
A Combined Supplement of Probiotic Strains AP-32, bv-77, and CP-9 Increased Akkermansia mucinphila and Reduced Non-Esterified Fatty Acids and Energy Metabolism in HFD-Induced Obese Rats

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rats
2.2. HFD-Induced Obese Animal Model
2.3. Probiotic Treatments
2.4. Glycemic Level Investigation
2.5. Serum Biochemistry
2.6. 16S rRNA Sequencing of Fecal Sample
2.7. Bioinformatics Analysis
2.8. Statistical Analysis
3. Results
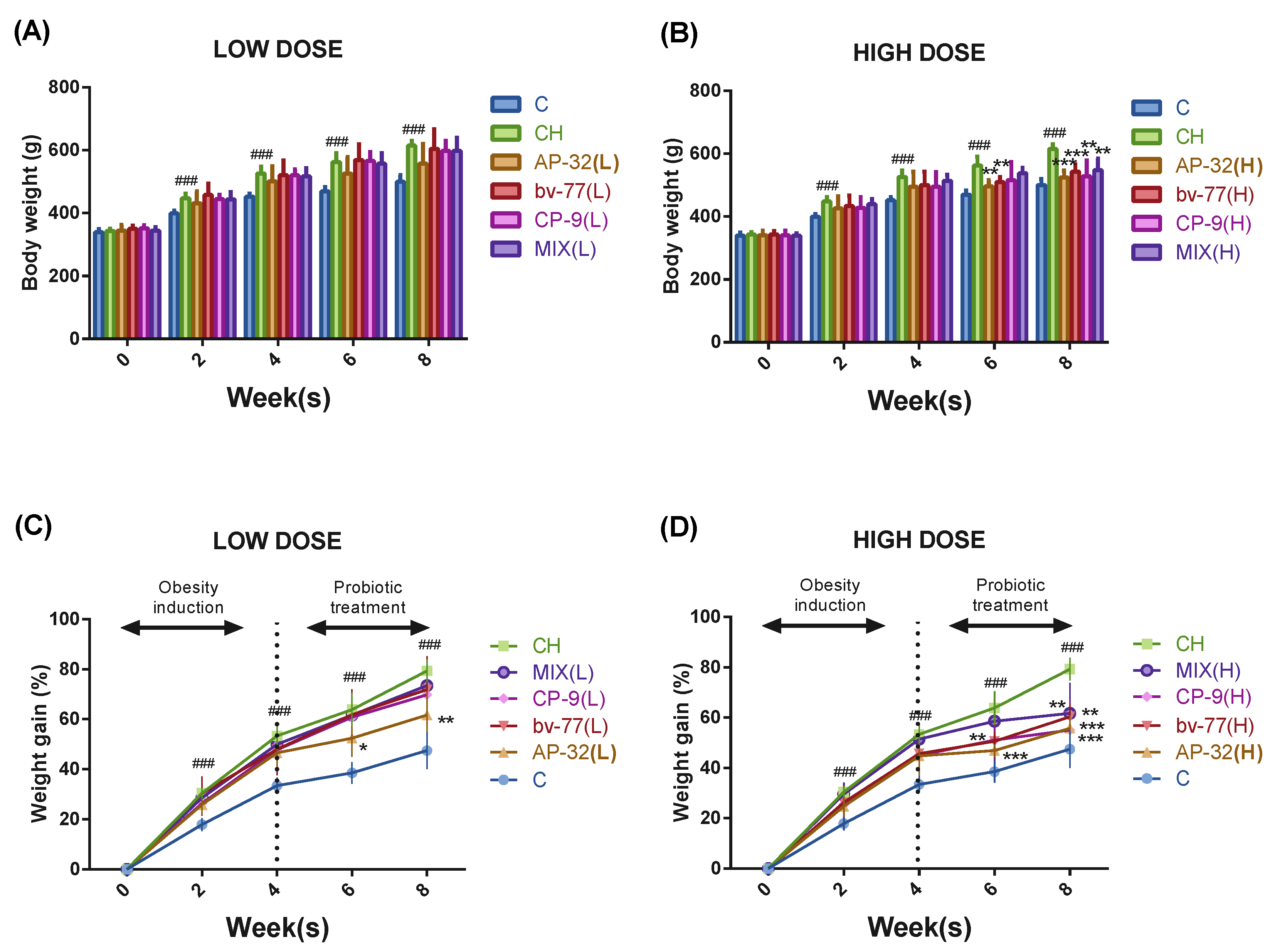
3.1. The Supplement of AP-32, bv-77, and CP-9 Alleviated Weight Gain in Obese Rats
3.2. The Supplement of AP-32, bv-77, and CP-9 Improved Dyslipidemia in Obese Rats
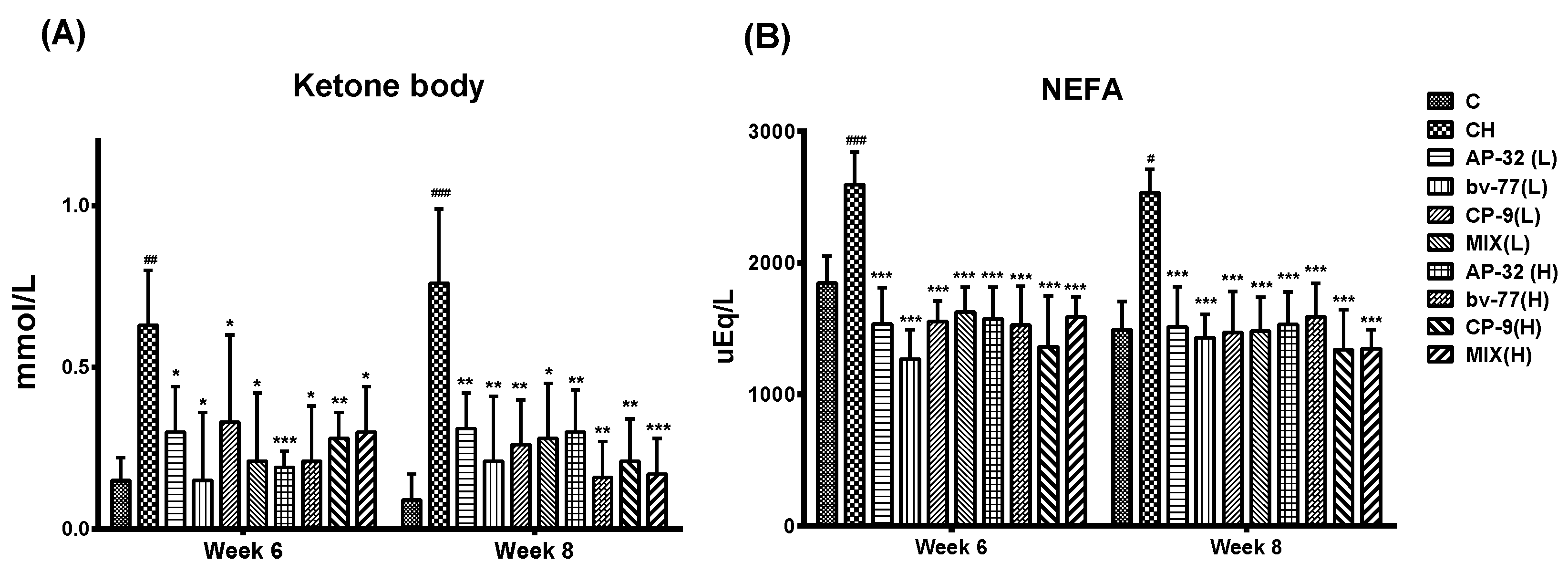
3.3. The Supplement of AP-32, bv-77, and CP-9 Improved Metabolism in Obese Rats
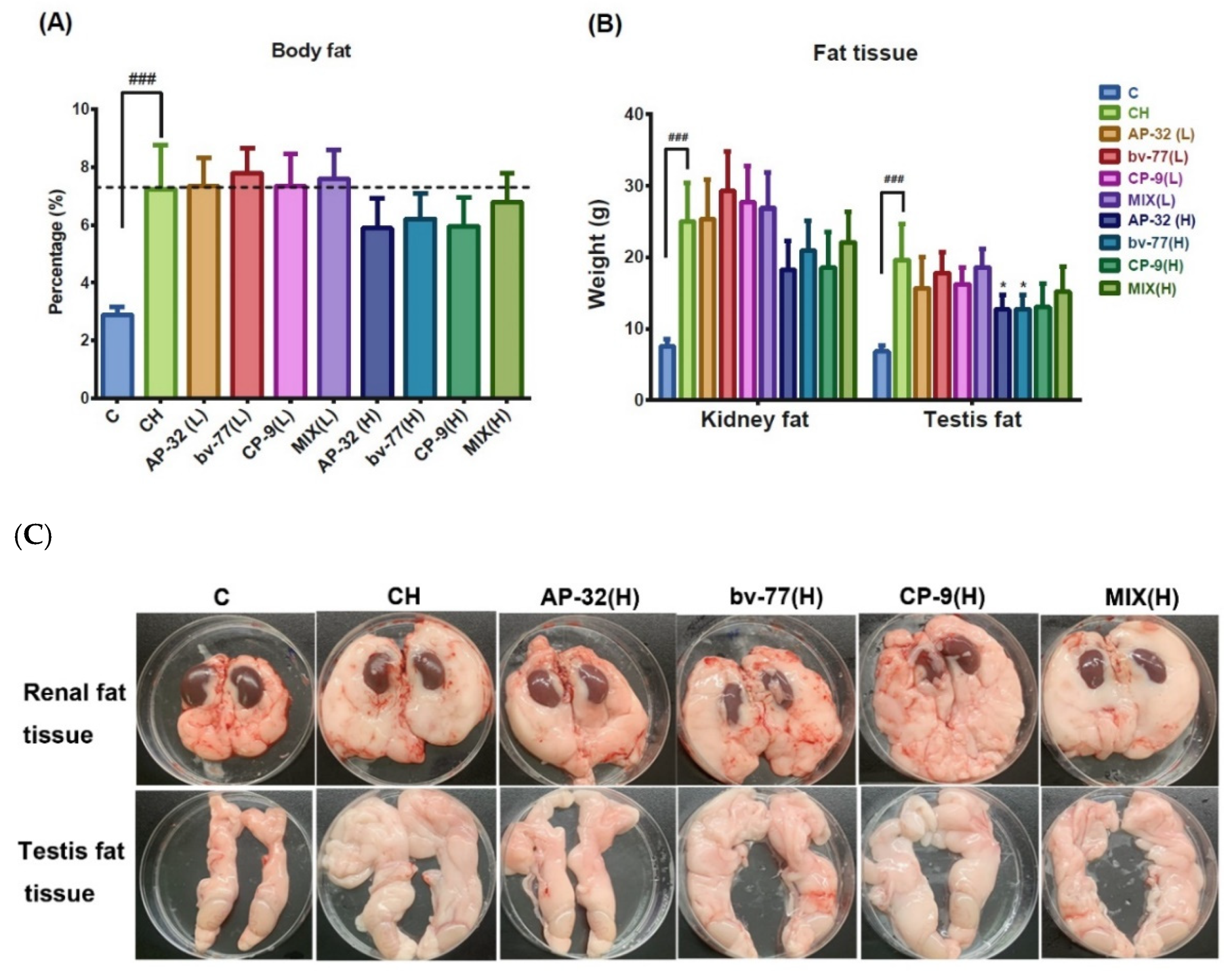
3.4. The Supplement of AP-32, bv-77, and CP-9 Reduced Fat Accumulation in Obese Rats
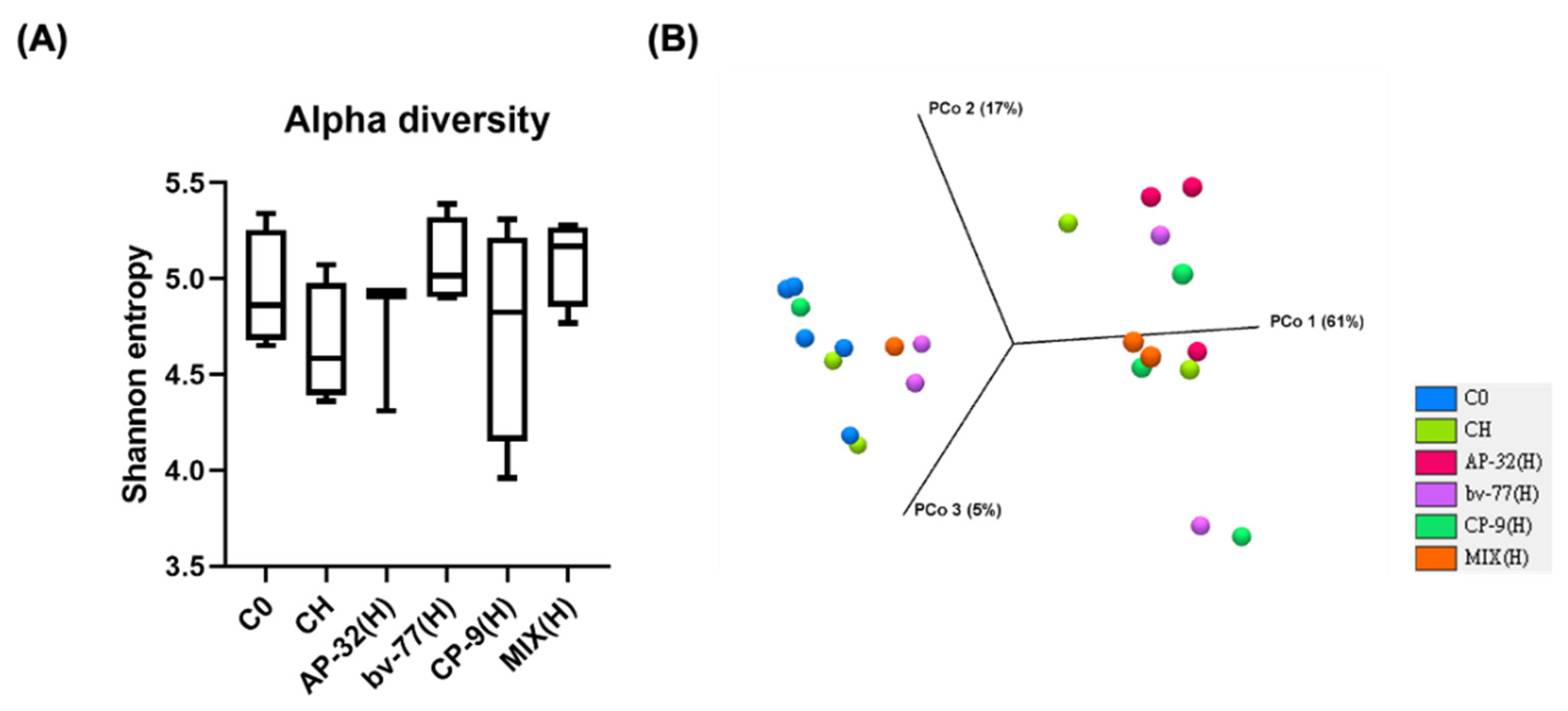
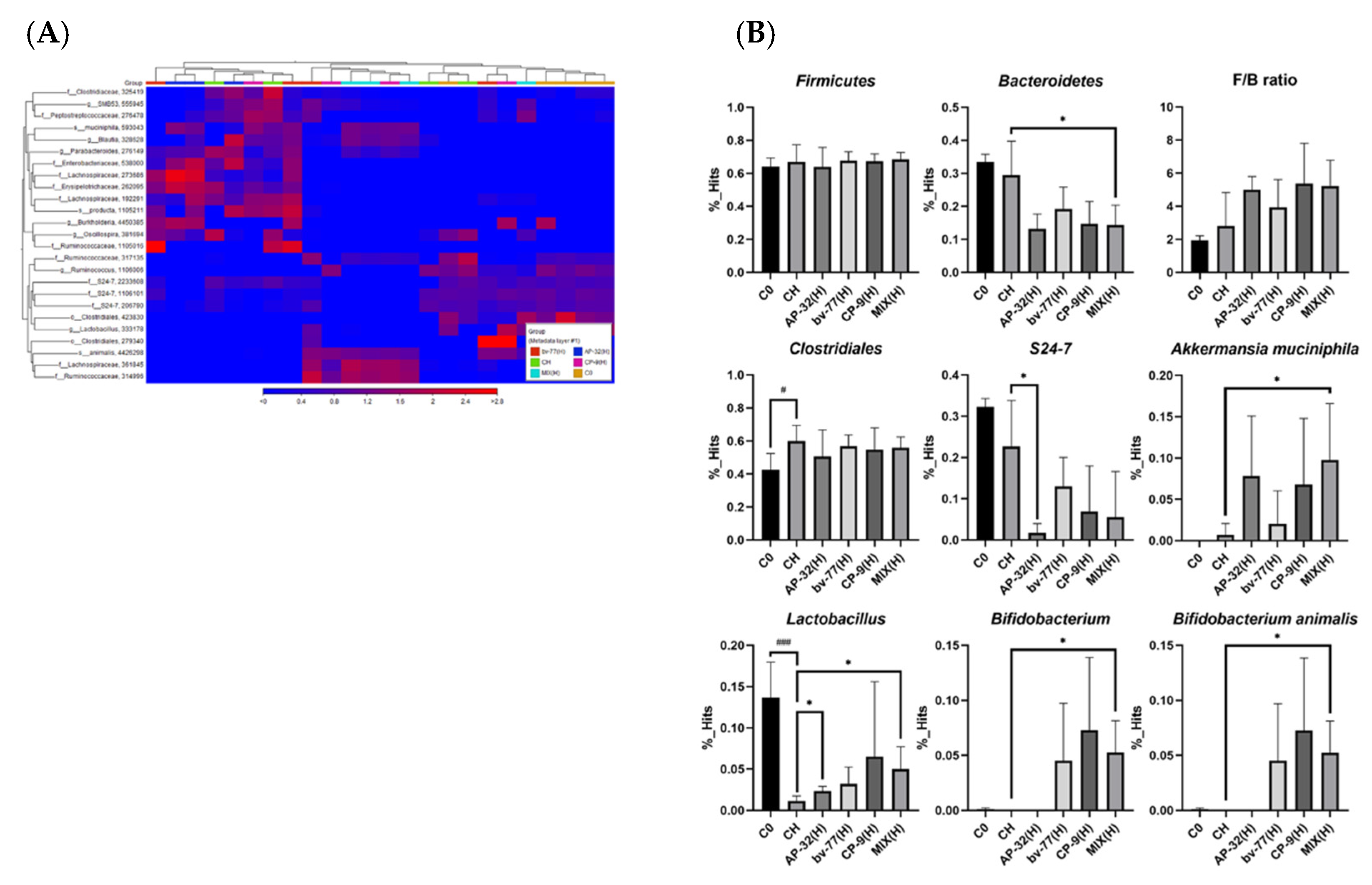
3.5. The Supplement of AP-32, bv-77, and CP-9 Changed Gut Microbiota Composition in Obese Rats
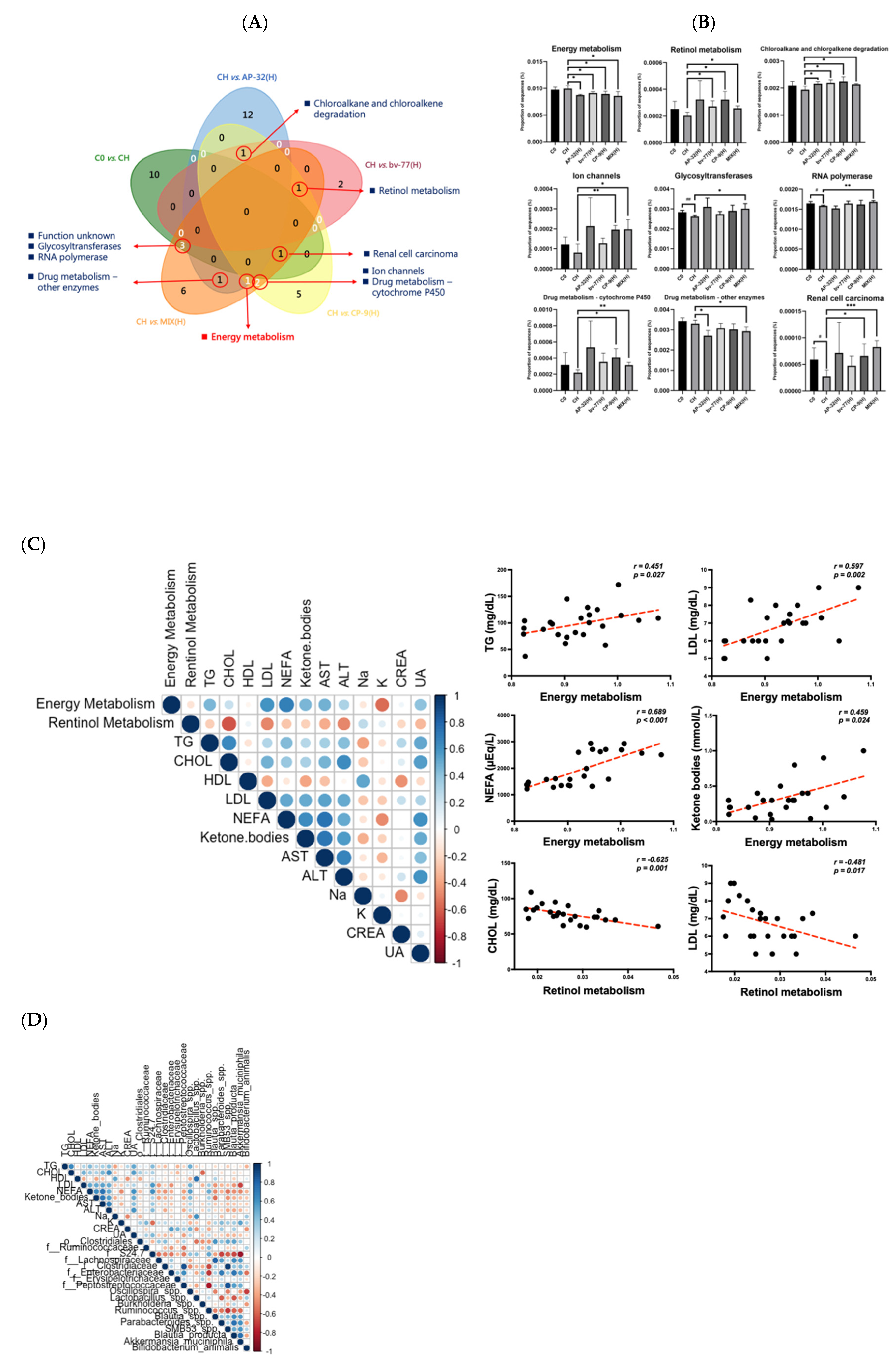
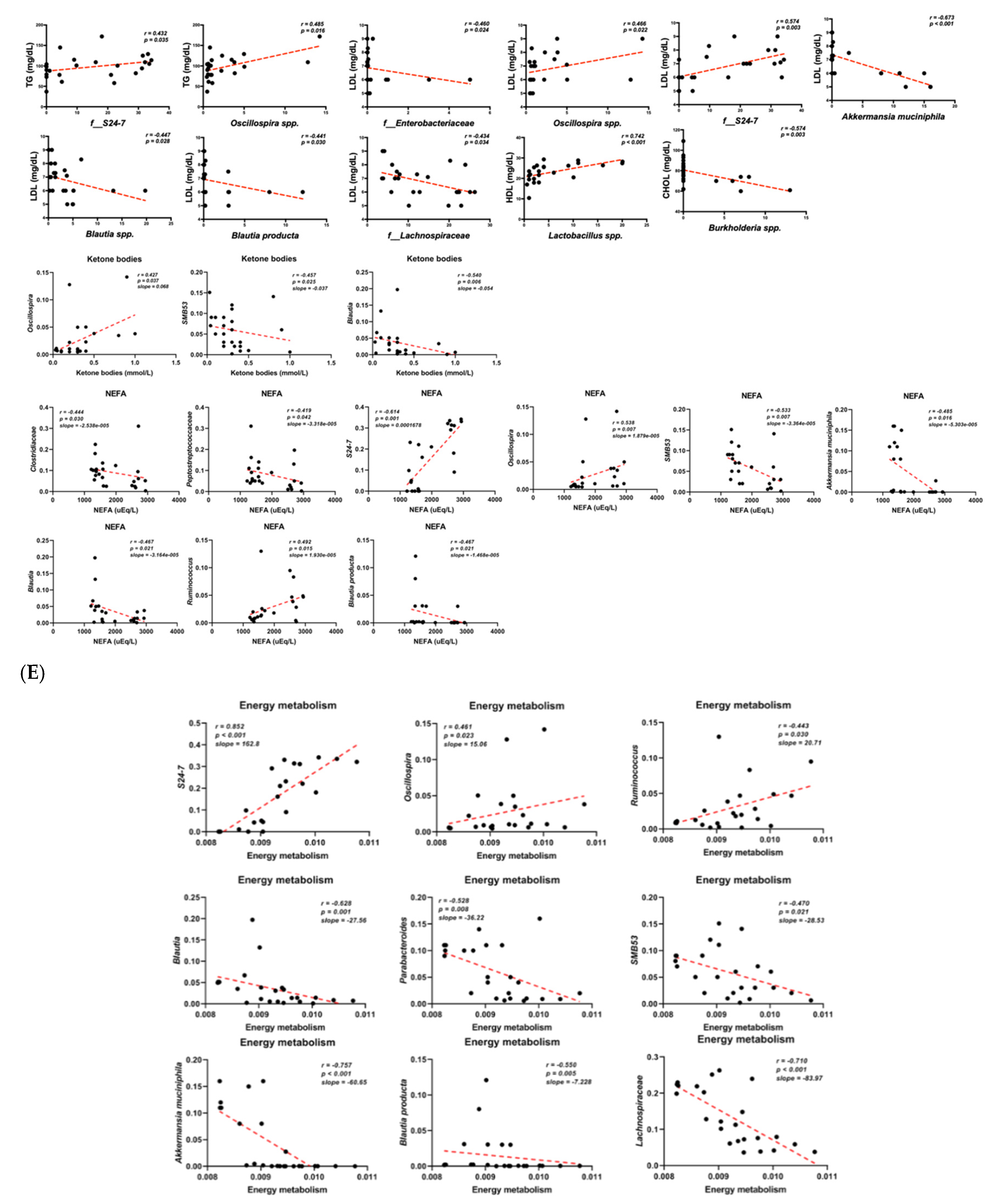
3.6. Functional Pathways of Energy and Retinol Metabolism Were Modulated by Probiotic Intervention
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Expert Consultation. Appropriate body-mass index for Asian populations and its implications for policy and intervention strategies. Lancet 2004, 363, 157–163. [Google Scholar] [CrossRef]
- Grundy, S.M. Obesity, metabolic syndrome, and cardiovascular disease. J. Clin. Endocrinol. Metab. 2004, 89, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Puhl, R.; Brownell, K.D. Bias, discrimination, and obesity. Obes. Res. 2001, 9, 788–805. [Google Scholar] [CrossRef] [PubMed]
- Keith, S.W.; Redden, D.T.; Katzmarzyk, P.T.; Boggiano, M.M.; Hanlon, E.C.; Benca, R.M.; Ruden, D.; Pietrobelli, A.; Barger, J.L.; Fontaine, K.R.; et al. Putative contributors to the secular increase in obesity: Exploring the roads less traveled. Int. J. Obes. 2006, 30, 1585–1594. [Google Scholar] [CrossRef] [Green Version]
- Allen, G.; Safranek, S. FPIN’s clinical inquiries. Secondary causes of obesity. Am. Fam. Phys. 2011, 83, 972–973. [Google Scholar]
- Agha, M.; Agha, R. The rising prevalence of obesity: Part A: Impact on public health. Int. J. Surg. Oncol. 2017, 2, e17. [Google Scholar] [CrossRef] [Green Version]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef]
- Morimoto, C.; Kameda, K.; Tsujita, T.; Okuda, H. Relationships between lipolysis induced by various lipolytic agents and hormone-sensitive lipase in rat fat cells. J. Lipid Res. 2001, 42, 120–127. [Google Scholar] [CrossRef]
- Senior, J.R. Intestinal Absorption of Fats. J. Lipid Res. 1964, 5, 495–521. [Google Scholar] [CrossRef]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Sun, L.; Ma, L.; Ma, Y.; Zhang, F.; Zhao, C.; Nie, Y. Insights into the role of gut microbiota in obesity: Pathogenesis, mechanisms, and therapeutic perspectives. Protein Cell 2018, 9, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; Delzenne, N.M. The role of the gut microbiota in energy metabolism and metabolic disease. Curr. Pharm. Des. 2009, 15, 1546–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbaum, M.; Knight, R.; Leibel, R.L. The gut microbiota in human energy homeostasis and obesity. Trends Endocrinol. Metab. 2015, 26, 493–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.R.; Park, H.-J.; Kang, D.; Chung, H.; Nam, M.H.; Lee, Y.; Park, J.-H.; Lee, H.-Y. A protective mechanism of probiotic Lactobacillus against hepatic steatosis via reducing host intestinal fatty acid absorption. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; Qin, H. Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 2019, 60, 175–184. [Google Scholar] [CrossRef]
- Karpe, F.; Dickmann, J.R.; Frayn, K.N. Fatty acids, obesity, and insulin resistance: Time for a reevaluation. Diabetes 2011, 60, 2441–2449. [Google Scholar] [CrossRef] [Green Version]
- Post, A.; Garcia, E.; van den Berg, E.H.; Flores-Guerrero, J.L.; Gruppen, E.G.; Groothof, D.; Westenbrink, B.D.; Connelly, M.A.; Bakker, S.J.L.; Dullaart, R.P.F. Nonalcoholic fatty liver disease, circulating ketone bodies and all-cause mortality in a general population-based cohort. Eur. J. Clin. Investig. 2021, 51, e13627. [Google Scholar] [CrossRef]
- Donnelly, K.L.; Smith, C.I.; Schwarzenberg, S.J.; Jessurun, J.; Boldt, M.D.; Parks, E.J. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J. Clin. Investig. 2005, 115, 1343–1351. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Li, Y.-L.; Zhang, X.; Wang, K.-B.; Huang, J.-A.; Liu, Z.-H.; Zhu, M.-Z. Polyphenols from Fu Brick Tea Reduce Obesity via Modulation of Gut Microbiota and Gut Microbiota-Related Intestinal Oxidative Stress and Barrier Function. J. Agric. Food Chem. 2021, 69, 14530–14543. [Google Scholar] [CrossRef]
- Pan, X.; Kaminga, A.C.; Liu, A.; Wen, S.W.; Luo, M.; Luo, J. Gut Microbiota, Glucose, Lipid, and Water-Electrolyte Metabolism in Children with Nonalcoholic Fatty Liver Disease. Front. Cell. Infect. Microbiol. 2021, 11, 683743. [Google Scholar] [CrossRef] [PubMed]
- Dehghanbanadaki, H.; Aazami, H.; Keshavarz Azizi Raftar, S.; Ashrafian, F.; Ejtahed, H.S.; Hashemi, E.; Hoseini Tavassol, Z.; Ahmadi Badi, S.; Siadat, S.D. Global scientific output trend for Akkermansia muciniphila research: A bibliometric and scientometric analysis. BMC Med. Inform. Decis. Mak. 2020, 20, 291. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.M.; Lu, G.; Greene, E.L. Vascular effects of non-esterified fatty acids: Implications for the cardiovascular risk factor cluster. Prostaglandins Leukot. Essent. Fat. Acids 1999, 60, 411–420. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Kang, H.W.; Bhimidi, G.R.; Odom, D.P.; Brun, P.-J.; Fernandez, M.-L.; McGrane, M.M. Altered lipid catabolism in the vitamin A deficient liver. Mol. Cell. Endocrinol. 2007, 271, 18–27. [Google Scholar] [CrossRef]
- Ross, A.C. Vitamin A and retinoic acid in T cell-related immunity. Am. J. Clin. Nutr. 2012, 96, 1166S–1172S. [Google Scholar] [CrossRef]
- Shabana; Shahid, S.U.; Irfan, U. The gut microbiota and its potential role in obesity. Future Microbiol. 2018, 13, 589–603. [Google Scholar] [CrossRef]
- Mazloom, K.; Siddiqi, I.; Covasa, M. Probiotics: How Effective Are They in the Fight against Obesity? Nutrients 2019, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.-S.; An, Y.; Tsai, Y.-C.; Chen, Y.-C.; Chuang, C.-J.; Zeng, C.-T.; Wang, C.-T.; An-Erl King, V. Potential of probiotic strains to modulate the inflammatory responses of epithelial and immune cells in vitro. New Microbiol. 2013, 36, 167–179. [Google Scholar]
- Hsieh, P.-S.; Tsai, Y.-C.; Chen, Y.-C.; Teh, S.-F.; Ou, C.-M.; King, V.A. Eradication of Helicobacter pylori infection by the probiotic strains Lactobacillus johnsonii MH-68 and L. salivarius ssp. salicinius AP-32. Helicobacter 2012, 17, 466–477. [Google Scholar] [CrossRef]
- Ou, C.-C.; Lin, S.-L.; Tsai, J.-J.; Lin, M.-Y. Heat-killed lactic acid bacteria enhance immunomodulatory potential by skewing the immune response toward Th1 polarization. J. Food Sci. 2011, 76, M260–M267. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Hsieh, P.S.; Ho, H.H.; Hsieh, S.H.; Kuo, Y.W.; Yang, S.F.; Lin, C.W. Antibacterial activity of viable and heat-killed probiotic strains against oral pathogens. Lett. Appl. Microbiol. 2020, 70, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.-S.; Ho, H.-H.; Hsieh, S.-H.; Kuo, Y.-W.; Tseng, H.-Y.; Kao, H.-F.; Wang, J.-Y. Lactobacillus salivarius AP-32 and Lactobacillus reuteri GL-104 decrease glycemic levels and attenuate diabetes-mediated liver and kidney injury in db/db mice. BMJ Open Diabetes Res. Care 2020, 8, e001028. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.-S.; Ho, H.-H.; Tsao, S.-P.; Hsieh, S.-H.; Lin, W.-Y.; Chen, J.-F.; Kuo, Y.-W.; Tsai, S.-Y.; Huang, H.-Y. Multi-strain probiotic supplement attenuates streptozotocin-induced type-2 diabetes by reducing inflammation and beta-cell death in rats. PLoS ONE 2021, 16, e0251646. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Carbajal, A.; Pizano-Zárate, M.L.; Hernández-Quiroz, F.; Ortiz-Luna, G.F.; Morales-Hernández, R.M.; De Sales-Millán, A.; Hernández-Trejo, M.; García-Vite, A.; Beltrán-Lagunes, L.; Hoyo-Vadillo, C.; et al. Characterization of the Gut Microbiota of Individuals at Different T2D Stages Reveals a Complex Relationship with the Host. Microorganisms 2020, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Michael, D.R.; Davies, T.S.; Moss, J.W.E.; Calvente, D.L.; Ramji, D.P.; Marchesi, J.R.; Pechlivanis, A.; Plummer, S.F.; Hughes, T.R. The anti-cholesterolaemic effect of a consortium of probiotics: An acute study in C57BL/6J mice. Sci. Rep. 2017, 7, 2883. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.; Lee, Y.; Kang, H.J.; Ju, J.; Ji, Y.; Park, H.; Park, H.; Lee, H.; Holzapfel, W.H. Two putative probiotic strains improve diet-induced hypercholesterolemia through modulating intestinal cholesterol uptake and hepatic cholesterol efflux. J. Appl. Microbiol. 2022, 132, 562–570. [Google Scholar] [CrossRef]
- Çaglar, E.; Kuscu, O.O.; Selvi Kuvvetli, S.; Kavaloglu Cildir, S.; Sandalli, N.; Twetman, S. Short-term effect of ice-cream containing Bifidobacterium lactis Bb-12 on the number of salivary mutans streptococci and lactobacilli. Acta Odontol. Scand. 2008, 66, 154–158. [Google Scholar] [CrossRef]
- Rizzardini, G.; Eskesen, D.; Calder, P.C.; Capetti, A.; Jespersen, L.; Clerici, M. Evaluation of the immune benefits of two probiotic strains Bifidobacterium animalis ssp. lactis, BB-12(R) and Lactobacillus paracasei ssp. paracasei, L. casei 431(R) in an influenza vaccination model: A randomised, double-blind, placebo-controlled study. Br. J. Nutr. 2012, 107, 876–884. [Google Scholar] [CrossRef] [Green Version]
- Million, M.; Maraninchi, M.; Henry, M.; Armougom, F.; Richet, H.; Carrieri, P.; Valero, R.; Raccah, D.; Vialettes, B.; Raoult, D. Obesity-associated gut microbiota is enriched in Lactobacillus reuteri and depleted in Bifidobacterium animalis and Methanobrevibacter smithii. Int. J. Obes. 2012, 36, 817–825. [Google Scholar] [CrossRef] [Green Version]
- Carson, J.A.S.; Lichtenstein, A.H.; Anderson, C.A.M.; Appel, L.J.; Kris-Etherton, P.M.; Meyer, K.A.; Petersen, K.; Polonsky, T.; Van Horn, L.; on behalf of the American Heart Association Nutrition Committee of the Council on Lifestyle and Cardiometabolic Health; et al. Dietary Cholesterol and Cardiovascular Risk: A Science Advisory from the American Heart Association. Circulation 2020, 141, e39–e53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.-R.; Kim, Y.-J.; Park, D.-Y.; Jung, U.-J.; Jeon, S.-M.; Ahn, Y.-T.; Huh, C.-S.; McGregor, R.; Choi, M.S. Probiotics, L. plantarum and L. curvatus in combination alter hepatic lipid metabolism and suppress diet-induced obesity. Obesity 2013, 21, 2571–2578. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Puri, S.; Kurpad, A.V. Potential of Probiotics in Hypercholesterolemia: A Review of In Vitro and In Vivo Findings. Altern. Ther. Health Med. 2018, 24, 36–43. [Google Scholar] [PubMed]
- Hsieh, P.-S.; Chen, C.-W.; Kuo, Y.-W.; Ho, H.-H. Lactobacillus spp. reduces ethanol-induced liver oxidative stress and inflammation in a mouse model of alcoholic steatohepatitis. Exp. Ther. Med. 2021, 21, 188. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Bergman, R.N. Non-esterified fatty acids and the liver: Why is insulin secreted into the portal vein? Diabetologia 2000, 43, 946–952. [Google Scholar] [CrossRef] [Green Version]
- Cai, D.; Yuan, M.; Frantz, D.F.; Melendez, P.A.; Hansen, L.; Lee, J.; Shoelson, S.E. Local and systemic insulin resistance resulting from hepatic activation of IKK-beta and NF-kappaB. Nat. Med. 2005, 11, 183–190. [Google Scholar] [CrossRef]
- Afzali, A.; Weiss, N.S.; Boyko, E.J.; Ioannou, G.N. Association between serum uric acid level and chronic liver disease in the United States. Hepatology 2010, 52, 578–589. [Google Scholar] [CrossRef]
- Kovesdy, C.P.; Furth, S.L.; Zoccali, C.; on behalf of the World Kidney Day Steering Committee. Obesity and Kidney Disease: Hidden Consequences of the Epidemic. Can. J. Kidney Health Dis. 2017, 4, 2054358117698669. [Google Scholar] [CrossRef] [Green Version]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Ouwerkerk, J.P.; van der Ark, K.C.H.; Davids, M.; Claassens, N.J.; Finestra, T.R.; de Vos, W.M.; Belzer, C. Adaptation of Akkermansia muciniphila to the Oxic-Anoxic Interface of the Mucus Layer. Appl. Environ. Microbiol. 2016, 82, 6983–6993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gophna, U.; Konikoff, T.; Nielsen, H.B. Oscillospira and related bacteria—From metagenomic species to metabolic features. Environ. Microbiol. 2017, 19, 835–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-R.; Zheng, H.-M.; Zhang, G.-X.; Chen, F.-L.; Chen, L.-D.; Yang, Z.-C. High Oscillospira abundance indicates constipation and low BMI in the Guangdong Gut Microbiome Project. Sci. Rep. 2020, 10, 9364. [Google Scholar] [CrossRef] [PubMed]
- Yun, K.E.; Kim, J.; Kim, M.-H.; Park, E.; Kim, H.-L.; Chang, Y.; Ryu, S.; Kim, H.-N. Major Lipids, Apolipoproteins, and Alterations of Gut Microbiota. J. Clin. Med. 2020, 9, 1589. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.-L.; Tang, X.-H.; Li, X.; Yu, H.-B.; Xie, Z.-G.; Liu, X.-Y.; Zhou, Z.-G. Effect of high-fat or high-glucose diet on obesity and visceral adipose tissue in mice. Acta Acad. Med. Sin. 2014, 36, 614–619. [Google Scholar] [CrossRef]
- Ji, M.; Du, H.; Xu, Y. Structural and metabolic performance of p-cresol producing microbiota in different carbon sources. Food Res. Int. 2020, 132, 109049. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Gálvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Pereira, F.C.; Wasmund, K.; Cobankovic, I.; Jehmlich, N.; Herbold, C.W.; Lee, K.S.; Sziranyi, B.; Vesely, C.; Decker, T.; Stocker, R.; et al. Rational design of a microbial consortium of mucosal sugar utilizers reduces Clostridiodes difficile colonization. Nat. Commun. 2020, 11, 5104. [Google Scholar] [CrossRef]
- Allaband, C.; Lingaraju, A.; Martino, C.; Russell, B.; Tripathi, A.; Poulsen, O.; Dantas Machado, A.C.; Zhou, D.; Xue, J.; Elijah, E.; et al. Intermittent Hypoxia and Hypercapnia Alter Diurnal Rhythms of Luminal Gut Microbiome and Metabolome. mSystems 2021, 6, e0011621. [Google Scholar] [CrossRef]
- Liu, S.; Qin, P.; Wang, J. High-Fat Diet Alters the Intestinal Microbiota in Streptozotocin-Induced Type 2 Diabetic Mice. Microorganisms 2019, 7, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanueva-Millán, M.J.; Pérez-Matute, P.; Oteo, J.A. Gut microbiota: A key player in health and disease. A review focused on obesity. J. Physiol. Biochem. 2015, 71, 509–525. [Google Scholar] [CrossRef] [PubMed]
- DiBaise, J.K.; Zhang, H.; Crowell, M.D.; Krajmalnik-Brown, R.; Decker, G.A.; Rittmann, B.E. Gut microbiota and its possible relationship with obesity. Mayo Clin. Proc. 2008, 83, 460–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leatham, M.P.; Banerjee, S.; Autieri, S.M.; Mercado-Lubo, R.; Conway, T.; Cohen, P.S. Precolonized human commensal Escherichia coli strains serve as a barrier to E. coli O157:H7 growth in the streptomycin-treated mouse intestine. Infect. Immun. 2009, 77, 2876–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadooka, Y.; Sato, M.; Ogawa, A.; Miyoshi, M.; Uenishi, H.; Ogawa, H.; Ikuyama, K.; Kagoshima, M.; Tsuchida, T. Effect of Lactobacillus gasseri SBT2055 in fermented milk on abdominal adiposity in adults in a randomised controlled trial. Br. J. Nutr. 2013, 110, 1696–1703. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week 6 | C | CH | AP-32(L) | bv-77(L) | CP-9(L) | MIX(L) | AP-32(H) | bv-77(H) | CP-9(H) | MIX(H) |
|---|---|---|---|---|---|---|---|---|---|---|
| Glucose (mg/dL) | 102.4 ± 4.16 | 109.5 ± 6.35 | 100.8 ± 9.65 | 111.4 ± 5.13 | 116.4 ± 1.67 | 113.6 ± 7.86 | 109.2 ± 5.72 | 116.4 ± 6.02 | 115.6 ± 4.93 | 106.8 ± 4.15 |
| Cholesterol (mg/dL) | 80.50 ± 8.46 | 85.67 ± 12.52 | 71.00 ± 24.49 | 70.50 ± 18.28 | 89.33 ± 15.64 | 85.50 ± 19.04 | 73.00 ± 8.27 | 63.50 * ± 12.14 | 77.33 ± 13.16 | 71.33 ± 9.69 |
| Triacylglycerol (mg/dL) | 99.50 ± 9.05 | 122.50 # ± 18.36 | 99.00 ± 25.97 | 91.83 ± 37.72 | 101.50 ± 32.38 | 105.80 ± 22.29 | 96.00 * ± 17.10 | 89.67 ± 32.16 | 91.17 ** ± 15.52 | 80.67 ** ± 19.34 |
| LDL (mg/dL) | 6.63 ± 0.75 | 8.52 # ± 0.61 | 7.90 ± 0.67 | 7.55 ± 1.30 | 7.30 ± 1.22 | 7.22 * ± 1.10 | 6.90 * ± 1.19 | 7.72 ± 0.32 | 6.00 *** ± 0.18 | 6.42 *** ± 0.49 |
| HDL (mg/dL) | 25.28 ± 2.35 | 15.82 ## ± 2.55 | 19.53 ± 5.32 | 21.33 * ± 4.77 | 21.75 * ± 4.97 | 23.63 ** ± 4.50 | 21.63 ** ± 3.41 | 22.83 ** ± 4.77 | 22.00 ** ± 2.26 | 20.48 * ± 3.42 |
| AST (U/L) | 103.83 ± 16.64 | 142.33 # ± 29.51 | 112.33 * ± 12.32 | 128.40 * ± 15.01 | 110.50 * ± 10.99 | 110.00 ** ± 5.01 | 107.67 ** ± 7.80 | 116.33 ** ± 4.27 | 109.80 ** ± 7.98 | 99.83 ** ± 13.36 |
| ALT (U/L) | 29.83 ± 5.56 | 40.83 ## ± 4.17 | 29.00 ** ± 6.10 | 22.00 ** ± 3.58 | 29.00 *** ± 6.99 | 25.67 *** ± 1.86 | 30.17 *** ± 2.04 | 21.17 *** ± 5.60 | 24.40 *** ± 3.65 | 25.00 *** ± 3.74 |
| Uric acid (mg/dL) | 1.10 ± 0.24 | 2.08 ### ± 0.21 | 1.52 ** ± 0.26 | 1.62 * ± 0.30 | 1.67 ** ± 0.10 | 1.55 * ± 0.30 | 1.35 *** ± 0.27 | 1.60 ** ± 0.30 | 1.80 * ± 0.23 | 1.55 ** ± 0.32 |
| Creatinine (mg/dL) | 0.25 ± 0.03 | 0.31 # ± 0.04 | 0.26 ± 0.05 | 0.30 ± 0.07 | 0.32 ± 0.03 | 0.25 ** ± 0.02 | 0.25 * ± 0.03 | 0.24 ** ± 0.03 | 0.27 ± 0.02 | 0.26 ± 0.03 |
| Week 8 | C | CH | AP-32(L) | bv-77(L) | CP-9(L) | MIX(L) | AP-32(H) | bv-77(H) | CP-9(H) | MIX(H) |
|---|---|---|---|---|---|---|---|---|---|---|
| Glucose (mg/dL) | 105.6 ± 6.47 | 108.75 ± 5.91 | 106.6 ± 5.41 | 105.00 ± 5.96 | 116.6 ± 10.92 | 109.2 ± 6.30 | 99.2 * ± 4.44 | 116.6 ± 6.06 | 107.6 ± 4.62 | 107.2 ± 6.02 |
| Cholesterol (mg/dL) | 79.33 ± 6.12 | 94.50 # ± 10.43 | 77.67 * ± 10.48 | 75.67 ± 21.81 | 79.50 ± 15.18 | 72.17 * ± 15.87 | 71.83 ** ± 10.68 | 64.17 * ± 16.07 | 71.33 ** ± 7.09 | 75.00 ** ± 7.48 |
| Triacylglycerol (mg/dL) | 98.33 ± 15.16 | 129.83 # ± 22.60 | 89.33 * ± 22.47 | 104.42 ± 16.82 | 101.00 * ± 21.56 | 89.17 ** ± 18.39 | 82.50 ** ± 11.79 | 81.83 * ± 29.60 | 89.75 * ± 37.21 | 80.33 ** ± 18.68 |
| LDL (mg/dL) | 6.90 ± 1.19 | 8.13 ± 0.80 | 7.60 ± 0.89 | 7.63 ± 1.71 | 6.87 * ± 0.75 | 6.10 *** ± 0.68 | 6.52 * ± 1.26 | 6.72 * ± 0.92 | 6.22 *** ± 0.83 | 6.00 ** ± 0.89 |
| HDL (mg/dL) | 25.60 ± 2.30 | 16.22 ## ± 4.08 | 19.47 ± 4.26 | 24.57 ** ± 2.86 | 24.63 ** ± 3.24 | 25.77 ** ± 3.59 | 22.50 * ± 3.14 | 23.07 ** ± 2.93 | 23.62 ** ± 3.47 | 25.18 ** ± 2.69 |
| AST (U/L) | 118.80 ± 25.06 | 137.17 ± 19.65 | 105.00 ** ± 10.92 | 109.20 ** ± 16.58 | 107.83 * ± 17.52 | 84.67 *** ± 7.79 | 102.33 ** ± 9.33 | 88.00 *** ± 6.42 | 91.50 *** ± 10.01 | 89.83 *** ± 8.23 |
| ALT (U/L) | 33.33 ± 7.03 | 43.33 # ± 7.50 | 25.50 *** ± 4.09 | 22.83 *** ± 3.92 | 27.00 ** ± 5.40 | 28.00 ** ± 3.85 | 29.17 ** ± 4.58 | 25.83 *** ± 3.31 | 25.00 *** ± 5.55 | 28.83 ** ± 5.19 |
| Uric acid (mg/dL) | 1.37 ± 0.61 | 2.32 ## ± 0.32 | 1.38 *** ± 0.32 | 1.60 * ± 0.57 | 1.63 *** ± 0.18 | 1.77 * ± 0.47 | 1.42 ** ± 0.40 | 1.67 * ± 0.55 | 1.38 *** ± 0.30 | 1.63 ** ± 0.41 |
| Creatinine (mg/dL) | 0.31 ± 0.02 | 0.35 ± 0.07 | 0.25 * ± 0.02 | 0.26 * ± 0.04 | 0.27 * ± 0.05 | 0.26 * ± 0.04 | 0.26 * ± 0.02 | 0.24 ** ± 0.02 | 0.22 ** ± 0.04 | 0.24 ** ± 0.04 |
| Na+ (mmol/L) | 151.17 ± 1.72 | 151.17 ± 1.94 | 151.33 ± 1.37 | 153.17 ± 0.98 | 150.80 ± 1.64 | 153.33 ± 1.21 | 151.50 ± 1.64 | 152.50 ± 2.88 | 152.50 ± 0.58 | 151.40 ± 1.14 |
| K+ (mmol/L) | 6.08 ± 0.26 | 6.20 ± 0.48 | 6.45 ± 0.52 | 6.45 ± 0.51 | 6.77 ± 0.48 | 6.38 ± 0.24 | 6.70 ± 0.70 | 6.27 ± 0.41 | 6.68 ± 0.90 | 6.55 ± 0.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, C.-A.; Huang, C.-H.; Ho, H.-H.; Chen, J.-F.; Kuo, Y.-W.; Lin, J.-H.; Tsai, S.-Y.; Tsai, H.-Y.; Yeh, Y.-T. A Combined Supplement of Probiotic Strains AP-32, bv-77, and CP-9 Increased Akkermansia mucinphila and Reduced Non-Esterified Fatty Acids and Energy Metabolism in HFD-Induced Obese Rats. Nutrients 2022, 14, 527. https://doi.org/10.3390/nu14030527
Liao C-A, Huang C-H, Ho H-H, Chen J-F, Kuo Y-W, Lin J-H, Tsai S-Y, Tsai H-Y, Yeh Y-T. A Combined Supplement of Probiotic Strains AP-32, bv-77, and CP-9 Increased Akkermansia mucinphila and Reduced Non-Esterified Fatty Acids and Energy Metabolism in HFD-Induced Obese Rats. Nutrients. 2022; 14(3):527. https://doi.org/10.3390/nu14030527
Chicago/Turabian StyleLiao, Chorng-An, Cheng-Hsieh Huang, Hsieh-Hsun Ho, Jui-Fen Chen, Yi-Wei Kuo, Jia-Hung Lin, Shin-Yu Tsai, Hui-Yun Tsai, and Yao-Tsung Yeh. 2022. "A Combined Supplement of Probiotic Strains AP-32, bv-77, and CP-9 Increased Akkermansia mucinphila and Reduced Non-Esterified Fatty Acids and Energy Metabolism in HFD-Induced Obese Rats" Nutrients 14, no. 3: 527. https://doi.org/10.3390/nu14030527
APA StyleLiao, C.-A., Huang, C.-H., Ho, H.-H., Chen, J.-F., Kuo, Y.-W., Lin, J.-H., Tsai, S.-Y., Tsai, H.-Y., & Yeh, Y.-T. (2022). A Combined Supplement of Probiotic Strains AP-32, bv-77, and CP-9 Increased Akkermansia mucinphila and Reduced Non-Esterified Fatty Acids and Energy Metabolism in HFD-Induced Obese Rats. Nutrients, 14(3), 527. https://doi.org/10.3390/nu14030527