
Myristica fragrans Extract Regulates Gut Microbes and Metabolites to Attenuate Hepatic Inflammation and Lipid Metabolism Disorders via the AhR–FAS and NF-κB Signaling Pathways in Mice with Non-Alcoholic Fatty Liver Disease

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of MFE and Identification of the Main Components
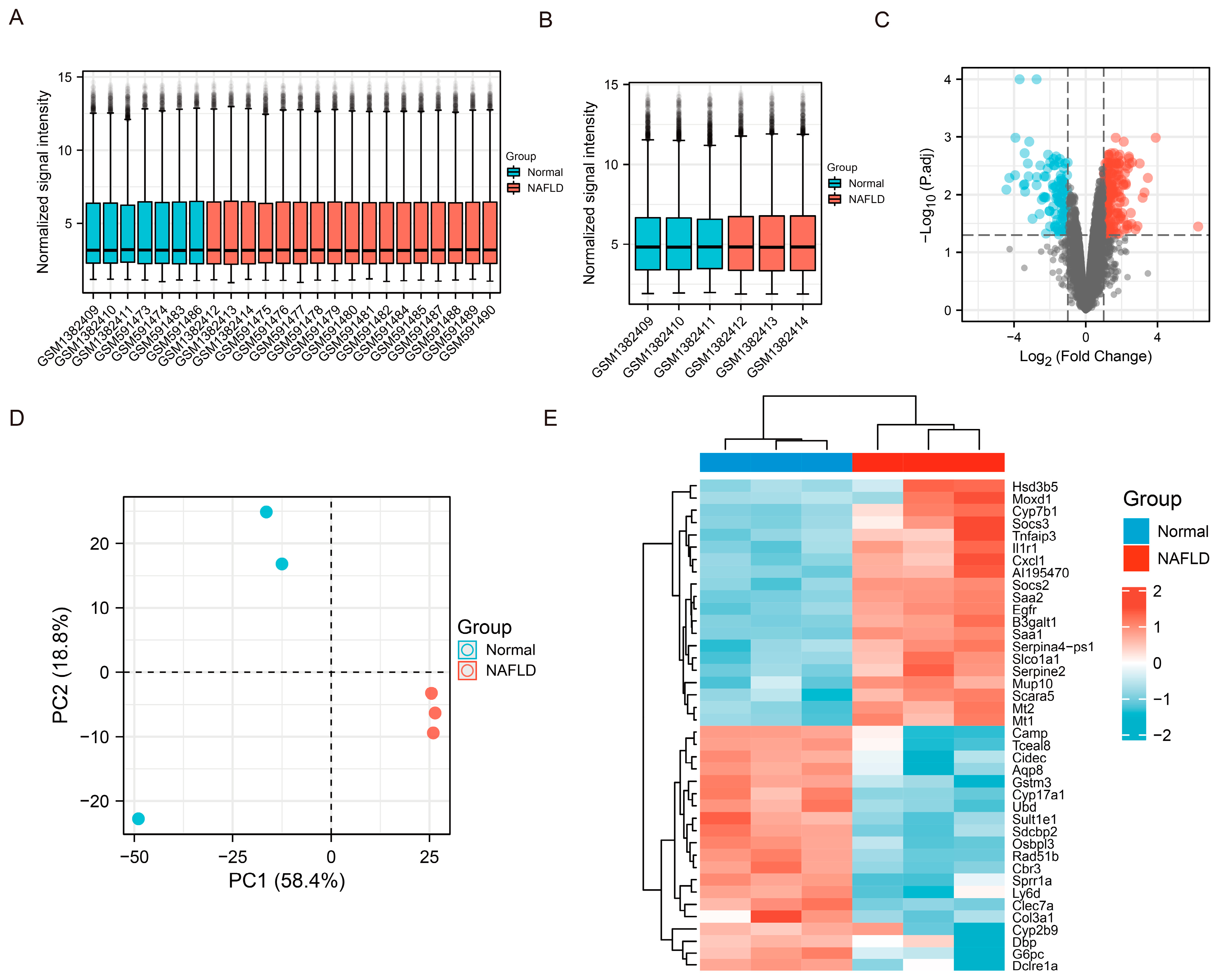
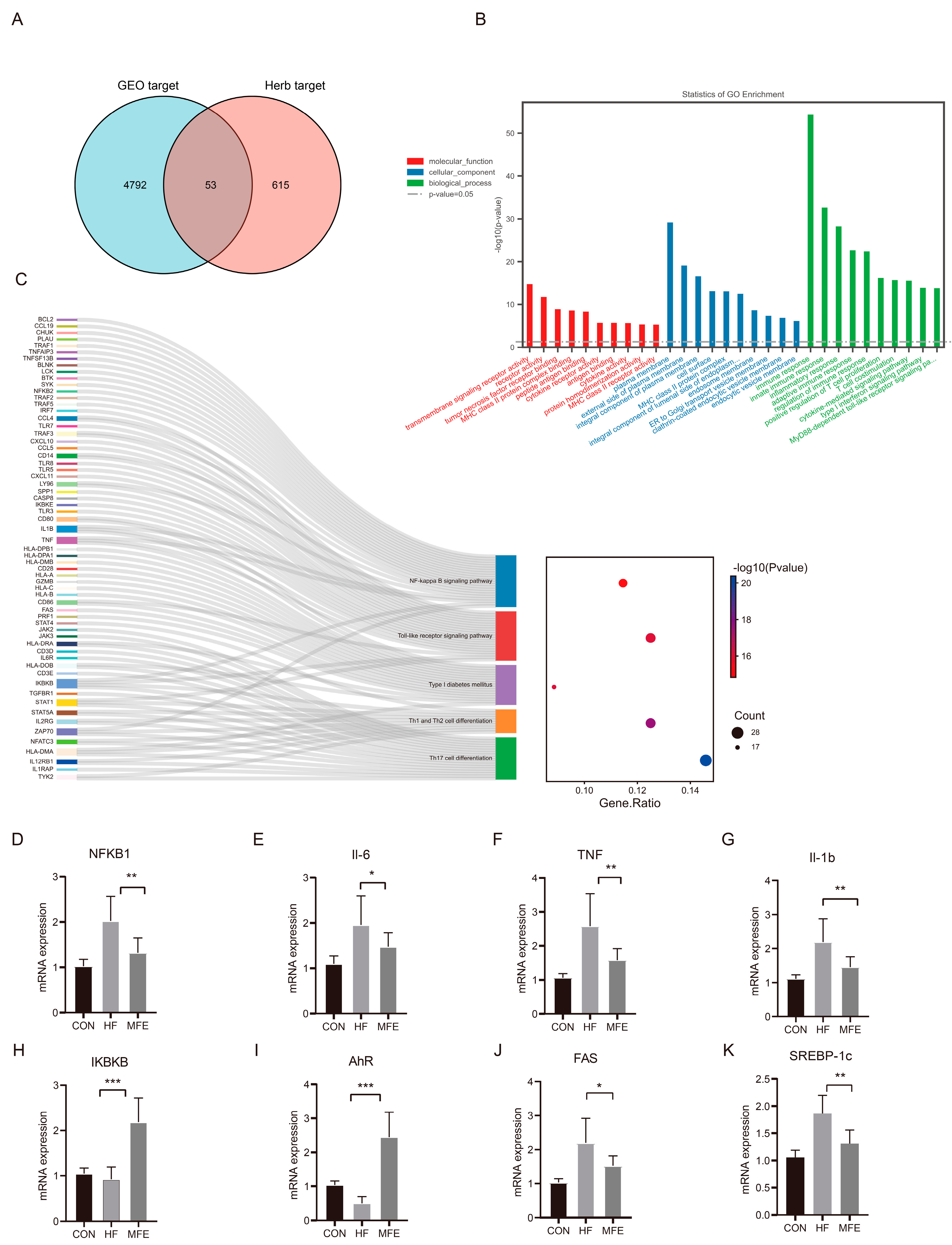
2.2. Gene Expression Omnibus and Herb Database Mining
2.3. Animal Experiments
2.4. Serum Biochemical Markers in Mice
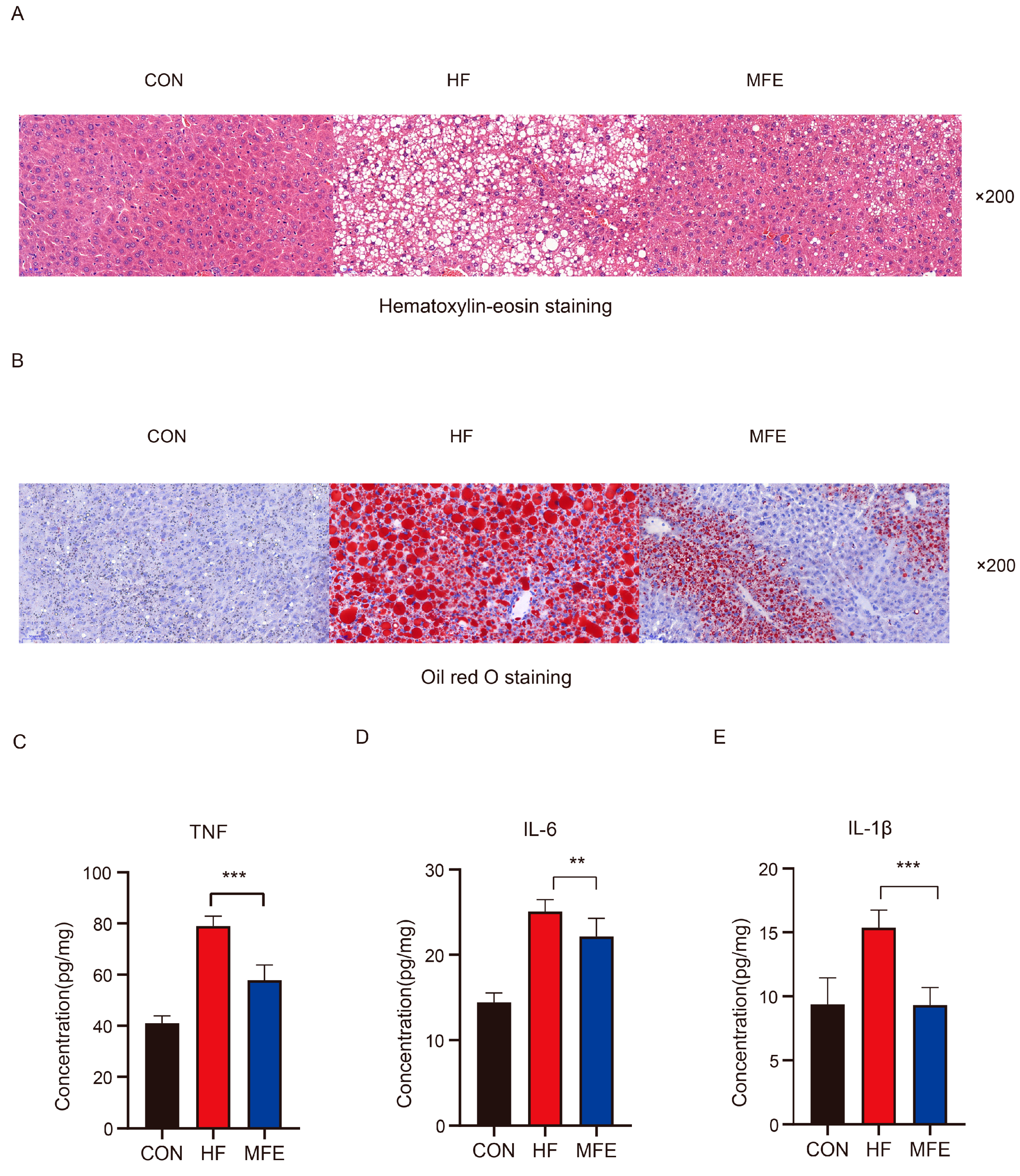
2.5. Hematoxylin & Eosin and Oil Red O Staining
2.6. Quantitative Reverse Transcription Polymerase Chain Reaction Analysis
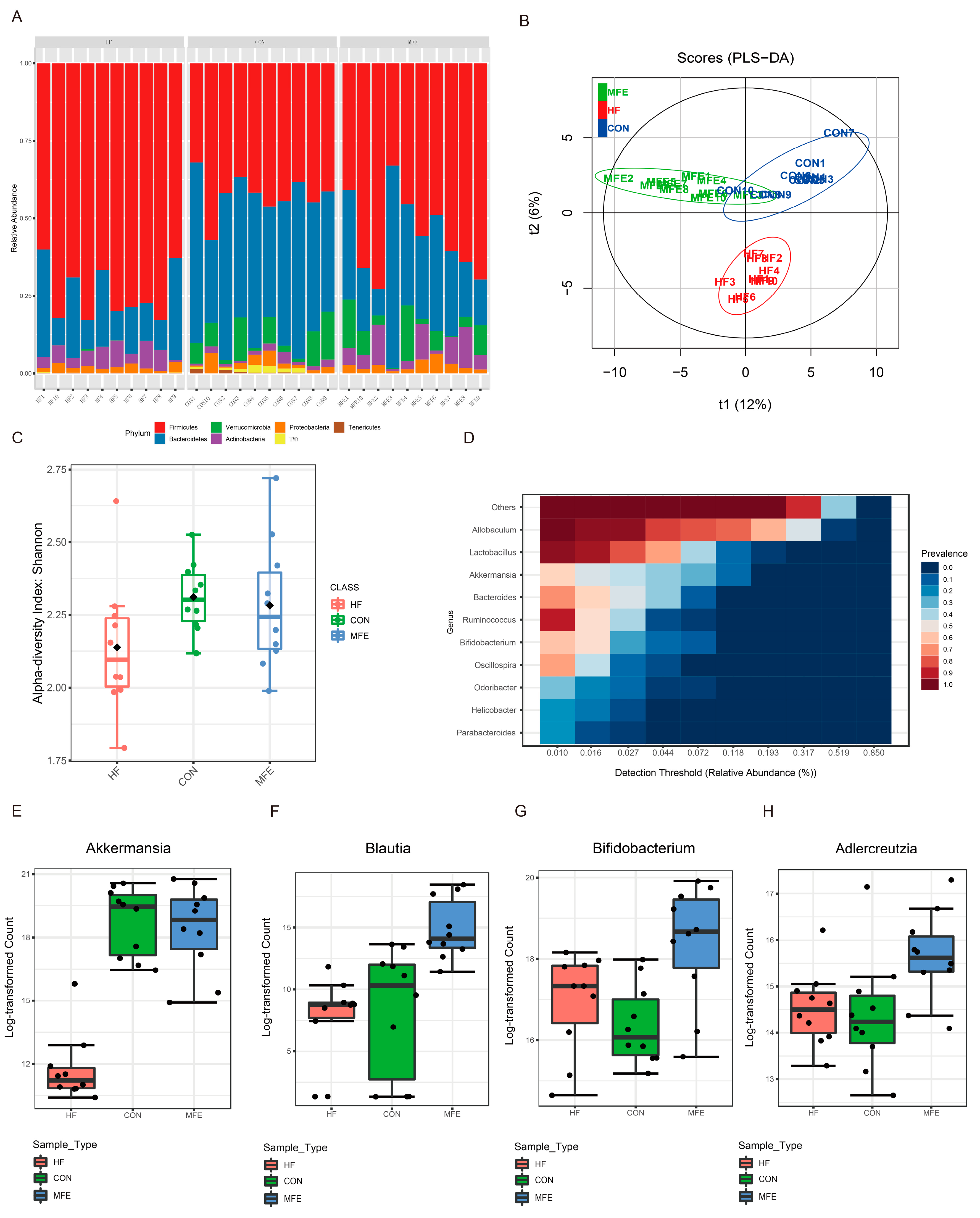
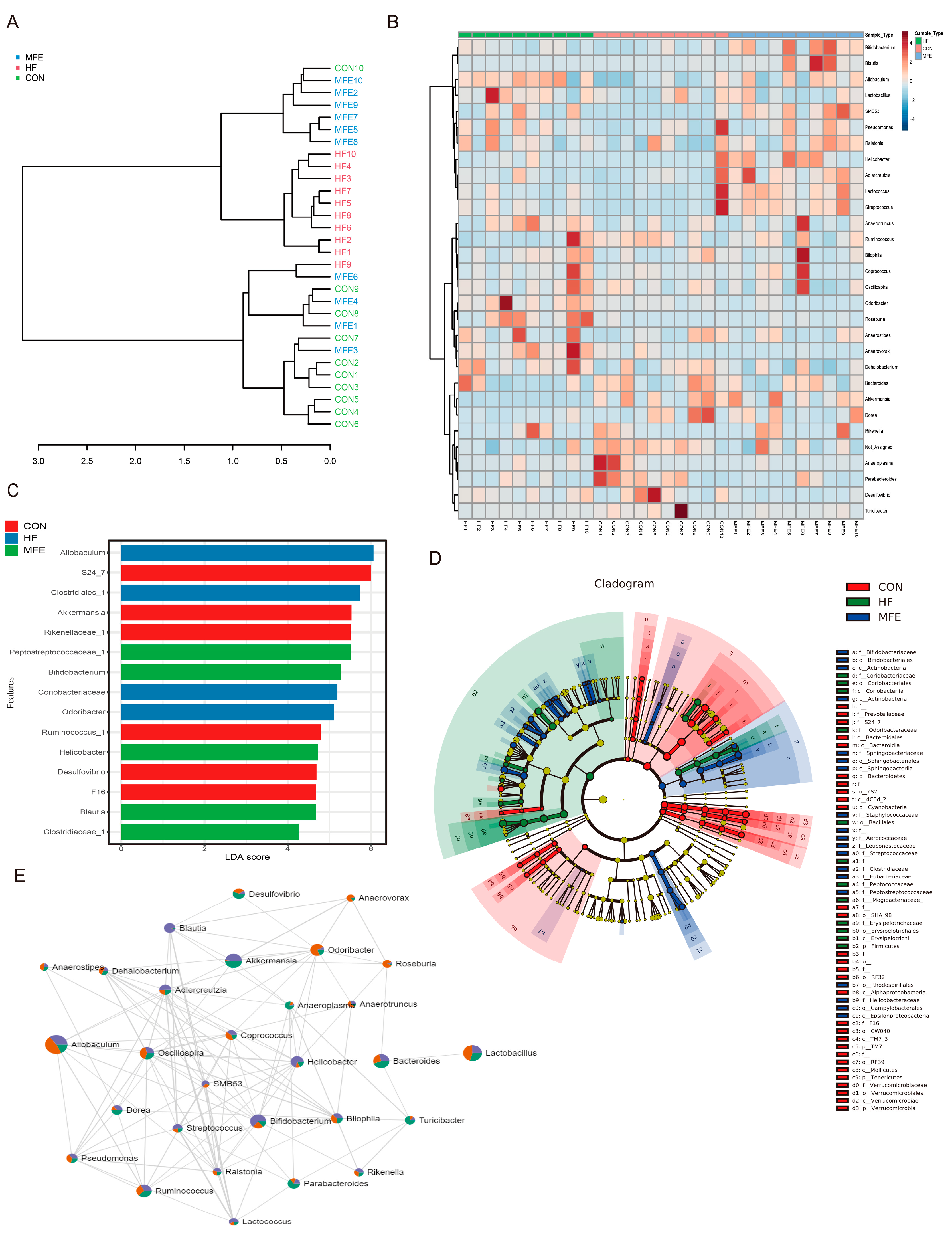
2.7. Gut Microbiota Analysis by 16S rRNA Gene Sequencing
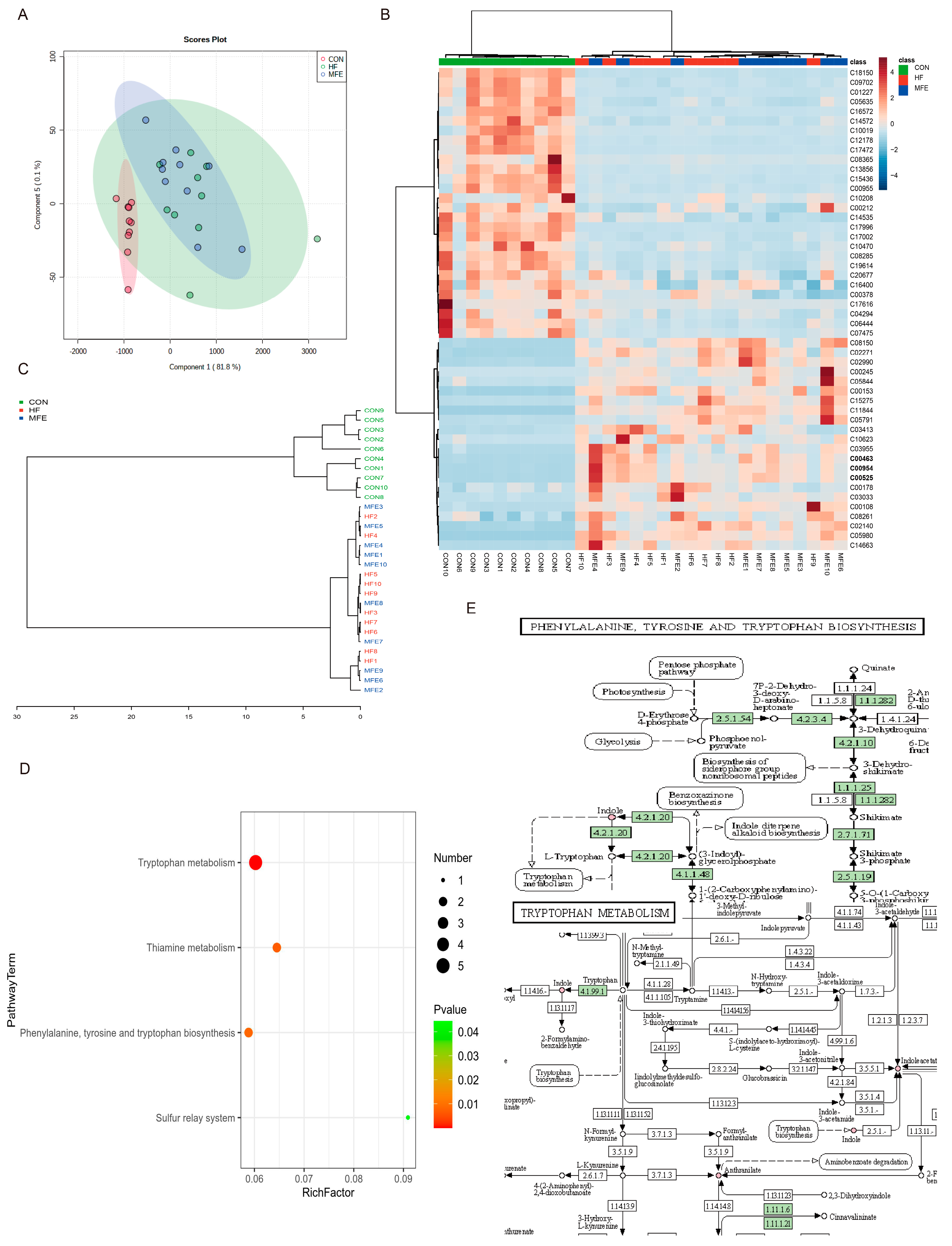
2.8. Non-Targeted Metabolomics Analysis
2.9. Statistical Analysis
3. Results
3.1. MFE Principal Component Analysis and Identification
3.2. Alleviation Effect of MFE on High-Fat-Diet-Induced NAFLD in Mice
3.3. Modulation of the Gut Microbiota by MFE in Mice Fed a High-Fat Diet
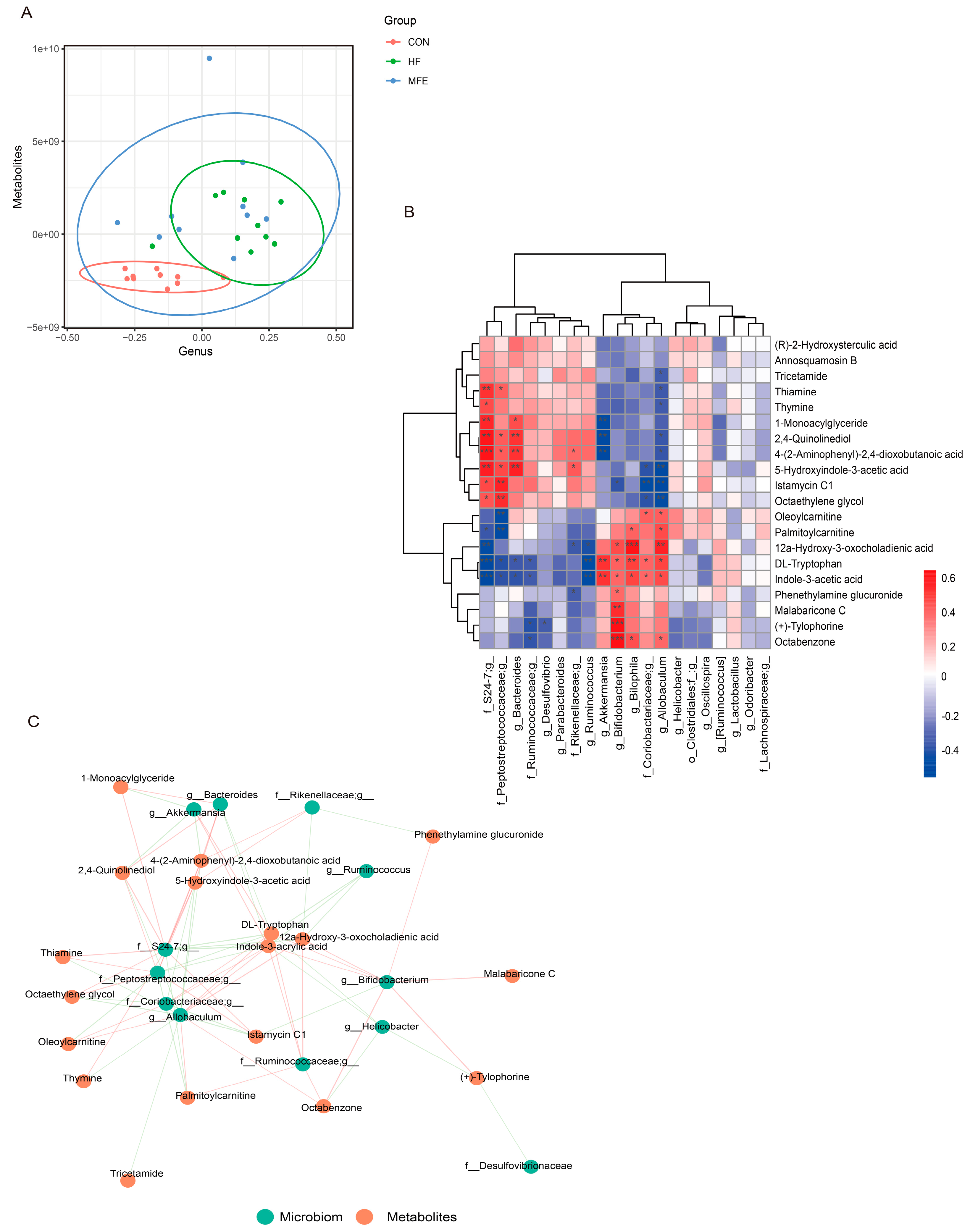
3.4. Effects of MFE on Intestinal Metabolites in Mice Fed a High-Fat Diet
3.5. Omics Analysis of the Intestinal Flora Metabolites in NAFLD Mice after MFE Treatment
3.6. MFE Alleviates Hepatic Fat Accumulation and the Inflammatory Response by Regulating the Aryl Hydrocarbon Receptor–Fatty Acid Synthase/Sterol Regulatory Element-Binding Protein 1c and Nuclear Factor Kappa B Signaling Pathways in Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brunt, E.M.; Wong, W.S.; Nobili, V.; Day, C.P.; Rinella, M.E. Nonalcoholic fatty liver disease. Nat. Rev. Dis. Primers 2015, 1, 15080. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, A.; Bonelli, P.; Tuccillo, F.M.; Goldfine, I.D.; Evans, J.L.; Buonaguro, F.M.; Mancini, A. Role of gut microbiota and oxidative stress in the progression of non-alcoholic fatty liver disease to hepatocarcinoma: Current and innovative therapeutic approaches. Redox Biol. 2018, 15, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Bedossa, P. Pathology of non-alcoholic fatty liver disease. Liver Int. 2017, 37, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Jiang, Y.; Wang, M.; Melaku, M.; Liu, L.; Zhao, Y.; Everaert, N.; Yi, B.; Zhang, H. Intestinal dysbiosis in nonalcoholic fatty liver disease (NAFLD): Focusing on the gut–liver axis. Crit. Rev. Food Sci. Nutr. 2021, 1–18. [Google Scholar] [CrossRef]
- Rahman, K.; Desai, C.; Iyer, S.S.; Thorn, N.E.; Kumar, P.; Liu, Y.; Smith, T.; Neish, A.S.; Li, H.; Tan, S. Loss of Junctional Adhesion Molecule A Promotes Severe Steatohepatitis in Mice on a Diet High in Saturated Fat, Fructose, and Cholesterol-ScienceDirect. Gastroenterology 2016, 151, 733–746. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Liang, X.; Wang, K.; Lin, J.; Wang, X.; Wang, P.; Zhang, Y.; Nie, Q.; Liu, H.; Zhang, Z. Intestinal hypoxia-inducible factor 2α regulates lactate levels to shape the gut microbiome and alter thermogenesis. Cell Metab. 2021, 33, 1988–2003.e1987. [Google Scholar] [CrossRef]
- Krishnan, S.; Ding, Y.; Saedi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 2018, 23, 1099–1111. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Tremaroli, V.; Backhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 71, 242–249. [Google Scholar] [CrossRef]
- Miura, K.; Ishioka, M.; Iijima, K. The Roles of the Gut Microbiota and Toll-like Receptors in Obesity and Nonalcoholic Fatty Liver Disease. J. Obes. Metab. Syndr. 2017, 26, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.S.; Youn, G.S.; Choi, J.; Kim, C.H.; Kim, B.Y.; Yang, S.J.; Lee, J.H.; Park, T.S.; Kim, B.K.; Kim, Y.B. Lactobacillus lactis and Pediococcus pentosaceus-driven reprogramming of gut microbiome and metabolome ameliorates the progression of non-alcoholic fatty liver disease. Clin. Transl. Med. 2021, 11, e634. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Knauf, C.; Delzenne, N. Metabolic endotoxemia initiates obesity and insulin resistance. Ann. Nutr. Metab. 2007, 51, 79–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharpton, S.R.; Maraj, B.; Harding-Theobald, E.; Vittinghoff, E.; Terrault, N.A. Gut microbiome–targeted therapies in nonalcoholic fatty liver disease: A systematic review, meta-analysis, and meta-regression. Am. J. Clin. Nutr. 2019, 110, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Ohnishi, H. Role of gut microbiota and Toll-like receptors in nonalcoholic fatty liver disease. World J. Gastroenterol. WJG 2014, 20, 7381. [Google Scholar] [CrossRef]
- Loman, B.R.; Hernández-Saavedra, D.; An, R.; Rector, R.S. Prebiotic and probiotic treatment of nonalcoholic fatty liver disease: A systematic review and meta-analysis. Nutr. Rev. 2018, 76, 822–839. [Google Scholar] [CrossRef]
- Chen, J.; Wang, R.; Li, X.F.; Wang, R.L. Bifidobacterium adolescentis supplementation ameliorates visceral fat accumulation and insulin sensitivity in an experimental model of the metabolic syndrome. Br. J. Nutr. 2012, 107, 1429–1434. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.H.; Liu, X.Z.; Pan, W.; Zou, D.J. Berberine protects against diet-induced obesity through regulating metabolic endotoxemia and gut hormone levels. Mol. Med. Rep. 2017, 15, 2765–2787. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.M.; Young, J.D. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2017, 6, 7489. [Google Scholar] [CrossRef] [Green Version]
- Pathak, S.; Ojha, V. The component glycerides of nutmeg butter (Myristica fragrans). J. Sci. Food Agric. 1957, 8, 537–540. [Google Scholar] [CrossRef]
- Maya, K.; Zachariah, T.J.; Krishnamurthy, K.; Rema, J.; Krishnamoorthy, B. Fatty acids and leaf amino acids in Myristica fragrans Houtt and related taxa. Indian J. Hortic. 2006, 63, 316–318. [Google Scholar]
- Zhao, W.; Song, F.; Hu, D.; Chen, H.; Wang, G. The Protective Effect of Myristica fragrans Houtt Extracts Against Obesity and Inflammation by Regulating Free Fatty Acids Metabolism in Nonalcoholic Fatty Liver Disease. Nutrients 2020, 12, 2507. [Google Scholar] [CrossRef] [PubMed]
- Le, T.V.T.; Nguyen, P.H.; Choi, H.S.; Yang, J.-L.; Kang, K.W.; Ahn, S.-G.; Oh, W.K. Diarylbutane-type lignans from Myristica fragrans (Nutmeg) show the cytotoxicity against breast cancer cells through activation of AMP-activated protein kinase. Nat. Prod. Sci. 2017, 23, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.D.; Bansal, V.K.; Babu, V.; Maithil, N. Chemistry, antioxidant and antimicrobial potential of nutmeg (Myristica fragrans) Houtt. J. Genet. Eng. Biotechnol. 2013, 11, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Olaleye, M.T.; Akinmoladun, A.C.; Akindahunsi, A.A. Antioxidant properties of Myristica fragrans (Houtt) and its effect on selected organs of albino rats. Afr. J. Biotechnol. 2013, 5, 15. [Google Scholar]
- Yang, X.N.; Liu, X.M.; Fang, J.H.; Zhu, X.; Yang, X.W.; Xiao, X.R.; Huang, J.F.; Gonzalez, F.J.; Li, F. PPAR± Mediates the Hepatoprotective Effects of Nutmeg. J. Proteome Res. 2018, 17, 1887–1897. [Google Scholar] [CrossRef]
- Nguyen, P.H.; Le, T.V.T.; Kang, H.W.; Chae, J.; Kim, S.K.; Kwon, K.I.; Seo, D.B.; Lee, S.J.; Oh, W.K. AMP-activated protein kinase (AMPK) activators from Myristica fragrans (nutmeg) and their anti-obesity effect. Bioorganic Med. Chem. Lett. 2010, 20, 4128–4131. [Google Scholar] [CrossRef]
- Kong, Q.; Wang, B.; Tian, P.; Li, X.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. Daily intake of Lactobacillus alleviates autistic-like behaviors by ameliorating the 5-hydroxytryptamine metabolic disorder in VPA-treated rats during weaning and sexual maturation. Food Funct. 2021, 12, 2591–2604. [Google Scholar] [CrossRef]
- Khalil, A.; Giribabu, N.; Yelumalai, S.; Shahzad, H.; Salleh, N. Myristic acid defends against testicular oxidative stress, inflammation, apoptosis: Restoration of spermatogenesis, steroidogenesis in diabetic rats. Life Sci. 2021, 278, 119605. [Google Scholar] [CrossRef]
- Choi, Y.K.; Cho, G.S.; Hwang, S.; Kim, B.W.; Lim, J.H.; Lee, J.C.; Kim, H.C.; Kim, W.K.; Kim, Y.S. Methyleugenol reduces cerebral ischemic injury by suppression of oxidative injury and inflammation. Free Radic. Res. 2010, 44, 925. [Google Scholar] [CrossRef]
- Yang, L.; Yu, S.; Yang, Y.; Wu, H.; Zhang, X.; Lei, Y.; Lei, Z. Berberine improves liver injury induced glucose and lipid metabolic disorders via alleviating ER stress of hepatocytes and modulating gut microbiota in mice. Bioorganic Med. Chem. 2021, 55, 116598. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Hul, M.V.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2013, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Mithieux, G. Does Akkermansia muciniphila play a role in type 1 diabetes? Gut 2018, 67, gutjnl-2017-315732. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Chen, W. Blautia —a new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Benítez-Páez, A.; Pugar, E.; López-Almela, I.; Moya-Pérez, N.; Sanz, Y. Depletion of Blautia Species in the Microbiota of Obese Children Relates to Intestinal Inflammation and Metabolic Phenotype Worsening. mSystems 2020, 5, e00857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alderton, G.K. Tumour immunology: Intestinal bacteria are in command. Nat. Rev. Cancer 2016, 16, 4. [Google Scholar] [CrossRef]
- Butler, C.; Lau, M.; Gillespie, D.; Owen-Jones, E.; Francis, N.A. Effect of Probiotic Use on Antibiotic Administration Among Care Home Residents: A Randomized Clinical Trial. JAMA J. Am. Med. Assoc. 2020, 324, 47. [Google Scholar] [CrossRef]
- Ziętak, M.; Kovatcheva-Datchary, P.; Markiewicz, L.H.; Ståhlman, M.; Kozak, L.P.; Bäckhed, F. Altered microbiota contributes to reduced diet-induced obesity upon cold exposure. Cell Metab. 2016, 23, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Mawe, G.M.; Hoffman, J.M. Serotonin signalling in the gut--functions, dysfunctions and therapeutic targets. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 564–564. [Google Scholar] [CrossRef] [Green Version]
- Boon, N.; Kaur, M.; Aziz, A.; Bradnick, M.; Lund, P.A. The Signaling Molecule Indole Inhibits Induction of the AR2 Acid Resistance System in Escherichia coli. Front. Microbiol. 2020, 11, 474. [Google Scholar] [CrossRef] [Green Version]
- Kiss, E.A.; Vonarbourg, C.; Kopfmann, S.; Hobeika, E.; Finke, D.; Esser, C.; Diefenbach, A. Natural Aryl Hydrocarbon Receptor Ligands Control Organogenesis of Intestinal Lymphoid Follicles. Science 2012, 334, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Hankinson, O. The aryl hydrocarbon receptor complex. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 307–340. [Google Scholar] [CrossRef] [PubMed]
- Chaves Filho, A.J.M.; Lima, C.N.C.; Vasconcelos, S.M.M.; De Lucena, D.F.; Maes, M.; Macedo, D. IDO chronic immune activation and tryptophan metabolic pathway: A potential pathophysiological link between depression and obesity. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 80, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Devlin, A.; Marcobal, A.; Dodd, D.; Nayfach, S.; Plummer, N.; Meyer, T.; Pollard, K.; Sonnenburg, J.; Fischbach, M. Modulation of a Circulating Uremic Solute via Rational Genetic Manipulation of the Gut Microbiota. Cell Host Microbe 2016, 20, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Zhang, M.; Qi, H.; Gao, Y.; Yang, Y.; Yun, H.; Zhang, Q.; Yang, X.; Zhang, Y.; He, J. Gut microbiota–derived metabolite 3-idoleacetic acid together with LPS induces IL-35+ B cell generation. Microbiome 2022, 10, 1–20. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content (%) |
|---|---|
| Tetradecanoic acid | 8.57 |
| Licarin A | 9.94 |
| Licarin B | 6.46 |
| Methyleugenol | 2.84 |
| Isoelemicin | 2.55 |
| Cedrol | 1.72 |
| Melezitose | 2.14 |
| Sucrose | 1.32 |
| Otobain | 1.37 |
| CON | HF | MFE | |
|---|---|---|---|
| Food intake (g/day) | 2.92 ± 0.27 a | 2.74 ± 0.21 a | 2.94 ± 0.08 a |
| Final body weight (g) | 31.61 ± 1.09 c | 39.54 ± 1.39 a | 34.27 ± 1.25 b |
| Body weight gain (g) | 12.71 ± 1.41 c | 21.25 ± 1.27 a | 16.01 ± 1.29 b |
| Liver weight (g) | 1.29 ± 0.19 c | 1.94 ± 0.21 a | 1.56 ± 0.17 b |
| Liver index (%) | 4.09 ± 0.57 ab | 4.96 ± 0.59 a | 4.53 ± 0.42 a |
| CON | HF | MFE | |
|---|---|---|---|
| TC (U/L) | 2.15 ± 0.39 b | 3.59 ± 0.51 a | 2.44 ± 0.57 b |
| TG (U/L) | 0.45 ± 0.07 b | 0.72 ± 0.09 a | 0.51 ± 0.08 b |
| HDL-c (mmol/L) | 1.91 ± 0.23 a | 1.99 ± 0.13 a | 2.10 ± 0.31 a |
| LDL-c (mmol/L) | 0.46 ± 0.18 b | 0.93 ± 0.21 a | 0.47 ± 0.11 b |
| AST (mmol/L) | 65.76 ± 10.52 c | 130.98 ± 16.75 a | 90.98 ± 13.99 b |
| ALT (mmol/L) | 54.15 ± 16.9 b | 198.33 ± 30.78 a | 75.35 ± 23.17 b |
| Glu (mmol/L) | 4.89 ± 0.5 c | 6.97 ± 0.89 a | 5.85 ± 0.83 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Guo, M.; Feng, J.; Gu, Z.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Myristica fragrans Extract Regulates Gut Microbes and Metabolites to Attenuate Hepatic Inflammation and Lipid Metabolism Disorders via the AhR–FAS and NF-κB Signaling Pathways in Mice with Non-Alcoholic Fatty Liver Disease. Nutrients 2022, 14, 1699. https://doi.org/10.3390/nu14091699
Zhao W, Guo M, Feng J, Gu Z, Zhao J, Zhang H, Wang G, Chen W. Myristica fragrans Extract Regulates Gut Microbes and Metabolites to Attenuate Hepatic Inflammation and Lipid Metabolism Disorders via the AhR–FAS and NF-κB Signaling Pathways in Mice with Non-Alcoholic Fatty Liver Disease. Nutrients. 2022; 14(9):1699. https://doi.org/10.3390/nu14091699
Chicago/Turabian StyleZhao, Wenyu, Min Guo, Jun Feng, Zhennan Gu, Jianxin Zhao, Hao Zhang, Gang Wang, and Wei Chen. 2022. "Myristica fragrans Extract Regulates Gut Microbes and Metabolites to Attenuate Hepatic Inflammation and Lipid Metabolism Disorders via the AhR–FAS and NF-κB Signaling Pathways in Mice with Non-Alcoholic Fatty Liver Disease" Nutrients 14, no. 9: 1699. https://doi.org/10.3390/nu14091699
APA StyleZhao, W., Guo, M., Feng, J., Gu, Z., Zhao, J., Zhang, H., Wang, G., & Chen, W. (2022). Myristica fragrans Extract Regulates Gut Microbes and Metabolites to Attenuate Hepatic Inflammation and Lipid Metabolism Disorders via the AhR–FAS and NF-κB Signaling Pathways in Mice with Non-Alcoholic Fatty Liver Disease. Nutrients, 14(9), 1699. https://doi.org/10.3390/nu14091699