
Oral Acid Load Down-Regulates Fibroblast Growth Factor 23

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. In Vivo Experiments
2.2.1. Animals and Diets
2.2.2. Generation of Kidney Disease
2.2.3. Parathyroidectomy
2.2.4. Induction of Acidosis
2.2.5. Experimental Design
- (a)
- Acidosis with intact parathyroid function
- (b)
- Acidosis with parathyroidectomy
2.2.6. Blood and Urine Chemistries
2.3. In Vitro Experiments
2.3.1. Cell Culture
2.3.2. RNA Extraction and Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.4. Statistical Analysis
3. Results
3.1. In Vivo Studies
3.1.1. Renal Function
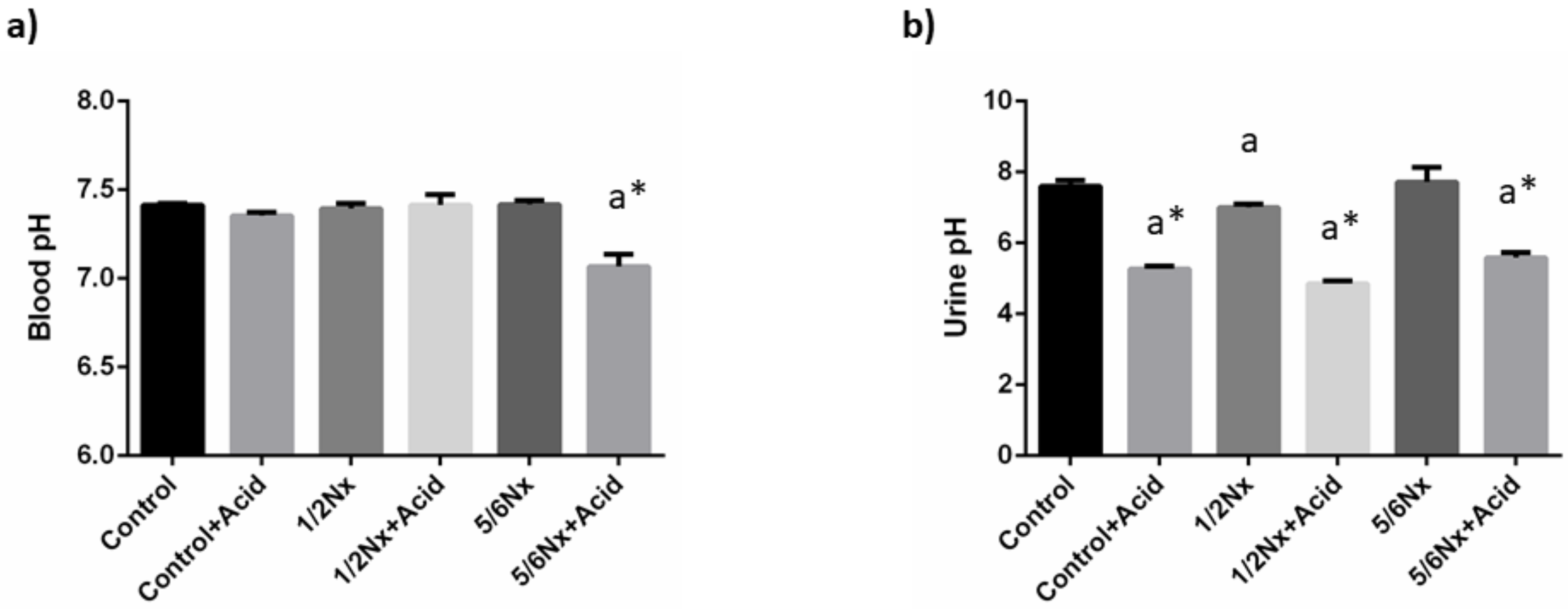
3.1.2. Acid-Base Balance
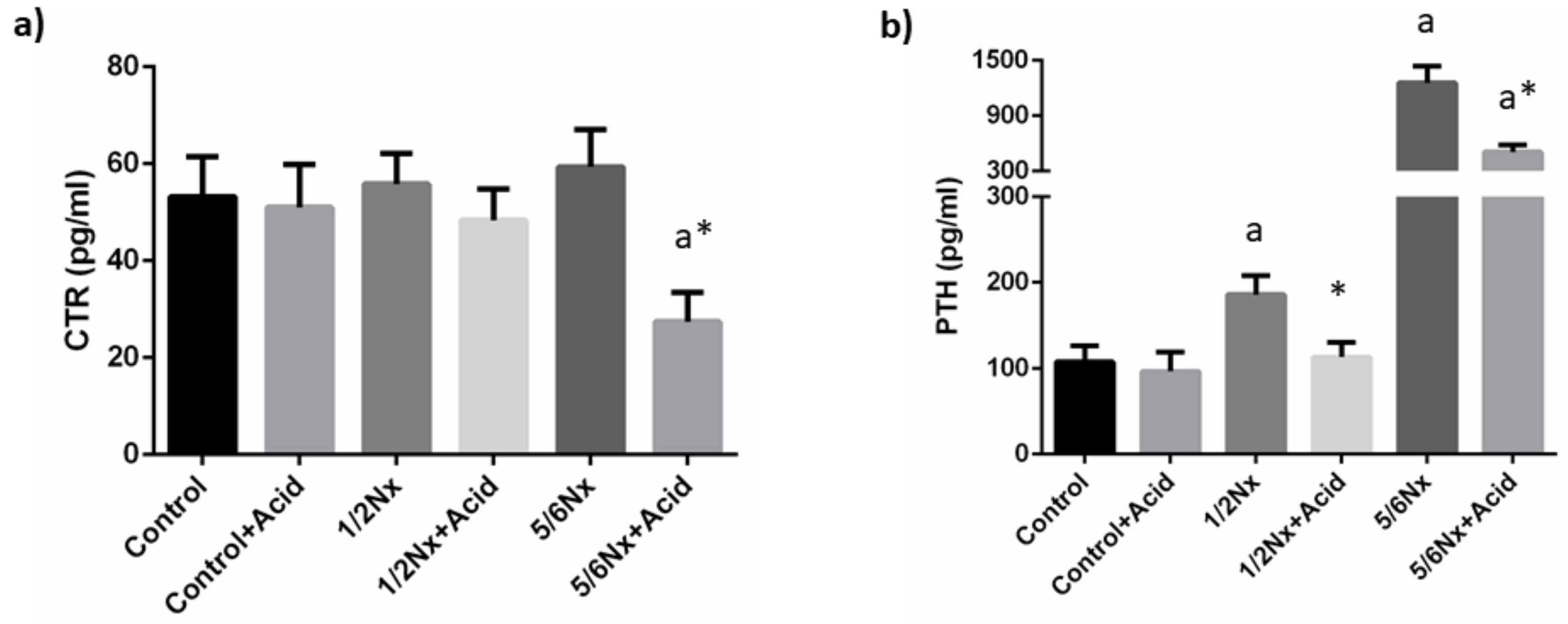
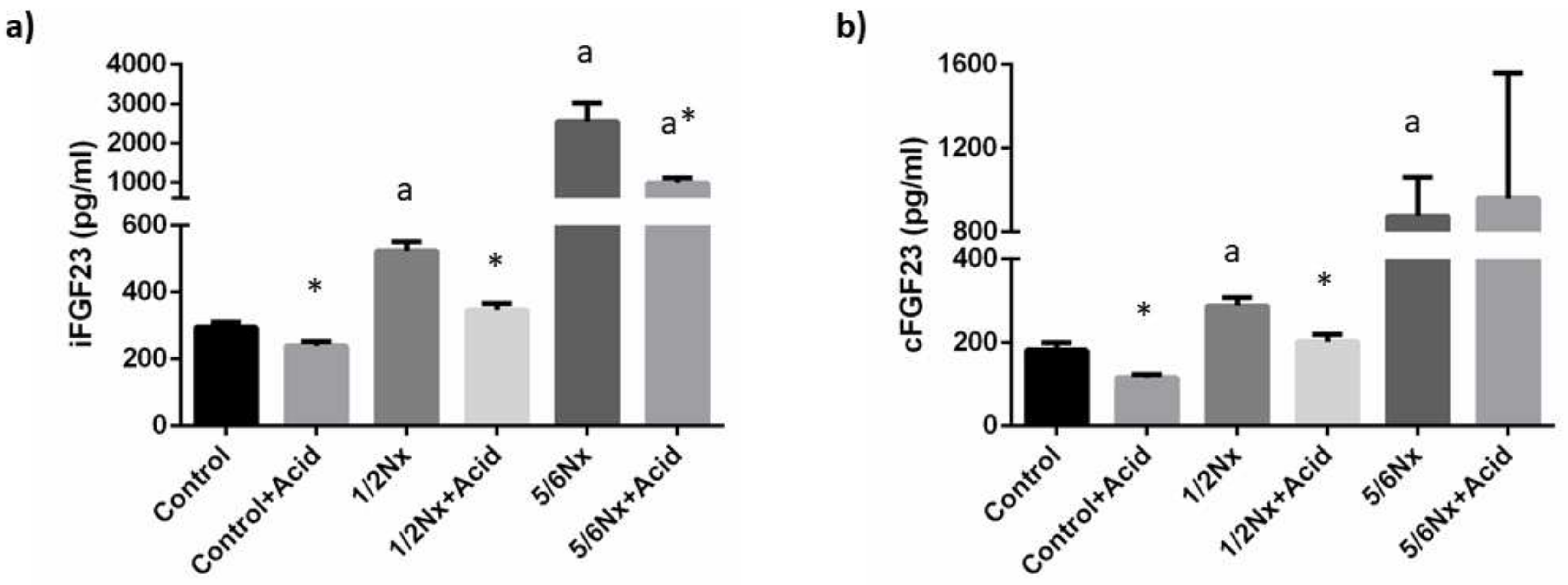
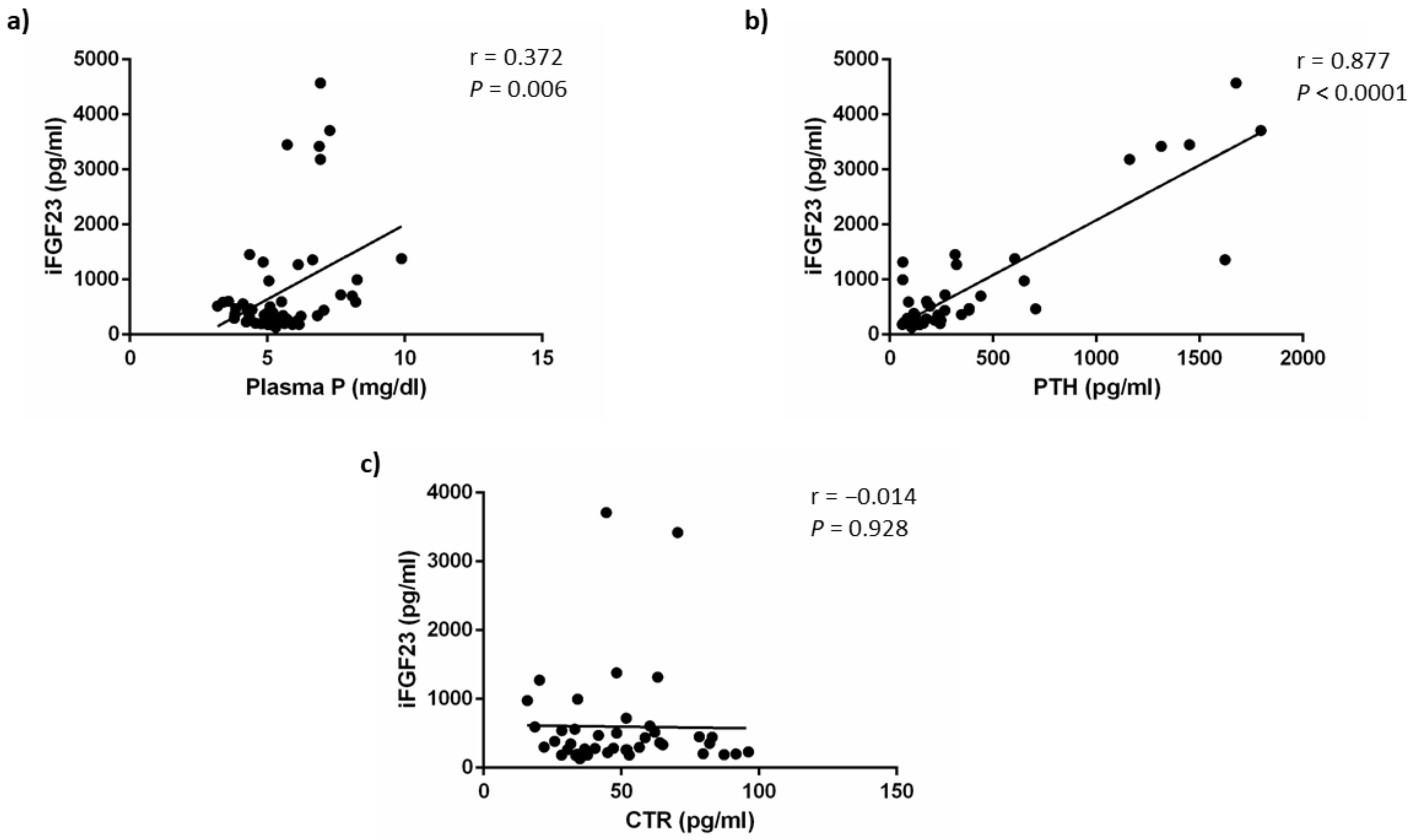
3.1.3. Mineral Metabolism
3.2. In Vitro Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Osuna-Padilla, I.A.; Leal-Escobar, G.; Garza-García, C.A.; Rodríguez-Castellanos, F.E. Dietary Acid Load: Mechanisms and evidence of its health repercussions. Nefrologia 2019, 39, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Kraut, J.A.; Kurtz, I. Metabolic Acidosis of CKD: Diagnosis, Clinical Characteristics, and Treatment. Am. J. Kidney Dis. 2005, 45, 978–993. [Google Scholar] [CrossRef]
- Bürki, R.; Mohebbi, N.; Bettoni, C.; Wang, X.; Serra, A.L.; Wagner, C.A. Impaired expression of key molecules of ammoniagenesis underlies renal acidosis in a rat model of chronic kidney disease. Nephrol. Dial. Transplant. 2014, 30, 770–781. [Google Scholar] [CrossRef] [Green Version]
- Raphael, K.L. Metabolic Acidosis in CKD: Core Curriculum 2019. Am. J. Kidney Dis. 2019, 74, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Levy, D.S.; Abramowitz, M.K. Acid Base Balance and Progression of Kidney Disease. Semin. Nephrol. 2019, 39, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Kovesdy, C.P.; Anderson, J.E.; Kalantar-Zadeh, K. Association of serum bicarbonate levels with mortality in patients with non-dialysis-dependent CKD. Nephrol. Dial. Transplant. 2008, 24, 1232–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vervloet, M. Renal and extrarenal effects of fibroblast growth factor 23. Nat. Rev. Nephrol. 2018, 15, 109–120. [Google Scholar] [CrossRef]
- Shimada, T.; Hasegawa, H.; Yamazaki, Y.; Muto, T.; Hino, R.; Takeuchi, Y.; Fujita, T.; Nakahara, K.; Fukumoto, S.; Yamashita, T. FGF-23 Is a Potent Regulator of Vitamin D Metabolism and Phosphate Homeostasis. J. Bone Miner. Res. 2004, 19, 429–435. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, S.L.; Bonjour, J.-P.; Rizzoli, R. Fibroblast Growth Factor-23 Relationship to Dietary Phosphate and Renal Phosphate Handling in Healthy Young Men. J. Clin. Endocrinol. Metab. 2005, 90, 1519–1524. [Google Scholar] [CrossRef] [Green Version]
- Vervloet, M.G.; Van Ittersum, F.J.; Büttler, R.M.; Heijboer, A.; Blankenstein, M.; Ter Wee, P.M. Effects of Dietary Phosphate and Calcium Intake on Fibroblast Growth Factor-23. Clin. J. Am. Soc. Nephrol. 2010, 6, 383–389. [Google Scholar] [CrossRef]
- Masuyama, R.; Stockmans, I.; Torrekens, S.; Van Looveren, R.; Maes, C.; Carmeliet, P.; Bouillon, R.; Carmeliet, G. Vitamin D receptor in chondrocytes promotes osteoclastogenesis and regulates FGF23 production in osteoblasts. J. Clin. Investig. 2006, 116, 3150–3159. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.T.; Lindsay, J.R.; Jain, A.; Kelly, M.H.; Cutler, C.M.; Weinstein, L.S.; Liu, J.; Fedarko, N.S.; Winer, K.K. Fibroblast Growth Factor-23 Is Regulated by 1α,25-Dihydroxyvitamin D. J. Bone Miner. Res. 2005, 20, 1944–1950. [Google Scholar] [CrossRef] [PubMed]
- López, I.; Rodríguez-Ortiz, M.E.; Almadén, Y.; Guerrero, F.; de Oca, A.M.; Pineda, C.; Shalhoub, V.; Rodriguez, M.; Aguilera-Tejero, E. Direct and indirect effects of parathyroid hormone on circulating levels of fibroblast growth factor 23 in vivo. Kidney Int. 2011, 80, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, V.; Martin, A.; Isakova, T.; Spaulding, C.; Qi, L.; Ramirez, V.; Zumbrennen-Bullough, K.B.; Sun, C.C.; Lin, H.Y.; Babitt, J.L.; et al. Inflammation and functional iron deficiency regulate fibroblast growth factor 23 production. Kidney Int. 2016, 89, 135–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bożentowicz-Wikarek, M.; Kocełak, P.; Owczarek, A.; Olszanecka-Glinianowicz, M.; Mossakowska, M.; Skalska, A.; Więcek, A.; Chudek, J. Plasma fibroblast growth factor 23 concentration and iron status. Does the relationship exist in the elderly population? Clin. Biochem. 2015, 48, 431–436. [Google Scholar] [CrossRef]
- Vidal, A.; Rios, R.; Pineda, C.; Lopez, I.; Muñoz-Castañeda, J.R.; Rodriguez, M.; Aguilera-Tejero, E.; Raya, A.I. Direct regulation of fibroblast growth factor 23 by energy intake through mTOR. Sci. Rep. 2020, 10, 1795. [Google Scholar] [CrossRef]
- Scialla, J.J.; Xie, H.; Rahman, M.; Anderson, A.H.; Isakova, T.; Ojo, A.; Zhang, X.; Nessel, L.; Hamano, T.; Grunwald, J.E.; et al. Fibroblast Growth Factor-23 and Cardiovascular Events in CKD. J. Am. Soc. Nephrol. 2014, 25, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Kestenbaum, B.; Sachs, M.C.; Hoofnagle, A.N.; Siscovick, D.S.; Ix, J.H.; Robinson-Cohen, C.; Lima, J.A.; Polak, J.F.; Blondon, M.; Ruzinski, J.; et al. Fibroblast Growth Factor-23 and Cardiovascular Disease in the General Population. Circ. Heart Fail. 2014, 7, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Panwar, B.; Jenny, N.S.; Howard, V.J.; Wadley, V.G.; Muntner, P.; Kissela, B.M.; Judd, S.E.; Gutiérrez, O.M. Fibroblast Growth Factor 23 and Risk of Incident Stroke in Community-Living Adults. Stroke 2015, 46, 322–328. [Google Scholar] [CrossRef]
- Krieger, N.S.; Culbertson, C.D.; Kyker-Snowman, K.; Bushinsky, D.A. Metabolic acidosis increases fibroblast growth factor 23 in neonatal mouse bone. Am. J. Physiol. Physiol. 2012, 303, F431–F436. [Google Scholar] [CrossRef] [Green Version]
- Krieger, N.S.; Bushinsky, D.A. Stimulation of fibroblast growth factor 23 by metabolic acidosis requires osteoblastic intracellular calcium signaling and prostaglandin synthesis. Am. J. Physiol. Physiol. 2017, 313, F882–F886. [Google Scholar] [CrossRef]
- Leibrock, C.B.; Alesutan, I.; Voelkl, J.; Pakladok, T.; Michael, D.; Schleicher, E.; Kamyabi-Moghaddam, Z.; Quintanilla-Martinez, L.; Kuro-O, M.; Lang, F. NH4Cl Treatment Prevents Tissue Calcification in Klotho Deficiency. J. Am. Soc. Nephrol. 2015, 26, 2423–2433. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Melamed, M.L.; Hostetter, T.H.; Bauer, C.; Raff, A.C.; Almudevar, A.L.; LaLonde, A.; Messing, S.; Abramowitz, M.K. Effect of oral sodium bicarbonate on fibroblast growth factor-23 in patients with chronic kidney disease: A pilot study. BMC Nephrol. 2016, 17, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khairallah, P.; Isakova, T.; Asplin, J.; Hamm, L.; Dobre, M.; Rahman, M.; Sharma, K.; Leonard, M.; Miller, E.; Jaar, B.; et al. Acid Load and Phosphorus Homeostasis in CKD. Am. J. Kidney Dis. 2017, 70, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Bär, L.; Feger, M.; Fajol, A.; Klotz, L.-O.; Zeng, S.; Lang, F.; Hocher, B.; Föller, M. Insulin suppresses the production of fibroblast growth factor 23 (FGF23). Proc. Natl. Acad. Sci. 2018, 115, 5804–5809. [Google Scholar] [CrossRef] [Green Version]
- Raya, A.I.; Rios, R.; Pineda, C.; Rodriguez-Ortiz, M.E.; Diez, E.; Almaden, Y.; Muñoz-Castañeda, J.R.; Rodriguez, M.; Aguilera-Tejero, E.; Lopez, I. Energy-dense diets increase FGF23, lead to phosphorus retention and promote vascular calcifications in rats. Sci. Rep. 2016, 6, 36881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanifa, M.A.; Skott, M.; Maltesen, R.G.; Rasmussen, B.S.; Nielsen, S.; Frøkiær, J.; Ring, T.; Wimmer, R. Tissue, urine and blood metabolite signatures of chronic kidney disease in the 5/6 nephrectomy rat model. Metabolomics 2019, 15, 1–16. [Google Scholar] [CrossRef]
- Jara, A.; Felsenfeld, A.J.; Bover, J.; Kleeman, C.R. Chronic metabolic acidosis in azotemic rats on a high-phosphate diet halts the progression of renal disease. Kidney Int. 2000, 58, 1023–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alber, J.; Föller, M. Lactic acid induces fibroblast growth factor 23 (FGF23) production in UMR106 osteoblast-like cells. Mol. Cell. Biochem. 2021, 477, 363–370. [Google Scholar] [CrossRef]
- Lopez, I.; Aguilera-Tejero, E.; Felsenfeld, A.J.; Estepa, J.C.; Rodriguez, M. Direct Effect of Acute Metabolic and Respiratory Acidosis on Parathyroid Hormone Secretion in the Dog. J. Bone Miner. Res. 2002, 17, 1691–1700. [Google Scholar] [CrossRef]
- Krapf, R.; Vetsch, R.; Vetsch, W.; Hulter, H.N. Chronic metabolic acidosis increases the serum concentration of 1,25-dihydroxyvitamin D in humans by stimulating its production rate. Critical role of acidosis-induced renal hypophosphatemia. J. Clin. Investig. 1992, 90, 2456–2463. [Google Scholar] [CrossRef]
- Bushinsky, D.A.; Frick, K.K. The effects of acid on bone. Curr. Opin. Nephrol. Hypertens. 2000, 9, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Lemann, J.; Litzow, J.R.; Lennon, E.J. The effects of chronic acid loads in normal man: Further evidence for the participation of bone mineral in the defense against chronic metabolic acidosis. J. Clin. Investig. 1966, 45, 1608–1614. [Google Scholar] [CrossRef] [Green Version]
- Stauber, A.; Radanovic, T.; Stange, G.; Murer, H.; Wagner, C.A.; Biber, J. Regulation of Intestinal Phosphate Transport II. Metabolic acidosis stimulates Na+-dependent phosphate absorption and expression of the Na+-Pi cotransporter NaPi-IIb in small intestine. Am. J. Physiol. Liver Physiol. 2005, 288, G501–G506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambühl, P.M.; Zajicek, H.K.; Wang, H.; Puttaparthi, K.; Levi, M.; Wilson, W.T.T.A.O.P. Regulation of renal phosphate transport by acute and chronic metabolic acidosis in the rat. Kidney Int. 1998, 53, 1288–1298. [Google Scholar] [CrossRef] [Green Version]
- Nowik, M.; Lecca, M.R.; Velic, A.; Rehrauer, H.; Brändli, A.W.; Wagner, C.A. Genome-wide gene expression profiling reveals renal genes regulated during metabolic acidosis. Physiol. Genom. 2008, 32, 322–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowik, M.; Picard, N.; Stange, G.; Capuano, P.; Tenenhouse, H.S.; Biber, J.; Murer, H.; Wagner, C.A. Renal phosphaturia during metabolic acidosis revisited: Molecular mechanisms for decreased renal phosphate reabsorption. Pflügers Arch.-Eur. J. Physiol. 2008, 457, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, R.C.; Salusky, I.B.; Bowen, R.E.; Freymiller, E.G.; Wesseling-Perry, K. Vitamin D sterols increase FGF23 expression by stimulating osteoblast and osteocyte maturation in CKD bone. Bone 2019, 127, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, H.; Kraut, J.A.; Kurokawa, K. Metabolic Acidosis Suppresses 25-Hydroxyvitamin D3-1α-Hydroxylase in the Rat Kidney. Distinct site and mechanism of action. J. Clin. Investig. 1982, 70, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Lopez, I.; Aguilera-Tejero, E.; Estepa, J.C.; Rodriguez, M.; Felsenfeld, A.J. Role of acidosis-induced increases in calcium on PTH secretion in acute metabolic and respiratory acidosis in the dog. Am. J. Physiol. Metab. 2004, 286, E780–E785. [Google Scholar] [CrossRef]
- Della Guardia, L.; Thomas, M.A.; Cena, H. Insulin Sensitivity and Glucose Homeostasis Can Be Influenced by Metabolic Acid Load. Nutrients 2018, 10, 618. [Google Scholar] [CrossRef] [Green Version]
- Wesson, D.E. Serum bicarbonate as a cardiovascular risk factor: Evolving from ‘non-traditional’? Nephrol. Dial. Transplant. 2020, 35, 1282–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goraya, N.; Munoz-Maldonado, Y.; Simoni, J.; Wesson, D.E. Fruit and Vegetable Treatment of Chronic Kidney Disease-Related Metabolic Acidosis Reduces Cardiovascular Risk Better than Sodium Bicarbonate. Am. J. Nephrol. 2019, 49, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Della Guardia, L.; Roggi, C.; Cena, H. Diet-induced acidosis and alkali supplementation. Int. J. Food Sci. Nutr. 2016, 67, 754–761. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Creatinine (mg/dL) | Urea (mg/dL) | |
|---|---|---|
| Control | 0.66 ± 0.01 | 30.2 ± 1.2 |
| Control+Acid | 0.63 ± 0.01 | 27.0 ± 0.8 |
| 1/2 Nx | 0.80 ± 0.02 a | 39.1 ± 1.8 a |
| 1/2 Nx+Acid | 0.76 ± 0.01 a | 34.1 ± 1.5 |
| 5/6 Nx | 1.12 ± 0.05 a | 70.3 ± 7.7 a |
| 5/6 Nx+Acid | 0.94 ± 0.03 a* | 82.1 ± 10.1 a |
| Bicarbonate (mmol/L) | Anion Gap (mmol/L) | |
|---|---|---|
| Control | 25.9 ± 0.5 | 11.4 ± 0.4 |
| Control+Acid | 24.7 ± 0.7 | 10.6 ± 0.6 |
| 1/2 Nx | 24.9 ± 0.3 | 10.9 ± 0.6 |
| 1/2 Nx+Acid | 23.7 ± 0.6 | 10.6 ± 0.4 |
| 5/6 Nx | 24.2 ± 1.1 | 20.3 ± 2.8 a |
| 5/6 Nx+Acid | 12.6 ± 2.9 a* | 15.6 ± 1.0 * |
| pH | Bicarbonate (mmol/L) | iFGF23 (pg/mL) | |
|---|---|---|---|
| PTX | 7.40 ± 0.01 | 24.6 ± 0.5 | 215.9 ± 25.6 |
| PTX+Acid | 7.27 ± 0.02 * | 16.7 ± 1.0 * | 190.0 ± 31.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidal, A.; Pineda, C.; Raya, A.I.; Rios, R.; Espartero, A.; Muñoz-Castañeda, J.R.; Rodriguez, M.; Aguilera-Tejero, E.; Lopez, I. Oral Acid Load Down-Regulates Fibroblast Growth Factor 23. Nutrients 2022, 14, 1041. https://doi.org/10.3390/nu14051041
Vidal A, Pineda C, Raya AI, Rios R, Espartero A, Muñoz-Castañeda JR, Rodriguez M, Aguilera-Tejero E, Lopez I. Oral Acid Load Down-Regulates Fibroblast Growth Factor 23. Nutrients. 2022; 14(5):1041. https://doi.org/10.3390/nu14051041
Chicago/Turabian StyleVidal, Angela, Carmen Pineda, Ana I. Raya, Rafael Rios, Azahara Espartero, Juan R. Muñoz-Castañeda, Mariano Rodriguez, Escolastico Aguilera-Tejero, and Ignacio Lopez. 2022. "Oral Acid Load Down-Regulates Fibroblast Growth Factor 23" Nutrients 14, no. 5: 1041. https://doi.org/10.3390/nu14051041
APA StyleVidal, A., Pineda, C., Raya, A. I., Rios, R., Espartero, A., Muñoz-Castañeda, J. R., Rodriguez, M., Aguilera-Tejero, E., & Lopez, I. (2022). Oral Acid Load Down-Regulates Fibroblast Growth Factor 23. Nutrients, 14(5), 1041. https://doi.org/10.3390/nu14051041