1209. [Google Scholar] [CrossRef] [PubMed]
- Geng, L.; Lam, K.S.L.; Xu, A. The therapeutic potential of FGF21 in metabolic diseases: From bench to clinic. Nat. Rev. Endocrinol. 2020, 16, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Investig. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, T.; Dutchak, P.; Zhao, G.; Ding, X.; Gautron, L.; Parameswara, V.; Li, Y.; Goetz, R.; Mohammadi, M.; Esser, V.; et al. Endocrine regulation of the fasting response by PPARα-mediated induction of fibroblast growth factor 21. Cell Metab. 2007, 5, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Lee, M.S. FGF21 as a Stress Hormone: The Roles of FGF21 in Stress Adaptation and the Treatment of Metabolic Diseases. Diabetes Metab. J. 2014, 38, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Kurosu, H.; Yamamoto, M.; Nandi, A.; Rosenblatt, K.P.; Goetz, R.; Eliseenkova, A.V.; Mohammadi, M.; Kuro-o, M. BetaKlotho is required for metabolic activity of fibroblast growth factor 21. Proc. Natl. Acad. Sci. USA 2007, 104, 7432–7437. [Google Scholar] [CrossRef]
- Suzuki, M.; Uehara, Y.; Motomura-Matsuzaka, K.; Oki, J.; Koyama, Y.; Kimura, M.; Asada, M.; Komi-Kuramochi, A.; Oka, S.; Imamura, T. βKlotho is required for fibroblast growth factor (FGF) 21 signaling through FGF receptor (FGFR) 1c and FGFR3c. Mol. Endocrinol. 2008, 22, 1006–1014. [Google Scholar] [CrossRef]
- Wu, X.; Ge, H.; Lemon, B.; Vonderfecht, S.; Weiszmann, J.; Hecht, R.; Gupte, J.; Hager, T.; Wang, Z.; Lindberg, R.; et al. FGF19-induced hepatocyte proliferation is mediated through FGFR4 activation. J. Biol. Chem. 2010, 285, 5165–5170. [Google Scholar] [CrossRef]
- Yang, C.; Jin, C.; Li, X.; Wang, F.; McKeehan, W.L.; Luo, Y. Differential specificity of endocrine FGF19 and FGF21 to FGFR1 and FGFR4 in complex with KLB. PLoS ONE 2012, 7, e33870. [Google Scholar] [CrossRef]
- Markan, K.R.; Naber, M.C.; Ameka, M.K.; Anderegg, M.D.; Mangelsdorf, D.J.; Kliewer, S.A.; Mohammadi, M.; Potthoff, M.J. Circulating FGF21 is liver derived and enhances glucose uptake during refeeding and overfeeding. Diabetes 2014, 63, 4057–4063. [Google Scholar] [CrossRef]
- Bartesaghi, S.; Wallenius, K.; Hovdal, D.; Liljeblad, M.; Wallin, S.; Dekker, N.; Barlind, L.; Davies, N.; Seeliger, F.; Winzell, M.S.; et al. Subcutaneous delivery of FGF21 mRNA therapy reverses obesity, insulin resistance, and hepatic steatosis in diet-induced obese mice. Mol. Ther. Nucleic Acids 2022, 28, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Queen, N.J.; Bates, R.; Huang, W.; Xiao, R.; Appana, B.; Cao, L. Visceral adipose tissue-directed FGF21 gene therapy improves metabolic and immune health in BTBR mice. Mol. Ther. Methods Clin. Dev. 2021, 20, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Camporez, J.P.; Jornayvaz, F.R.; Petersen, M.C.; Pesta, D.; Guigni, B.A.; Serr, J.; Zhang, D.; Kahn, M.; Samuel, V.T.; Jurczak, M.J.; et al. Cellular mechanisms by which FGF21 improves insulin sensitivity in male mice. Endocrinology 2013, 154, 3099–3109. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lloyd, D.J.; Hale, C.; Stanislaus, S.; Chen, M.; Sivits, G.; Vonderfecht, S.; Hecht, R.; Li, Y.S.; Lindberg, R.A.; et al. Fibroblast growth factor 21 reverses hepatic steatosis, increases energy expenditure, and improves insulin sensitivity in diet-induced obese mice. Diabetes 2009, 58, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Fang, Q.; Gao, F.; Fan, J.; Zhou, J.; Wang, X.; Zhang, H.; Pan, X.; Bao, Y.; Xiang, K.; et al. Fibroblast growth factor 21 levels are increased in nonalcoholic fatty liver disease patients and are correlated with hepatic triglyceride. J. Hepatol. 2010, 53, 934–940. [Google Scholar] [CrossRef]
- Rusli, F.; Deelen, J.; Andriyani, E.; Boekschoten, M.V.; Lute, C.; van den Akker, E.B.; Müller, M.; Beekman, M.; Steegenga, W.T. Fibroblast growth factor 21 reflects liver fat accumulation and dysregulation of signalling pathways in the liver of C57BL/6J mice. Sci. Rep. 2016, 6, 30484. [Google Scholar] [CrossRef]
- Yilmaz, Y.; Eren, F.; Yonal, O.; Kurt, R.; Aktas, B.; Celikel, C.A.; Ozdogan, O.; Imeryuz, N.; Kalayci, C.; Avsar, E. Increased serum FGF21 levels in patients with nonalcoholic fatty liver disease. Eur. J. Clin. Investig. 2010, 40, 887–892. [Google Scholar] [CrossRef]
- Fisher, F.M.; Chui, P.C.; Antonellis, P.J.; Bina, H.A.; Kharitonenkov, A.; Flier, J.S.; Maratos-Flier, E. Obesity is a fibroblast growth factor 21 (FGF21)-resistant state. Diabetes 2010, 59, 2781–2789. [Google Scholar] [CrossRef]
- Gong, S.G. Isoforms of receptors of fibroblast growth factors. J. Cell. Physiol. 2014, 229, 1887–1895. [Google Scholar] [CrossRef]
- Yan, G.; Fukabori, Y.; McBride, G.; Nikolaropolous, S.; McKeehan, W.L. Exon switching and activation of stromal and embryonic fibroblast growth factor (FGF)-FGF receptor genes in prostate epithelial cells accompany stromal independence and malignancy. Mol. Cell. Biol. 1993, 13, 4513–4522. [Google Scholar] [CrossRef]
- Nygaard, E.B.; Ørskov, C.; Almdal, T.; Vestergaard, H.; Andersen, B. Fasting decreases plasma FGF21 in obese subjects and the expression of FGF21 receptors in adipose tissue in both lean and obese subjects. J. Endocrinol. 2018, 239, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Gälman, C.; Lundåsen, T.; Kharitonenkov, A.; Bina, H.A.; Eriksson, M.; Hafström, I.; Dahlin, M.; Amark, P.; Angelin, B.; Rudling, M. The circulating metabolic regulator FGF21 is induced by prolonged fasting and PPARα activation in man. Cell Metab. 2008, 8, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Dushay, J.; Chui, P.C.; Gopalakrishnan, G.S.; Varela-Rey, M.; Crawley, M.; Fisher, F.M.; Badman, M.K.; Martinez-Chantar, M.L.; Maratos-Flier, E. Increased fibroblast growth factor 21 in obesity and nonalcoholic fatty liver disease. Gastroenterology 2010, 139, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.M.; Laeger, T.; Albarado, D.C.; McDougal, D.H.; Berthoud, H.R.; Münzberg, H.; Morrison, C.D. Low protein-induced increases in FGF21 drive UCP1-dependent metabolic but not thermoregulatory endpoints. Sci. Rep. 2017, 7, 8209. [Google Scholar] [CrossRef]
- Xu, C.; Markova, M.; Seebeck, N.; Loft, A.; Hornemann, S.; Gantert, T.; Kabisch, S.; Herz, K.; Loske, J.; Ost, M.; et al. High-protein diet more effectively reduces hepatic fat than low-protein diet despite lower autophagy and FGF21 levels. Liver Int. 2020, 40, 2982–2997. [Google Scholar] [CrossRef]
- Laeger, T.; Henagan, T.M.; Albarado, D.C.; Redman, L.M.; Bray, G.A.; Noland, R.C.; Münzberg, H.; Hutson, S.M.; Gettys, T.W.; Schwartz, M.W.; et al. FGF21 is an endocrine signal of protein restriction. J. Clin. Investig. 2014, 124, 3913–3922. [Google Scholar] [CrossRef]
- Qian, Z.; Zhang, Y.; Yang, N.; Nie, H.; Yang, Z.; Luo, P.; Wei, X.; Guan, Y.; Huang, Y.; Yan, J.; et al. Close association between lifestyle and circulating FGF21 levels: A systematic review and meta-analysis. Front. Endocrinol. 2022, 13, 984828. [Google Scholar] [CrossRef]
- Green, C.L.; Pak, H.H.; Richardson, N.E.; Flores, V.; Yu, D.; Tomasiewicz, J.L.; Dumas, S.N.; Kredell, K.; Fan, J.W.; Kirsh, C.; et al. Sex and genetic background define the metabolic, physiologic, and molecular response to protein restriction. Cell Metab. 2022, 34, 209–226.e205. [Google Scholar] [CrossRef]
- Maida, A.; Zota, A.; Sjøberg, K.A.; Schumacher, J.; Sijmonsma, T.P.; Pfenninger, A.; Christensen, M.M.; Gantert, T.; Fuhrmeister, J.; Rothermel, U.; et al. A liver stress-endocrine nexus promotes metabolic integrity during dietary protein dilution. J. Clin. Investig. 2016, 126, 3263–3278. [Google Scholar] [CrossRef]
- Wu, Y.; Li, B.; Li, L.; Mitchell, S.E.; Green, C.L.; D’Agostino, G.; Wang, G.; Wang, L.; Li, M.; Li, J.; et al. Very-low-protein diets lead to reduced food intake and weight loss, linked to inhibition of hypothalamic mTOR signaling, in mice. Cell Metab. 2021, 33, 888–904.e886. [Google Scholar] [CrossRef]
- Frayling, T.M.; Beaumont, R.N.; Jones, S.E.; Yaghootkar, H.; Tuke, M.A.; Ruth, K.S.; Casanova, F.; West, B.; Locke, J.; Sharp, S.; et al. A Common Allele in FGF21 Associated with Sugar Intake Is Associated with Body Shape, Lower Total Body-Fat Percentage, and Higher Blood Pressure. Cell Rep. 2018, 23, 327–336. [Google Scholar] [CrossRef]
- Søberg, S.; Sandholt, C.H.; Jespersen, N.Z.; Toft, U.; Madsen, A.L.; von Holstein-Rathlou, S.; Grevengoed, T.J.; Christensen, K.B.; Bredie, W.L.P.; Potthoff, M.J.; et al. FGF21 Is a Sugar-Induced Hormone Associated with Sweet Intake and Preference in Humans. Cell Metab. 2017, 25, 1045–1053.e1046. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, N.; Shi, J.; García-Closas, M. Developing and evaluating polygenic risk prediction models for stratified disease prevention. Nat. Rev. Genet. 2016, 17, 392–406. [Google Scholar] [CrossRef]
- Kim, Y.; Han, B.G. Cohort Profile: The Korean Genome and Epidemiology Study (KoGES) Consortium. Int. J. Epidemiol. 2017, 46, e20. [Google Scholar] [CrossRef]
- Jang, M.; Won, C.; Choi, H.; Kim, S.; Park, W.; Kim, D.; Jeong, S.; Kim, B. Effects of physical activity on fractures in adults: A community-based Korean cohort study. Korean J. Sport. Med. 2017, 35, 97–102. [Google Scholar] [CrossRef]
- Ahn, Y.; Kwon, E.; Shim, J.E.; Park, M.K.; Joo, Y.; Kimm, K.; Park, C.; Kim, D.H. Validation and reproducibility of food frequency questionnaire for Korean genome epidemiologic study. Eur. J. Clin. Nutr. 2007, 61, 1435–1441. [Google Scholar] [CrossRef]
- Korean Nutrition Society. 2020 Dietary Reference Intakes for Koreans: Energy and Macronutrients; Ministry of Health and Welfare: Sejong, Republic of Korea, 2020.
- Kotronen, A.; Peltonen, M.; Hakkarainen, A.; Sevastianova, K.; Bergholm, R.; Johansson, L.M.; Lundbom, N.; Rissanen, A.; Ridderstråle, M.; Groop, L.; et al. Prediction of non-alcoholic fatty liver disease and liver fat using metabolic and genetic factors. Gastroenterology 2009, 137, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.N.; Kawachi, I.; Chang, J.; Boo, K.; Shin, H.G.; Lee, H.; Cho, S.I. Marital status, gender, and depression: Analysis of the baseline survey of the Korean Longitudinal Study of Ageing (KLoSA). Soc. Sci. Med. 2009, 69, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.S.; Go, M.J.; Kim, Y.J.; Heo, J.Y.; Oh, J.H.; Ban, H.J.; Yoon, D.; Lee, M.H.; Kim, D.J.; Park, M.; et al. A large-scale genome-wide association study of Asian populations uncovers genetic factors influencing eight quantitative traits. Nat. Genet. 2009, 41, 527–534. [Google Scholar] [CrossRef]
- Abecasis, G.R.; Auton, A.; Brooks, L.D.; DePristo, M.A.; Durbin, R.M.; Handsaker, R.E.; Kang, H.M.; Marth, G.T.; McVean, G.A. An integrated map of genetic variation from 1,092 human genomes. Nature 2012, 491, 56–65. [Google Scholar] [CrossRef]
- Consortium, G. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, H.S.; Ahn, S.B.; Kwon, Y.J. Dairy protein intake is inversely related to development of non-alcoholic fatty liver disease. Clin. Nutr. 2021, 40, 5252–5260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Meng, G.; Zhang, Q.; Liu, L.; Wu, H.; Gu, Y.; Wang, Y.; Zhang, T.; Wang, X.; Zhang, J.; et al. Inflammatory potential of diet and risk of nonalcoholic fatty liver disease: A prospective cohort study. Eur. J. Clin. Nutr. 2022, 76, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhang, K.; Chen, Y.; Li, Y.; Li, Y.; Fu, K.; Feng, R. Associations between Dietary Nutrient Intakes and Hepatic Lipid Contents in NAFLD Patients Quantified by ¹H-MRS and Dual-Echo MRI. Nutrients 2016, 8, 527. [Google Scholar] [CrossRef]
- Alferink, L.J.; Kiefte-de Jong, J.C.; Erler, N.S.; Veldt, B.J.; Schoufour, J.D.; de Knegt, R.J.; Ikram, M.A.; Metselaar, H.J.; Janssen, H.; Franco, O.H.; et al. Association of dietary macronutrient composition and non-alcoholic fatty liver disease in an ageing population: The Rotterdam Study. Gut 2019, 68, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Kaess, B.M.; Barnes, T.A.; Stark, K.; Charchar, F.J.; Waterworth, D.; Song, K.; Wang, W.Y.; Vollenweider, P.; Waeber, G.; Mooser, V.; et al. FGF21 signalling pathway and metabolic traits—Genetic association analysis. Eur. J. Hum. Genet. 2010, 18, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Liu, Y.; Hao, J.G.; Wang, L.P.; Dai, M.J.; Shen, G.F.; Yan, X.B. KLB gene polymorphism is associated with obesity and non-alcoholic fatty liver disease in the Han Chinese. Aging 2019, 11, 7847–7858. [Google Scholar] [CrossRef]
- Tanaka, T.; Ngwa, J.S.; van Rooij, F.J.; Zillikens, M.C.; Wojczynski, M.K.; Frazier-Wood, A.C.; Houston, D.K.; Kanoni, S.; Lemaitre, R.N.; Luan, J.; et al. Genome-wide meta-analysis of observational studies shows common genetic variants associated with macronutrient intake. Am. J. Clin. Nutr. 2013, 97, 1395–1402. [Google Scholar] [CrossRef]
- Chu, A.Y.; Workalemahu, T.; Paynter, N.P.; Rose, L.M.; Giulianini, F.; Tanaka, T.; Ngwa, J.S.; CHARGE Nutrition Working Group; Qi, Q.; Curhan, G.C.; et al. Novel locus including FGF21 is associated with dietary macronutrient intake. Hum. Mol. Genet. 2013, 22, 1895–1902. [Google Scholar] [CrossRef]
- Meddens, S.F.W.; de Vlaming, R.; Bowers, P.; Burik, C.A.P.; Linner, R.K.; Lee, C.; Okbay, A.; Turley, P.; Rietveld, C.A.; Fontana, M.A.; et al. Genomic analysis of diet composition finds novel loci and associations with health and lifestyle. Mol. Psychiatry 2021, 26, 2056–2069. [Google Scholar] [CrossRef]
- Jensen, T.; Abdelmalek, M.F.; Sullivan, S.; Nadeau, K.J.; Green, M.; Roncal, C.; Nakagawa, T.; Kuwabara, M.; Sato, Y.; Kang, D.H.; et al. Fructose and sugar: A major mediator of non-alcoholic fatty liver disease. J. Hepatol. 2018, 68, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Dushay, J.R.; Toschi, E.; Mitten, E.K.; Fisher, F.M.; Herman, M.A.; Maratos-Flier, E. Fructose ingestion acutely stimulates circulating FGF21 levels in humans. Mol. Metab. 2015, 4, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Falzarano, C.; Lofton, T.; Osei-Ntansah, A.; Oliver, T.; Southward, T.; Stewart, S.; Andrisse, S. Nonalcoholic Fatty Liver Disease in Women and Girls With Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2022, 107, 258–272. [Google Scholar] [CrossRef] [PubMed]
- Won, Y.B.; Seo, S.K.; Yun, B.H.; Cho, S.; Choi, Y.S.; Lee, B.S. Non-alcoholic fatty liver disease in polycystic ovary syndrome women. Sci. Rep. 2021, 11, 7085. [Google Scholar] [CrossRef]
- Jung, J.Y.; Shim, J.J.; Park, S.K.; Ryoo, J.H.; Choi, J.M.; Oh, I.H.; Jung, K.W.; Cho, H.; Ki, M.; Won, Y.J.; et al. Serum ferritin level is associated with liver steatosis and fibrosis in Korean general population. Hepatol. Int. 2019, 13, 222–233. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variables in the Third Survey | Non-Incident NAFLD | Incident NAFLD | p |
|---|---|---|---|
| Number, n (%) | 1980 (56.6) | 1521 (43.4) | - |
| Age (years) | 55.0 ± 8.7 | 56.1 ± 8.6 | 0.0001 |
| BMI (kg/m2) | 25.5 ± 3.2 | 25.3 ± 3.2 | 0.0916 |
| Waist circumference (cm) | 83.9 ± 9.0 | 83.0 ± 8.6 | 0.0025 |
| Systolic blood pressure (mmHg) | 115.8 ± 17.1 | 114.2 ± 16.0 | 0.0076 |
| Diastolic blood pressure (mmHg) | 76.9 ± 10.5 | 76.3 ± 10.0 | 0.0492 |
| Fasting Blood Glucose (mg/dL) | 87.8 ± 8.3 | 93.3± 13.6 | <0.0001 |
| Insulin (µIU/mL) | 6.2 ± 2.0 | 7.4 ± 2.6 | <0.0001 |
| Total cholesterol (mg/dL) | 188.4 ± 31.7 | 194.8 ± 34.4 | <0.0001 |
| Triglyceride (mg/dL) | 107.8 ± 57.7 | 151.9 ± 103.3 | <0.0001 |
| HDL-C (mg/dL) | 47.0 ± 10.3 | 42.3 ± 9.2 | <0.0001 |
| LDL-C (mg/dL) | 120.0 ± 28.4 | 122.3 ± 31.0 | 0.0166 |
| AST (U/L) | 23.1 ± 6.7 | 24.0 ± 6.8 | <0.0001 |
| ALT(U/L) | 18.4 ± 7.5 | 21.8 ± 9.4 | <0.0001 |
| Alcohol intake (g/week) | 17.0 ± 32.6 | 16.8 ± 32.1 | 0.4577 |
| Smoking status, n (%) | - | - | 0.8330 |
| Never smoker | 1266 (64.0) | 986 (64.9) | - |
| Former smoker | 354 (17.9) | 269 (17.7) | - |
| Current smoker | 359 (18.1) | 265 (17.4) | - |
| Education level, n (%) | - | - | <0.0001 |
| Elementary or below | 612 (31.0) | 593 (39.4) | - |
| Junior high school | 412 (20.8) | 316 (20.8) | - |
| High school | 698 (35.3) | 440 (29.0) | - |
| College or above | 254 (12.8) | 170 (11.2) | - |
| Physical activity, METs-hr/wk | 335.5 ± 105.8 | 337.7 ± 110.2 | 0.6882 |
| NLFS | −1.9 ± 0.7 | −1.6 ± 0.6 | <0.0001 |
| Diabetes mellitus, n (%) | 62 (3.1) | 126 (8.3) | <0.0001 |
| Hypertension, n (%) | 364 (18.4) | 234 (15.4) | 0.0194 |
| Hyperlipidemia, n (%) | 685 (34.6) | 877 (57.7) | <0.0001 |
| Total calorie intake (kcal/day) | 1776.8 ± 530.0 | 1762.7 ± 566.3 | 0.0749 |
| Protein (g/day) | 58.4 ± 22.7 | 58.3 ± 25.2 | 0.1218 |
| CHO (g/day) | 322.5 ± 88.7 | 322.2 ± 92.8 | 0.4375 |
| Fat (g/day) | 28.1 ± 17.0 | 26.7 ± 17.1 | 0.0005 |
| Protein (%) (1) | 12.9 ± 2.3 | 12.9 ± 2.4 | 0.3370 |
| CHO (%) (1) | 72.5 ± 6.4 | 73.1 ± 6.5 | 0.0024 |
| Fat (%) (1) | 13.5 ± 5.1 | 12.8 ± 5.1 | <0.0001 |
| Gene | SNP ID | CHR | Position (hg19) | Major Allele | Minor Allele | Risk Allele | MAF | HWE | β Adjusted (1) | HR (95%CI) (1) | SE (1) | p(1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KLB | rs2608819 | 4 | chr4:39429811 | C | T | T | 0.1726 | 0.6820 | 0.1184 | 1.126 (0.840–1.509) | 0.1495 | 0.4286 |
| FGFR1 | rs881301 | 8 | chr8:38332318 | T | C | C | 0.3244 | 0.9034 | 0.0976 | 1.103 (0.929–1.309) | 0.0876 | 0.2653 |
| FGFR2 | rs9420328 | 10 | chr10:123140661 | A | C | C | 0.0996 | 0.9527 | 0.2260 | 1.254 (0.724–2.170) | 0.2800 | 0.4196 |
| rs4751832 | 10 | chr10:123023263 | G | C | G | 0.3124 | 0.5361 | 0.2047 | 1.227 (1.024–1.470) | 0.0921 | 0.0263 | |
| rs7913828 | 10 | chr10:123095255 | G | A | G | 0.3098 | 0.8229 | 0.2629 | 1.301 (1.068–1.583) | 0.1003 | 0.0088 | |
| rs2420941 | 10 | chr10:123229626 | T | G | T | 0.4338 | 0.7618 | 0.1676 | 1.183 (1.018–1.374) | 0.0764 | 0.0283 | |
| rs1649181 | 10 | chr10:123375856 | C | T | C | 0.0105 | 1.0000 | 0.3501 | 1.419 (0.922–2.184) | 0.2200 | 0.1115 | |
| rs17101702 | 10 | chr10:123003707 | G | C | G | 0.1035 | 0.0016 | 0.0408 | 1.042 (0.705–1.539) | 0.1991 | 0.8375 |
| Cases | HR (95% CI) | p-Trend | |
|---|---|---|---|
| All (1) | |||
| First quartile | 864 | 1.00 | <0.0001 |
| Second quartile | 840 | 1.280 (1.098–1.488) | |
| Third quartile | 919 | 1.279 (1.103–1.483) | |
| Fourth quartile | 878 | 1.572 (1.359–1.819) | |
| Male (2) | |||
| First quartile | 396 | 1.00 | 0.0171 |
| Second quartile | 382 | 1.155 (0.915–1.457) | |
| Third quartile | 412 | 1.248 (1.002–1.555) | |
| Fourth quartile | 403 | 1.455 (1.169–1.812) | |
| Female (3) | |||
| First quartile | 468 | 1.00 | <0.0001 |
| Second quartile | 458 | 1.357 (1.104–1.669) | |
| Third quartile | 491 | 1.336 (1.093–1.634) | |
| Fourth quartile | 491 | 1.648 (1.353–2.007) |
| Protein Intake (g/Day) | p | p-Trend | Protein Intake (g/Day) | p | p-Trend | ||||
|---|---|---|---|---|---|---|---|---|---|
| Low | Medium | High | <RNI | ≥RNI | |||||
| All (1) | |||||||||
| unadjusted | 1.084 (0.960–1.226) | 1.012 (0.894–1.147) | 1.00 | 0.3719 | 0.1756 | 1.088 (0.984–1.204) | 1.00 | 0.0992 | 0.1326 |
| model 1 | 0.988 (0.867–1.125) | 0.990 (0.873–1.124) | 1.00 | 0.9809 | 0.5572 | 1.061 (0.957–1.176) | 1.00 | 0.2613 | 0.1305 |
| model 2 | 0.935 (0.820–1.066) | 0.962 (0.847–1.092) | 1.00 | 0.5996 | 0.7173 | 1.026 (0.925–1.138) | 1.00 | 0.6314 | 0.4391 |
| model 3 | 0.982 (0.810–1.191) | 0.990 (0.850–1.152) | 1.00 | 0.9830 | 0.8125 | 1.121 (0.9778–1.285) | 1.00 | 0.0998 | 0.1066 |
| Male (2) | |||||||||
| unadjusted | 0.901 (0.748–1.087) | 1.010 (0.841–1.213) | 1.00 | 0.4264 | 0.1116 | 0.939 (0.807–1.093) | 1.00 | 0.4186 | 0.2141 |
| model 1 | 0.932 (0.769–1.131) | 1.069 (0.889–1.286) | 1.00 | 0.3775 | 0.5033 | 0.971 (0.832–1.133) | 1.00 | 0.7086 | 0.7556 |
| model 2 | 0.942 (0.776–1.143) | 1.080 (0.898–1.300) | 1.00 | 0.3779 | 0.4400 | 0.965 (0.827–1.127) | 1.00 | 0.6569 | 0.6636 |
| model 3 | 1.109 (0.830–1.481) | 1.186 (0.949–1.483) | 1.00 | 0.2954 | 0.6973 | 1.064 (0.864–1.310) | 1.00 | 0.5604 | 0.5315 |
| Female (3) | |||||||||
| unadjusted | 1.304 (1.106–1.539) | 1.140 (0.962–1.351) | 1.00 | 0.0067 | 0.0003 | 1.238 (1.082–1.417) | 1.00 | 0.0019 | 0.0010 |
| model 1 | 1.127 (0.948–1.339) | 1.064 (0.896–1.263) | 1.00 | 0.3992 | 0.0331 | 1.134 (0.988–1.303) | 1.00 | 0.0742 | 0.0403 |
| model 2 | 1.049 (0.881–1.249) | 1.050 (0.883–1.248) | 1.00 | 0.8273 | 0.2002 | 1.069 (0.929–1.230) | 1.00 | 0.3504 | 0.2388 |
| model 3 | 1.161 (0.900–1.498) | 1.117 (0.908–1.374) | 1.00 | 0.4893 | 0.1628 | 1.145 (0.956–1.371) | 1.00 | 0.1417 | 0.2438 |
| PHS | Protein Intake (g/Day) | p- Trend | p- Interaction | Protein Intake (g/Day) | p-Trend | p- Interaction | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cases | Low | Medium | High | <RNI | ≥RNI | |||||
| All (1) | 0.0189 | 0.8363 | ||||||||
| First quartile | 864 | 1.244 (0.800–1.934) | 0.861 (0.603–1.229) | 1.00 | 0.0105 | - | 1.494 (1.087–2.054) | 1.00 | 0.0287 | - |
| Second quartile | 840 | 0.830 (0.559–1.232) | 0.908 (0.667–1.237) | 1.00 | 0.6085 | - | 1.045 (0.788–1.386) | 1.00 | 0.8925 | - |
| Third quartile | 919 | 0.873 (0.603–1.263) | 0.843 (0.628–1.132) | 1.00 | 0.4591 | - | 1.004 (0.774–1.302) | 1.00 | 0.8165 | - |
| Fourth quartile | 878 | 1.062 (0.740–1.524) | 1.464 (1.103–1.945) | 1.00 | 0.0366 | - | 1.075 (0.835–1.384) | 1.00 | 0.3938 | - |
| Male (2) | 0.2071 | 0.1090 | ||||||||
| First quartile | 396 | 1.087 (0.571–2.072) | 1.125 (0.688–1.840) | 1.00 | 0.8071 | - | 1.060 (0.668–1.684) | 1.00 | 0.6988 | - |
| Second quartile | 382 | 1.858 (0.964–3.582) | 1.794 (1.084–2.970) | 1.00 | 0.1571 | - | 1.252 (0.794–1.974) | 1.00 | 0.4704 | - |
| Third quartile | 412 | 0.721 (0.402–1.292) | 0.823 (0.527–1.286) | 1.00 | 0.6315 | - | 0.854 (0.575–1.269) | 1.00 | 0.7186 | - |
| Fourth quartile | 403 | 1.529 (0.900–2.598) | 1.614 (1.060–2.458) | 1.00 | 0.3049 | - | 1.397 (0.937–2.083) | 1.00 | 0.1200 | - |
| Female (3) | 0.0131 | 0.0361 | ||||||||
| First quartile | 468 | 2.921 (1.523–5.602) | 1.803 (1.054–3.083) | 1.00 | <0.0001 | - | 2.021 (1.276–3.200) | 1.00 | 0.0016 | - |
| Second quartile | 458 | 0.649 (0.392–1.075) | 0.835 (0.553–1.261) | 1.00 | 0.5836 | - | 0.920 (0.636–1.330) | 1.00 | 0.5114 | - |
| Third quartile | 491 | 1.171 (0.711–1.927) | 0.861 (0.571–1.299) | 1.00 | 0.4094 | - | 1.176 (0.830–1.666) | 1.00 | 0.8531 | - |
| Fourth quartile | 491 | 1.016 (0.627–1.646) | 1.435 (0.986–2.089) | 1.00 | 0.0203 | - | 0.877 (0.629–1.222) | 1.00 | 0.4348 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.J.; Shon, J.; Park, Y.J. Association of NAFLD with FGF21 Polygenic Hazard Score, and Its Interaction with Protein Intake Level in Korean Adults. Nutrients 2023, 15, 2385. https://doi.org/10.3390/nu15102385
Lee HJ, Shon J, Park YJ. Association of NAFLD with FGF21 Polygenic Hazard Score, and Its Interaction with Protein Intake Level in Korean Adults. Nutrients. 2023; 15(10):2385. https://doi.org/10.3390/nu15102385
Chicago/Turabian StyleLee, Hae Jin, Jinyoung Shon, and Yoon Jung Park. 2023. "Association of NAFLD with FGF21 Polygenic Hazard Score, and Its Interaction with Protein Intake Level in Korean Adults" Nutrients 15, no. 10: 2385. https://doi.org/10.3390/nu15102385
APA StyleLee, H. J., Shon, J., & Park, Y. J. (2023). Association of NAFLD with FGF21 Polygenic Hazard Score, and Its Interaction with Protein Intake Level in Korean Adults. Nutrients, 15(10), 2385. https://doi.org/10.3390/nu15102385