
Perinatal Treatment with Leptin, but Not Celastrol, Protects from Metabolically Obese, Normal-Weight Phenotype in Rats




Abstract
:1. Introduction
2. Materials and Methods
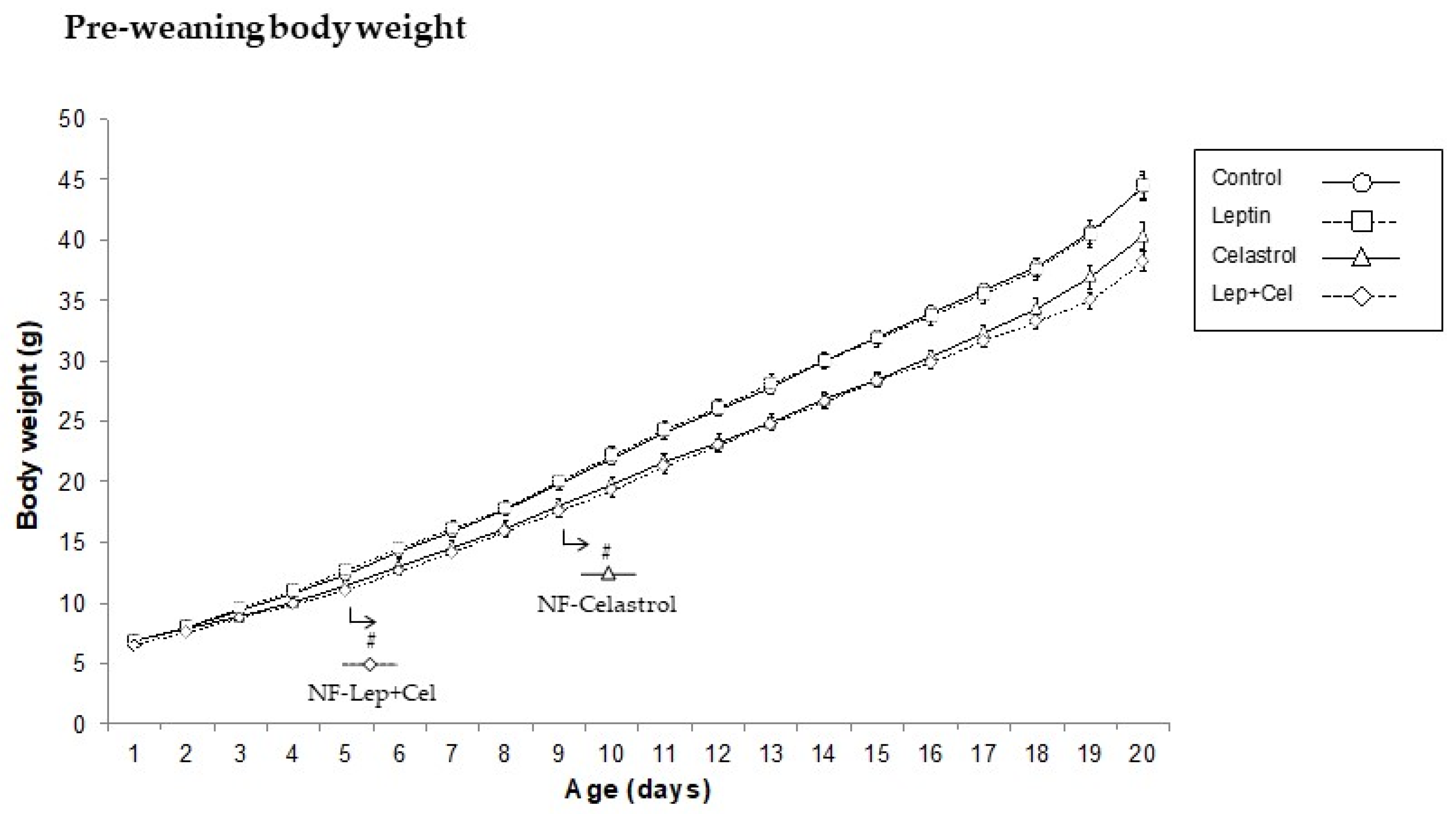
2.1. Animal and Experimental Design
2.2. Adiposity Index
2.3. Circulating Parameters (Glucose, Insulin, Leptin)
2.4. Homeostatic Model Assessment of Insulin Resistance (HOMA-IR)
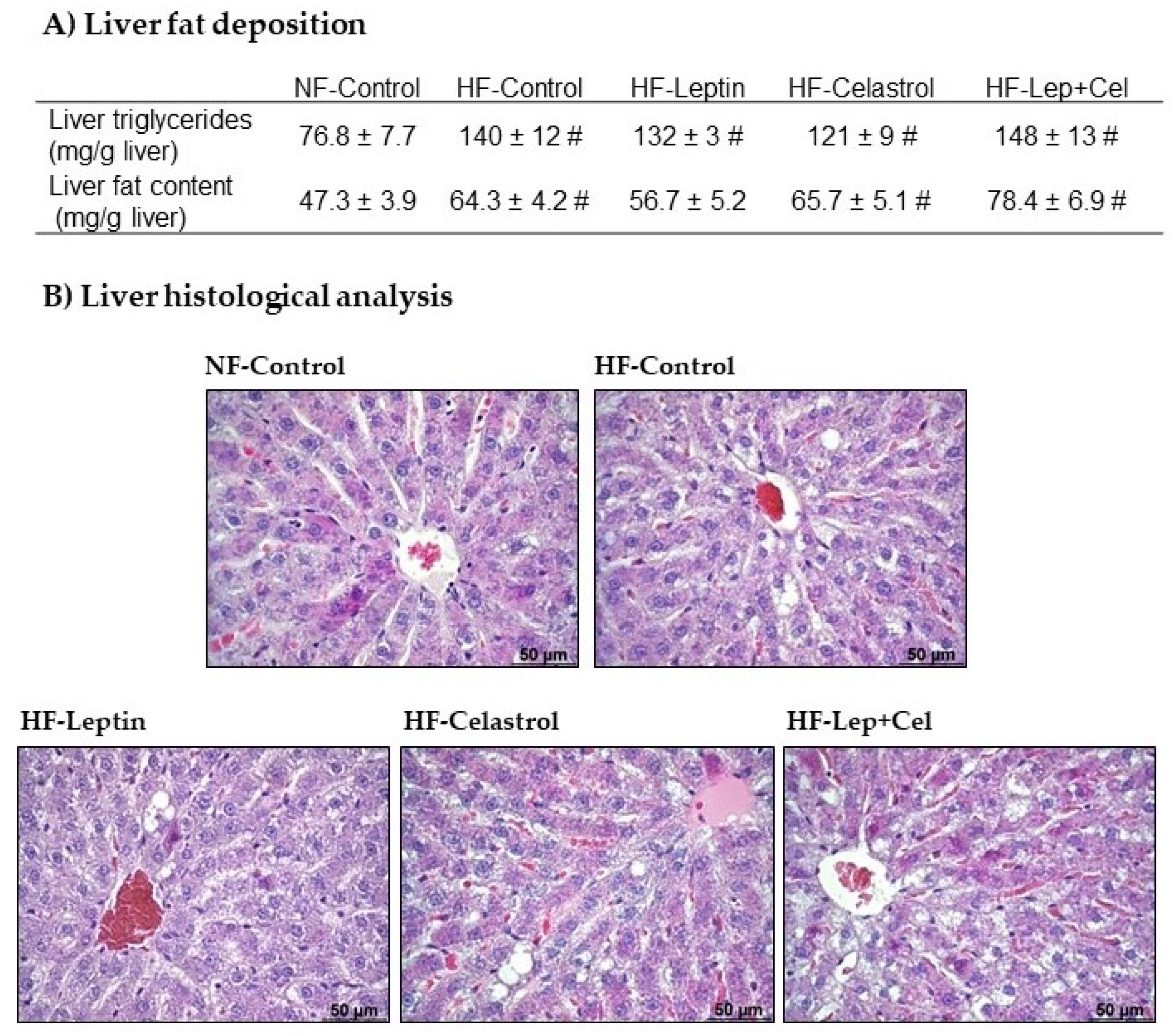
2.5. Quantification of Liver Lipid Levels
2.6. Liver Histological Analysis
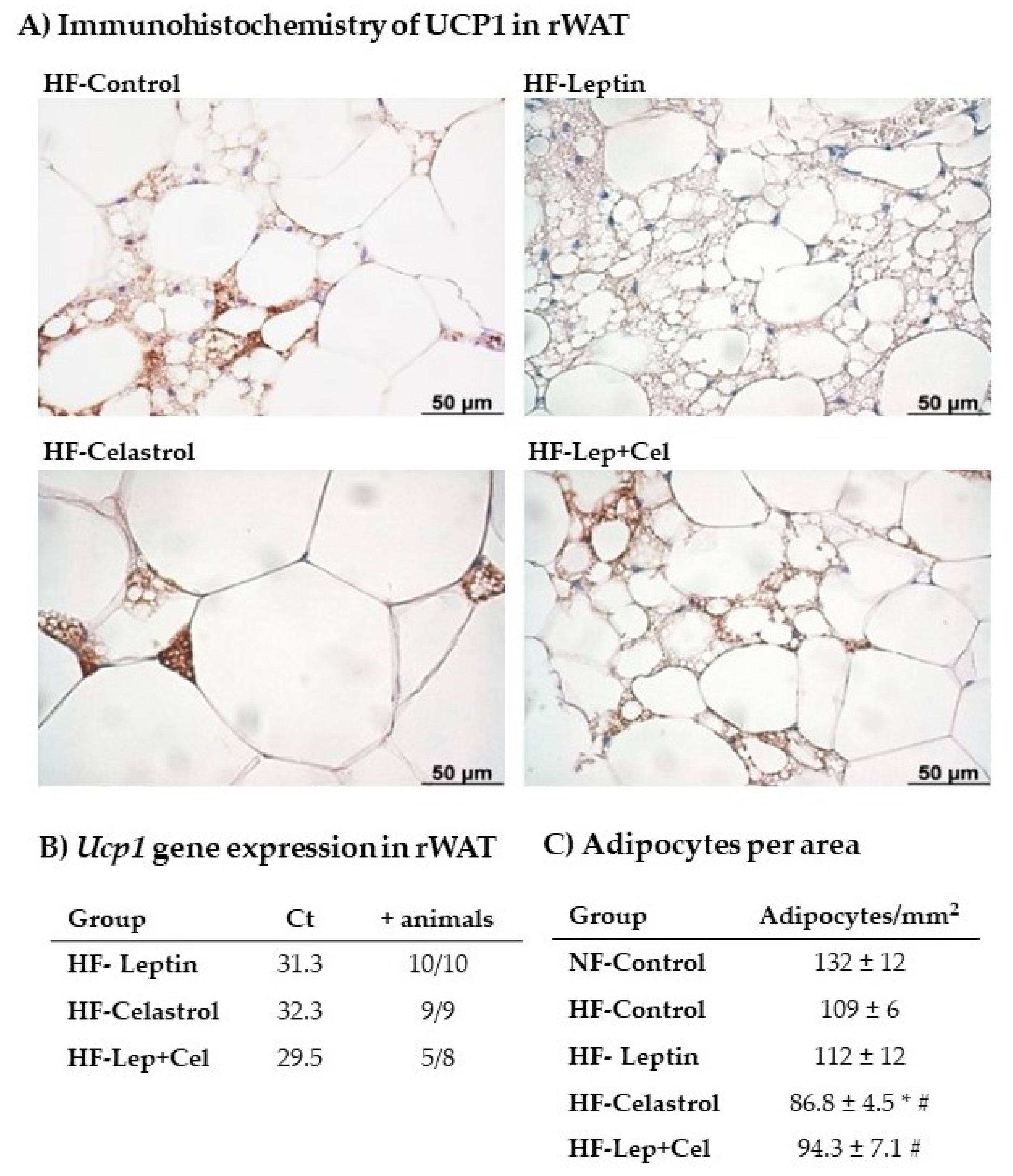
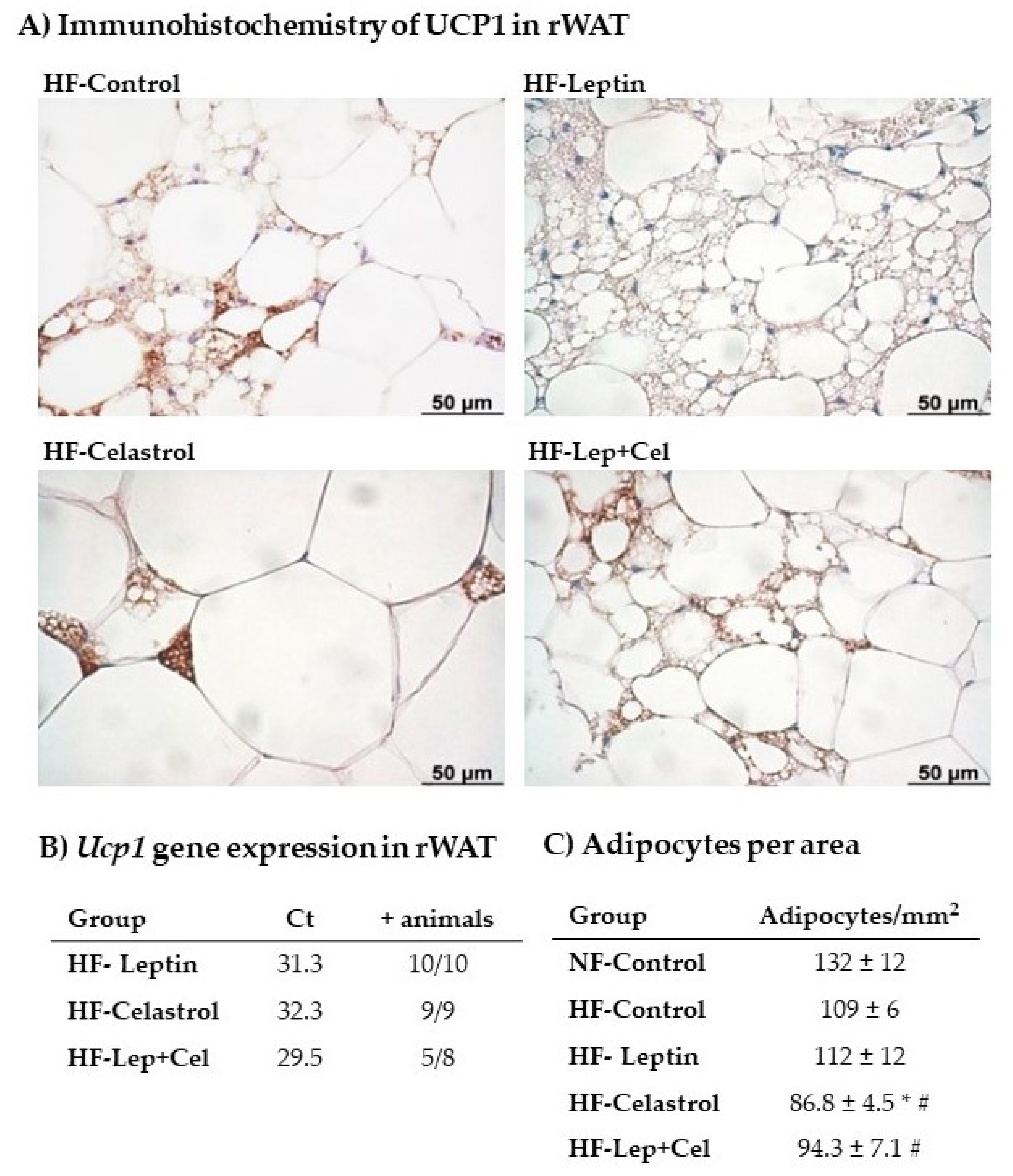
2.7. Histological and Immunohistochemistry Analysis of UCP1 in Retroperitoneal White Adipose Tissue
2.8. Total RNA Isolation
2.9. Real-Time Reverse Transcriptase Polymerase Chain Reaction (RT-qPCR)
2.10. Statistical Analysis
3. Results
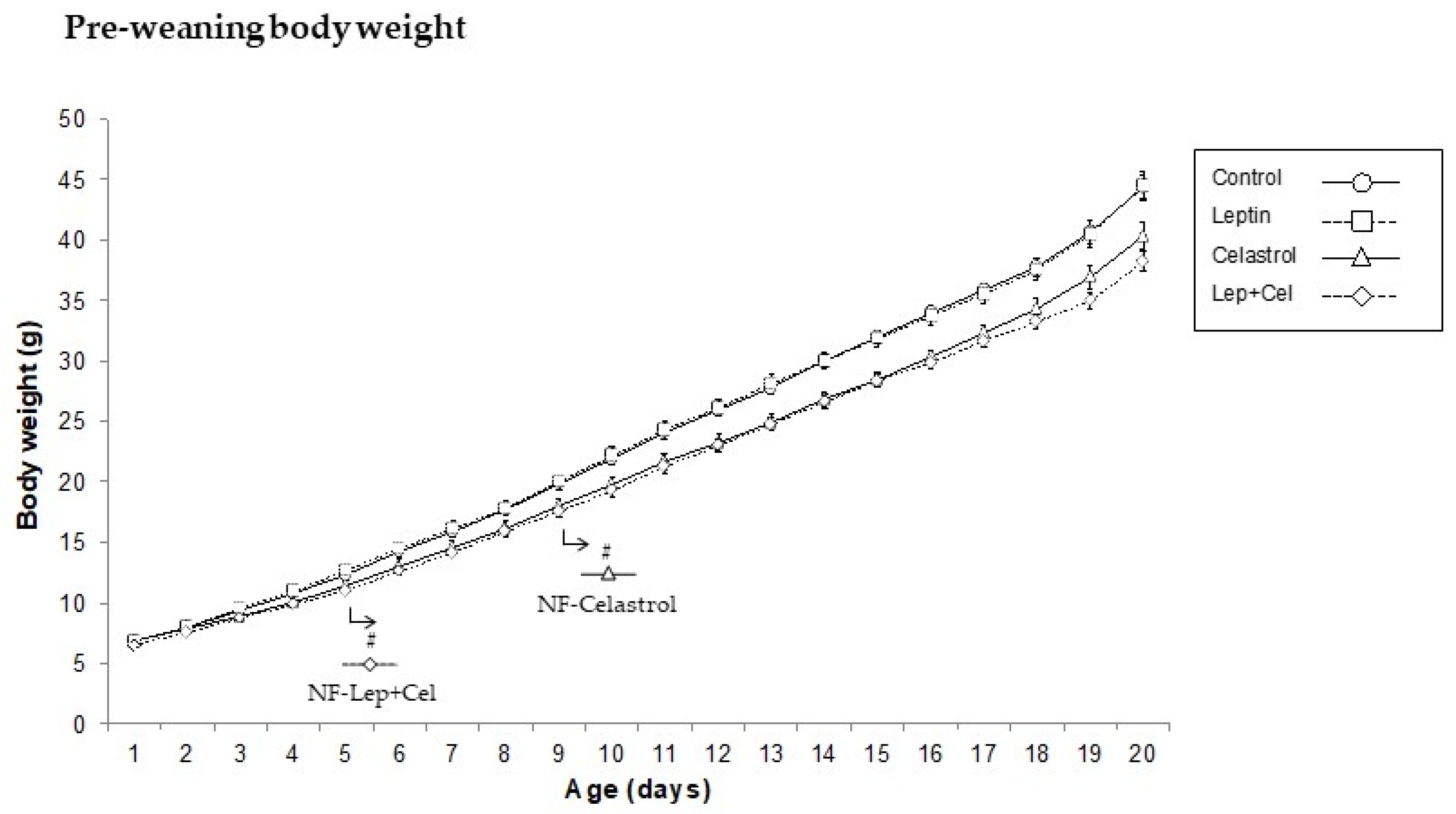
3.1. Body Weight, Adiposity, and Circulating Parameters
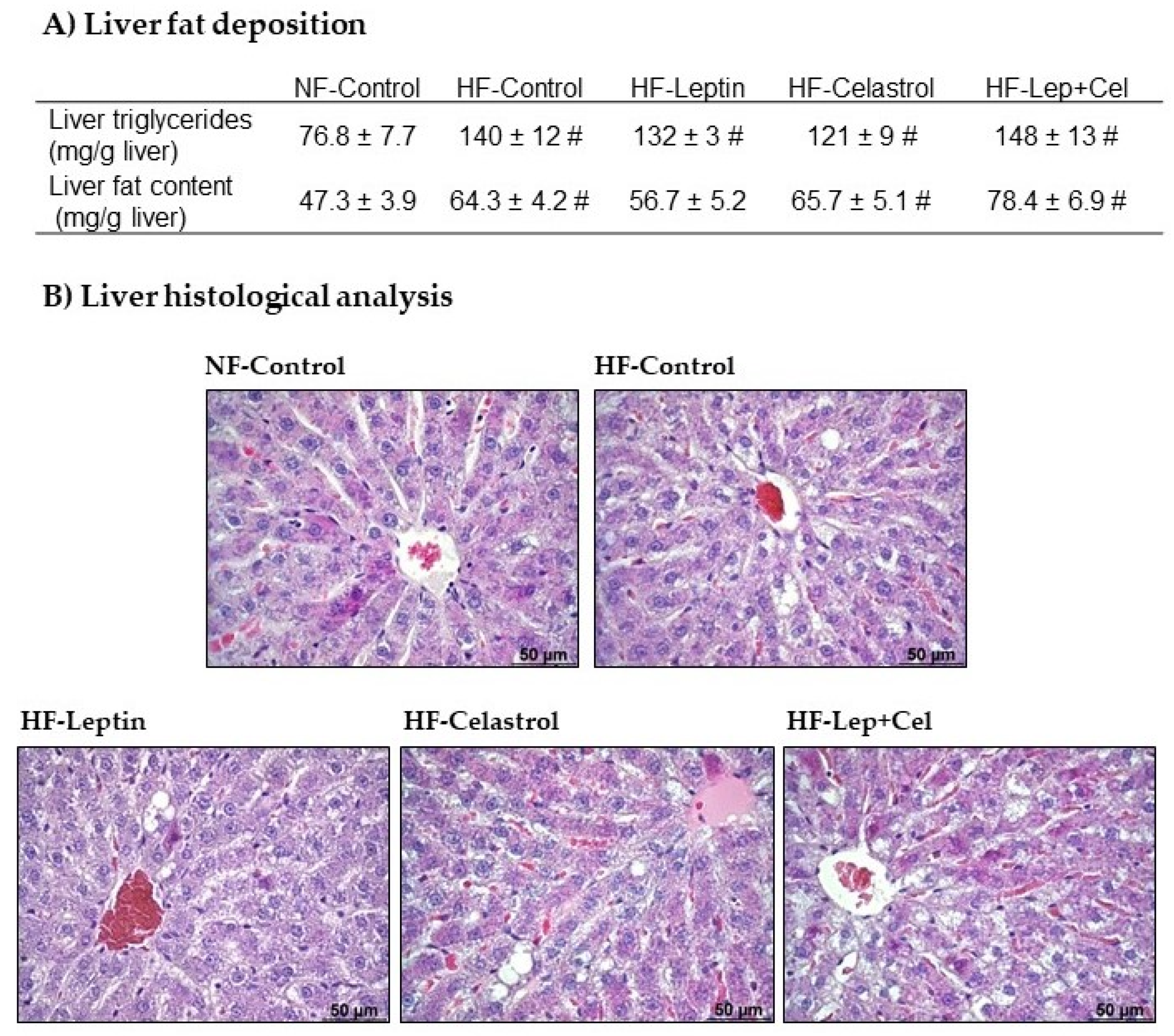
3.2. Liver Fat Deposition and Histological Analysis
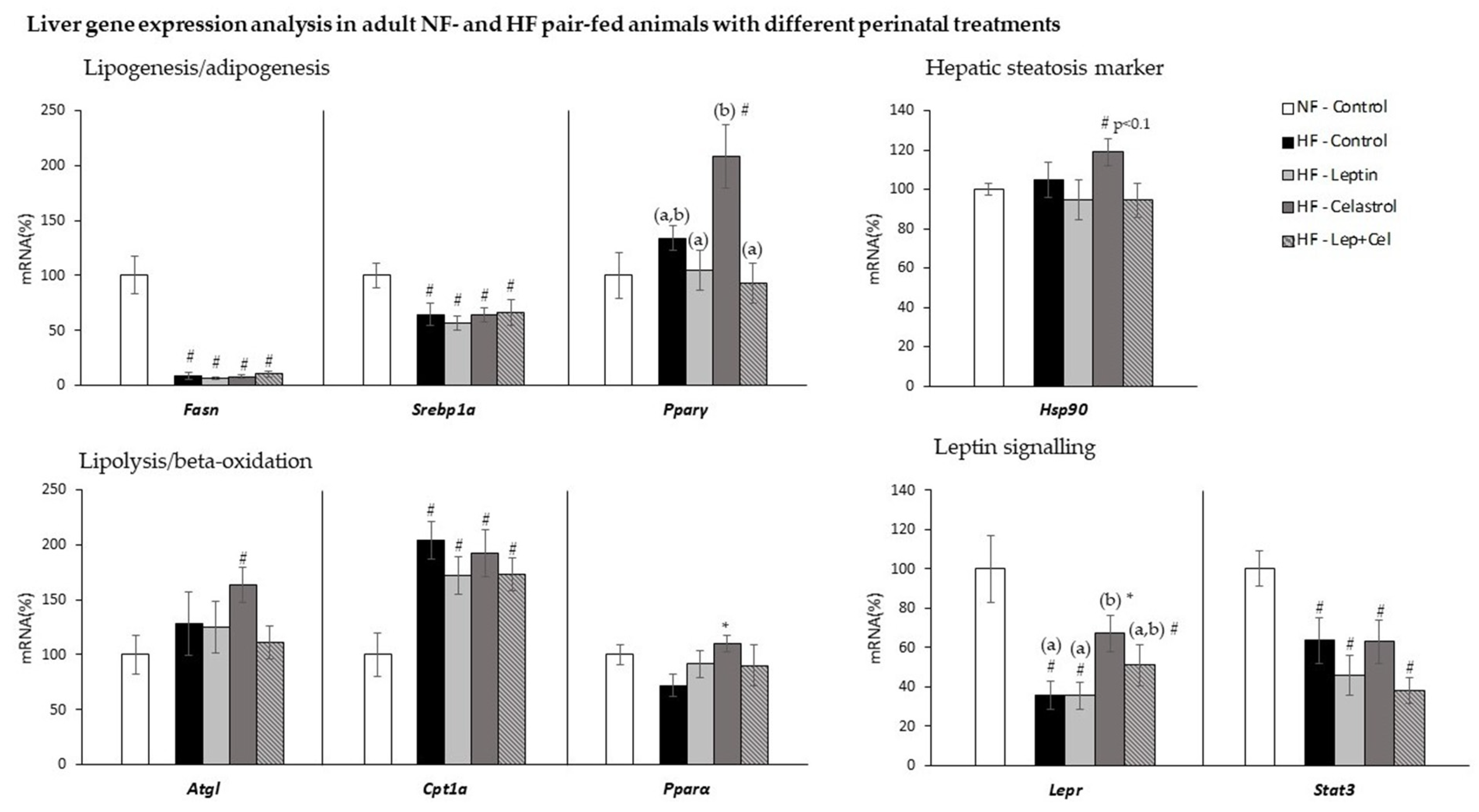
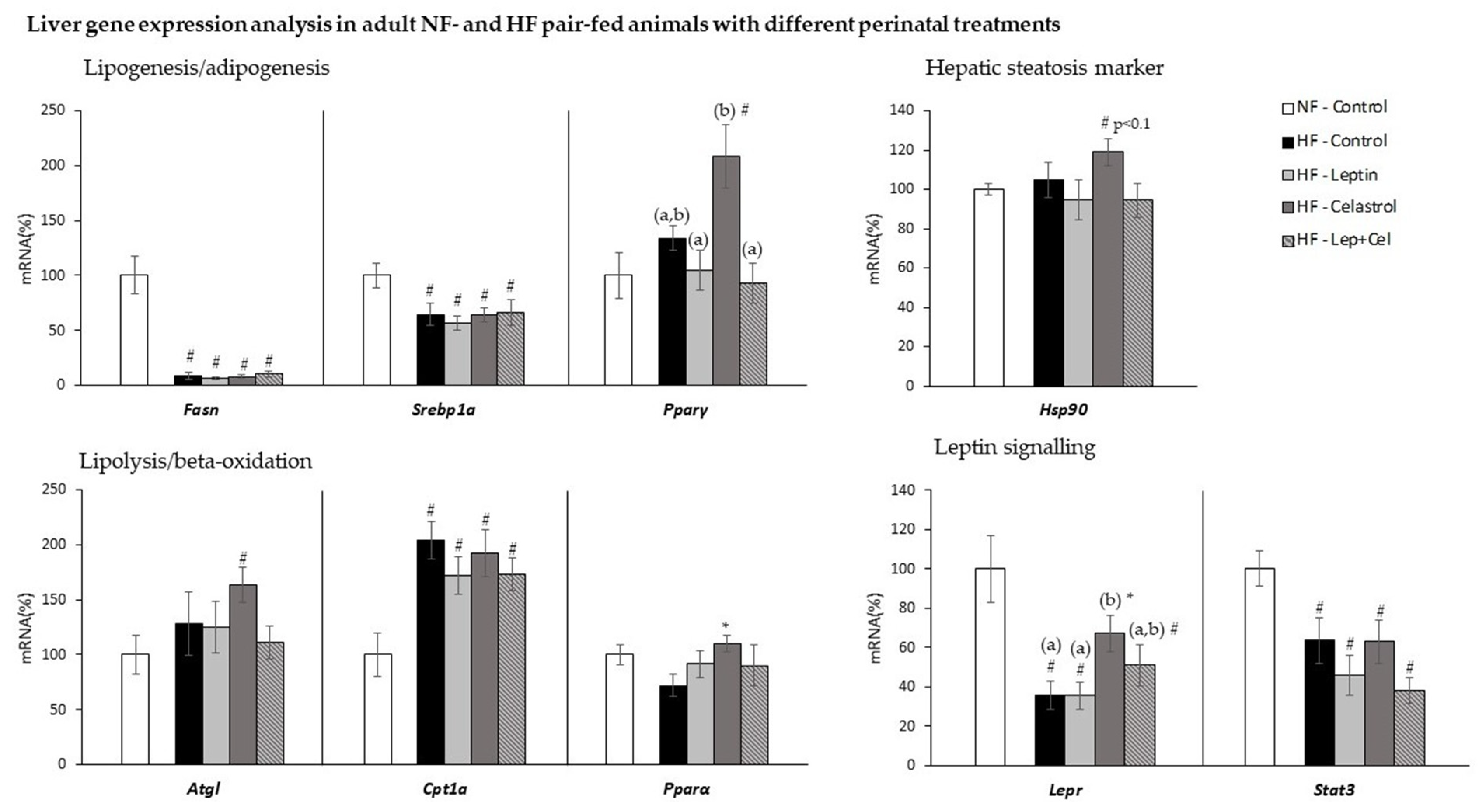
3.3. Liver Gene Expression Analysis
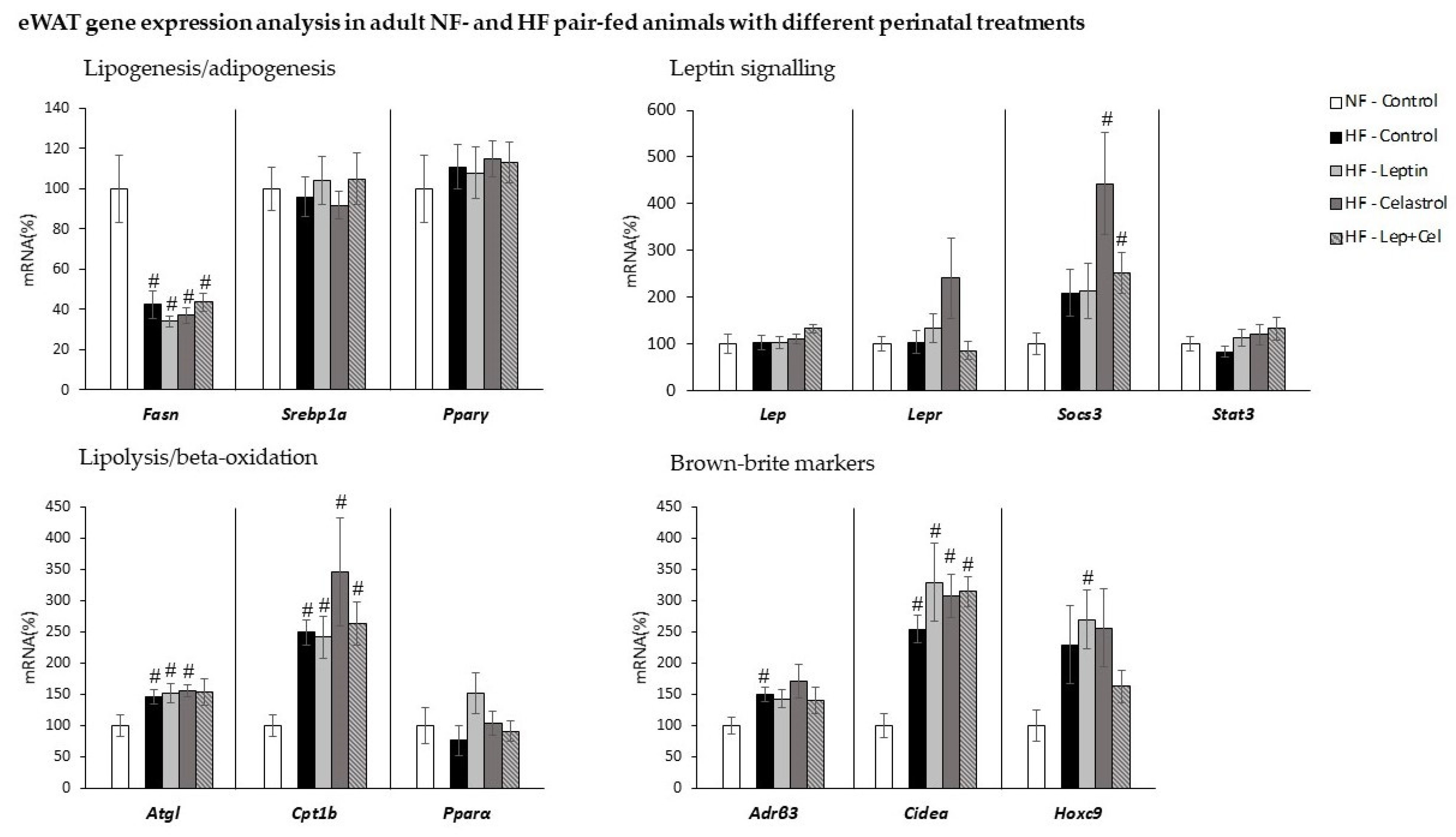
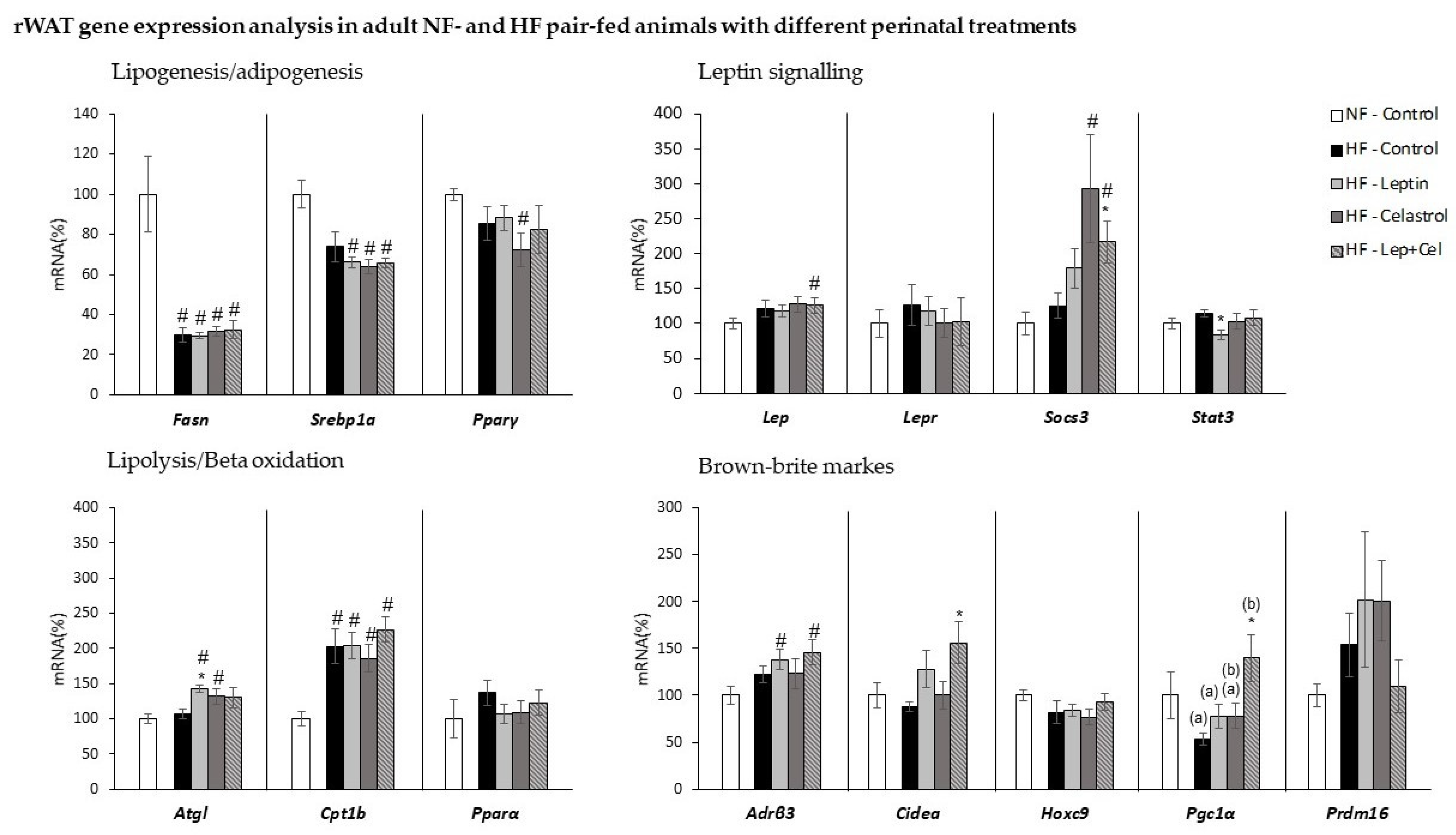
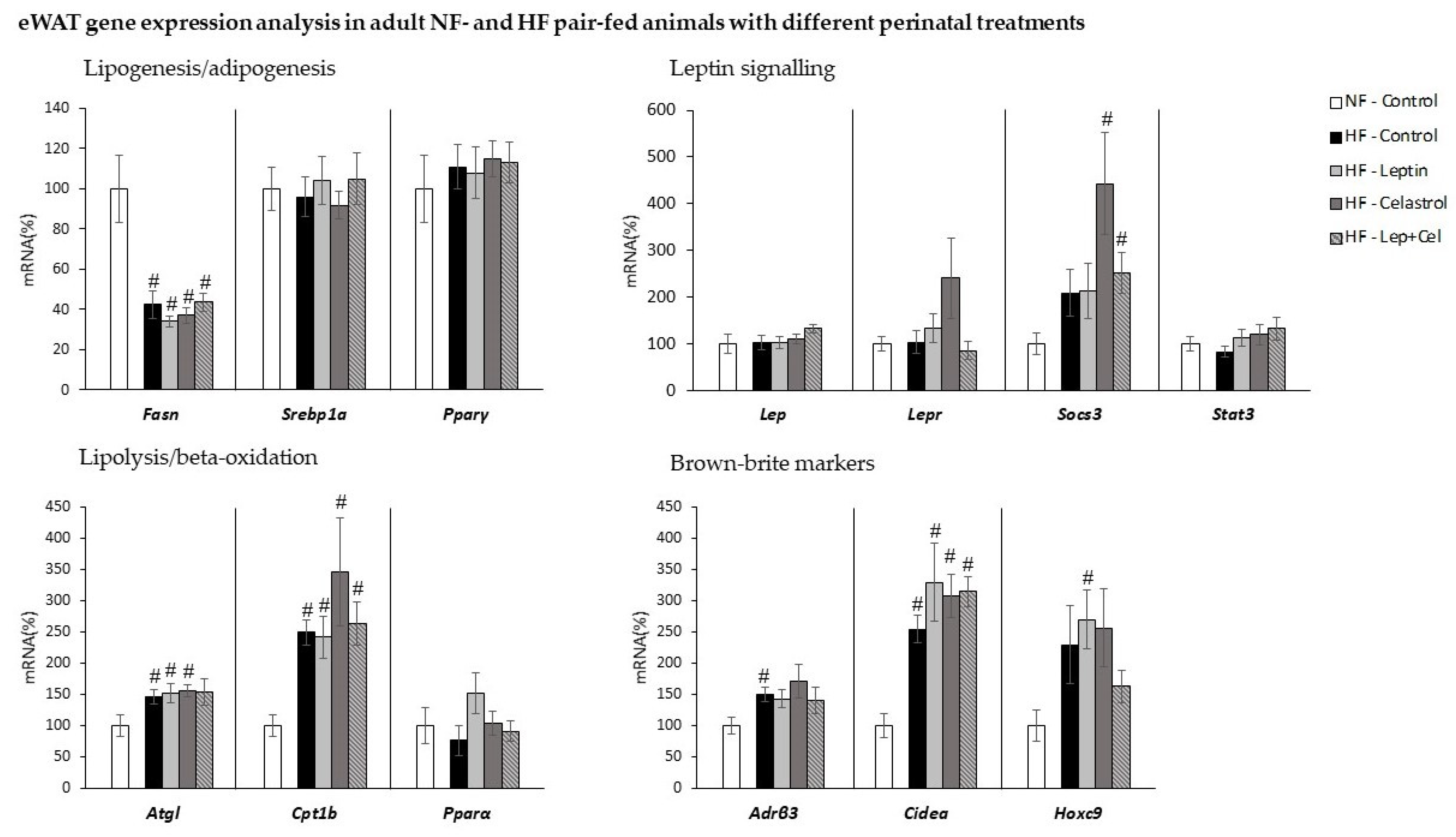
3.4. White Adipose Tissue Gene Expression and Histological and Immunohistochemical Analysis
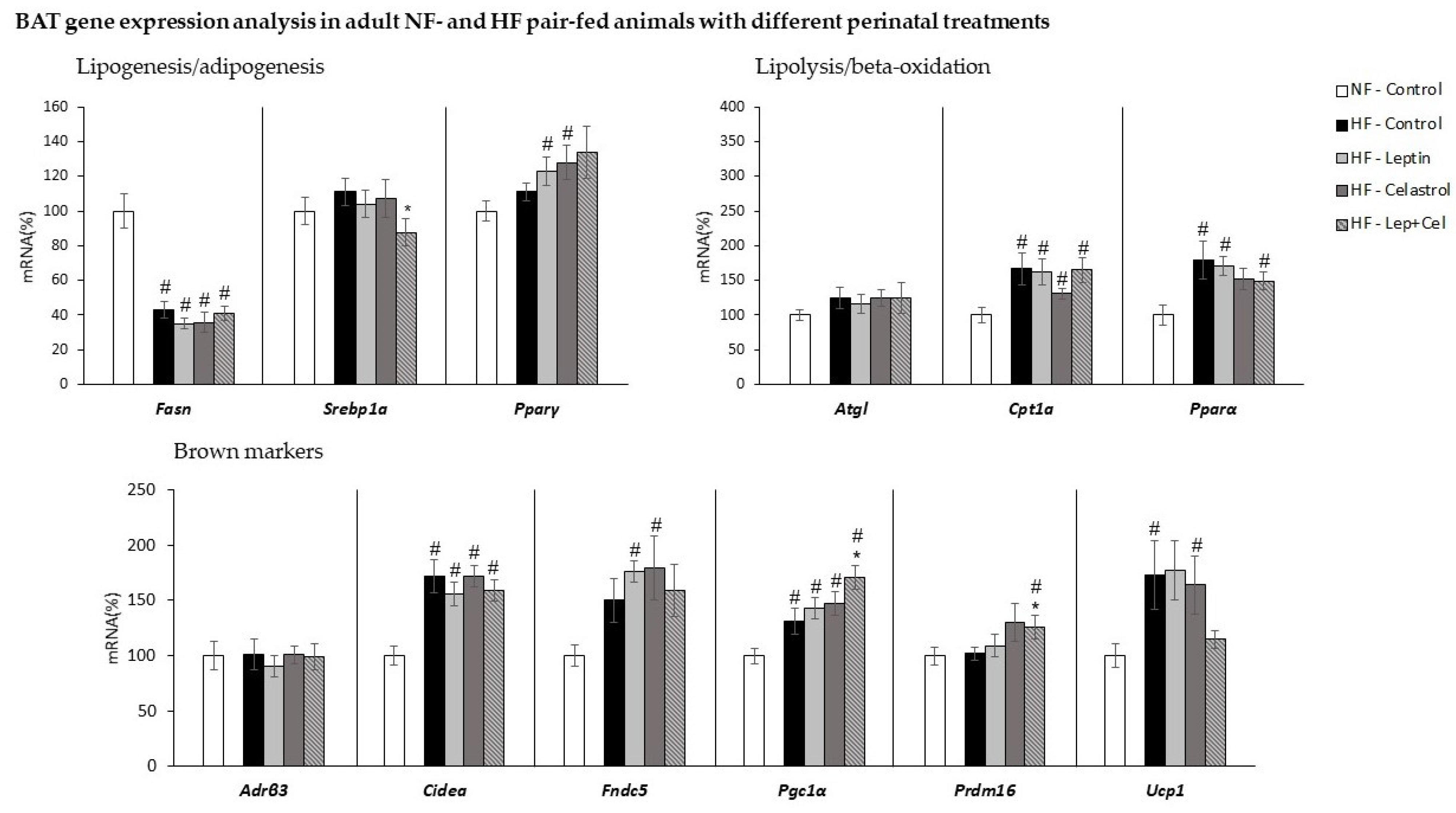
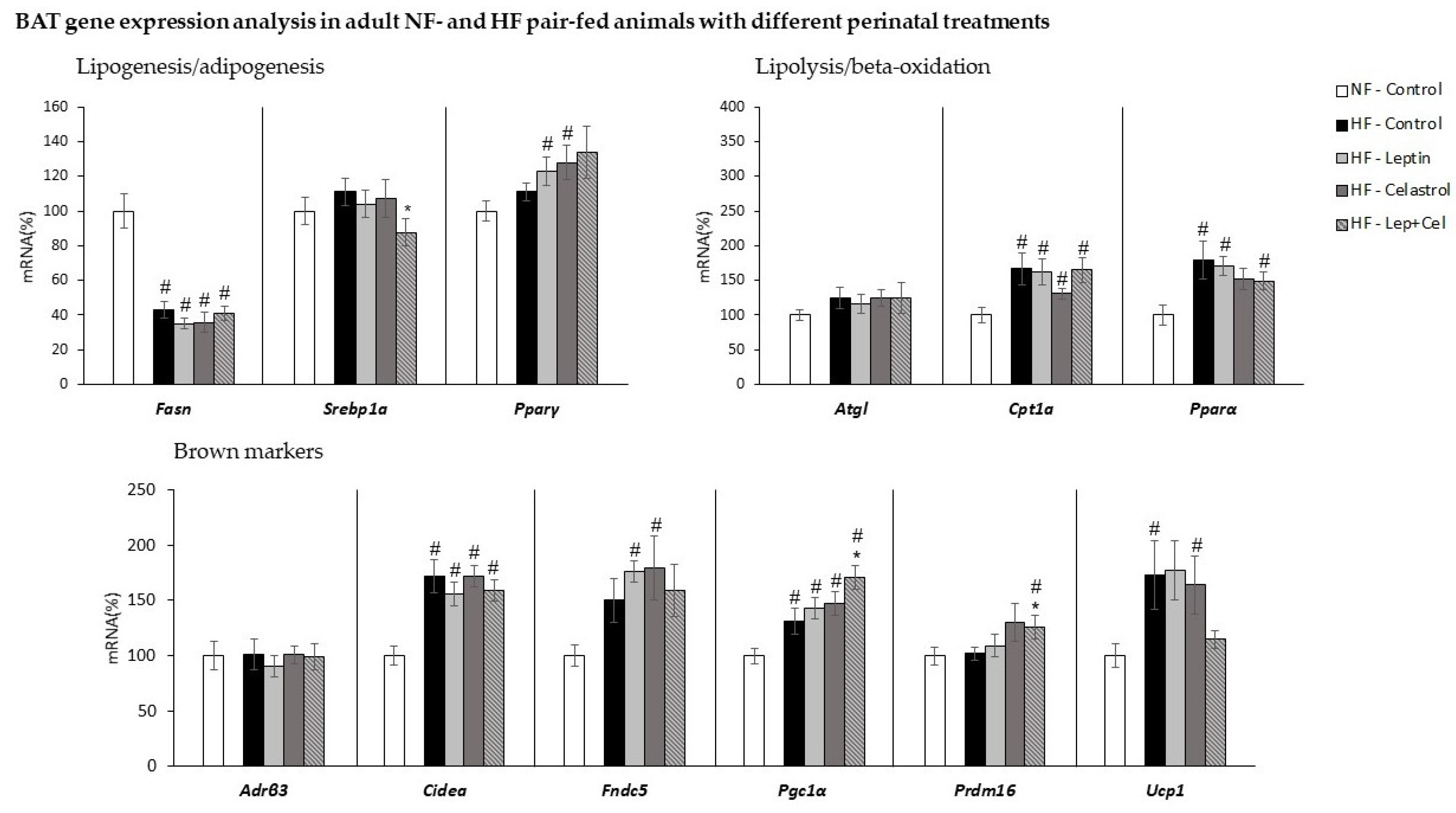
3.5. Brown Adipose Tissue Gene Expression Analysis
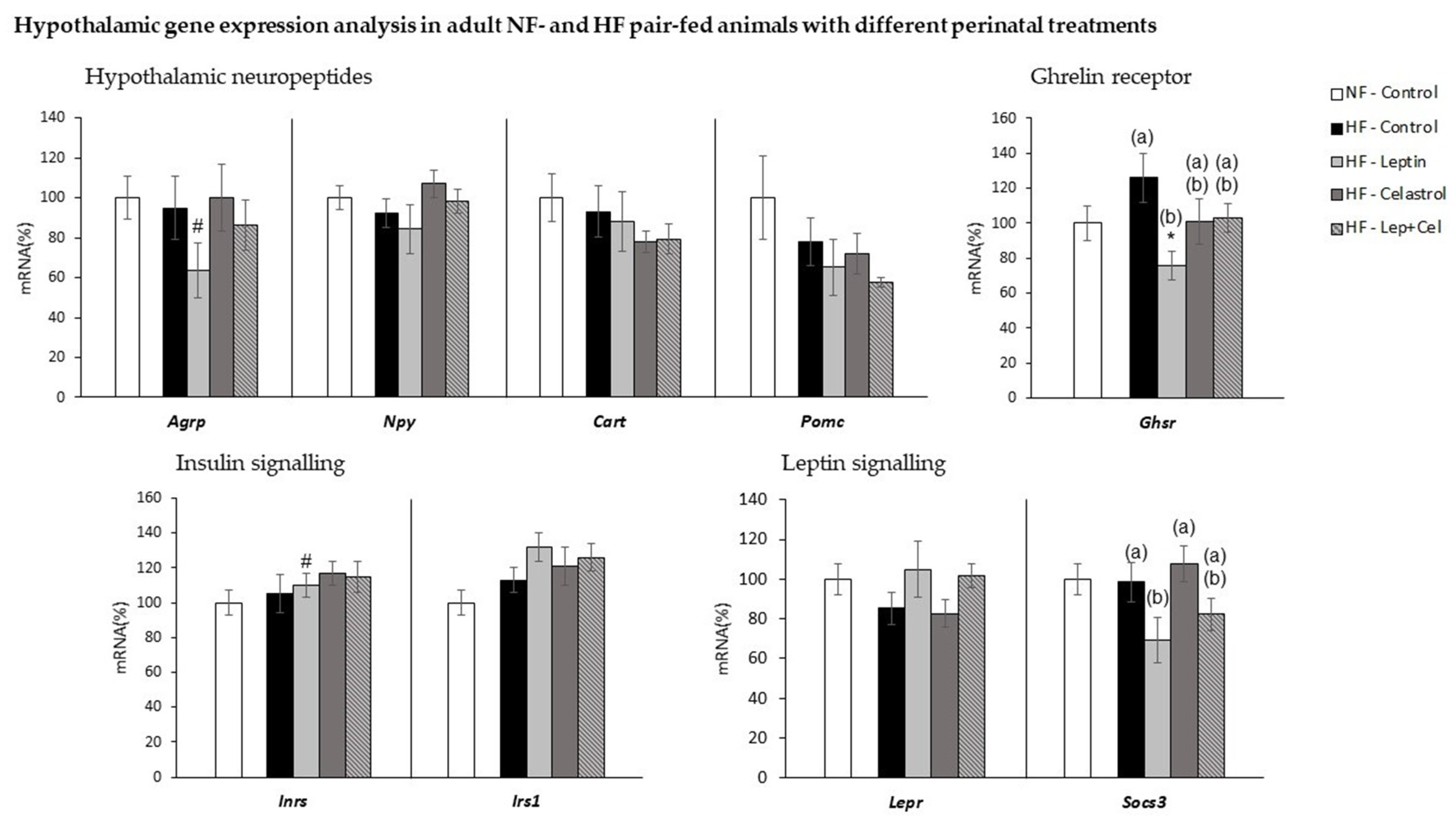
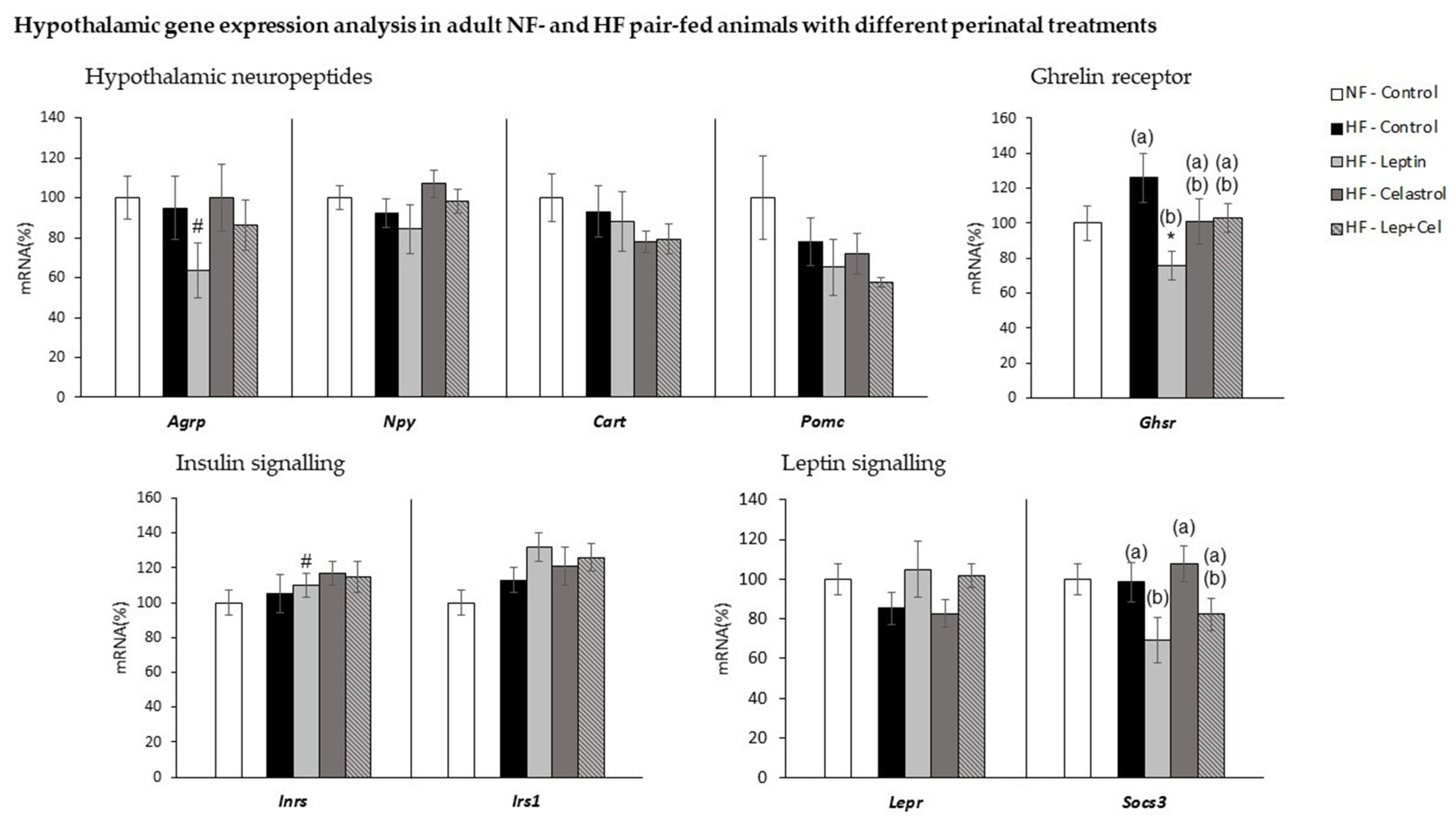
3.6. Hypothalamic Gene Expression Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Preston, J.D.; Reynolds, L.J.; Pearson, K.J. Developmental Origins of Health Span and Life Span: A Mini-Review. Gerontology 2018, 64, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Finn, S.; Culligan, E.; Snelling, W.; Sleator, R.D. Early Life Nutrition. Sci. Prog. 2018, 101, 332–359. [Google Scholar] [CrossRef]
- Harder, T.; Bergmann, R.; Kallischnigg, G.; Plagemann, A. Duration of Breastfeeding and Risk of Overweight: A Meta-Analysis. Am. J. Epidemiol. 2005, 162, 397–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillman, M.W.; Rifas-Shiman, S.L.; Camargo, C.A.; Berkey, C.S.; Frazier, A.L.; Rockett, H.R.; Field, A.E.; Colditz, G.A. Risk of overweight among adolescents who were breastfed as infants. JAMA 2001, 285, 2461–2467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picó, C.; Palou, M.; Pomar, C.A.; Palou, A. Benefits of Breastfeeding in INFANT health: A role for milk signaling peptides. In Molecular Nutrition: Mother and Infant; Vinciguerra, M., Sanchez, P.C., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 29–56. [Google Scholar] [CrossRef]
- Savino, F.; Liguori, S.A.; Fissore, M.F.; Oggero, R. Breast Milk Hormones and Their Protective Effect on Obesity. Int. J. Pediatr. Endocrinol. 2009, 2009, 327505. [Google Scholar] [CrossRef] [PubMed]
- Priego, T.; Sánchez, J.; Palou, A.; Picó, C. Leptin intake during the suckling period improves the metabolic response of adipose tissue to a high-fat diet. Int. J. Obes. 2010, 34, 809–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picó, C.; Oliver, P.; Sánchez, J.; Miralles, O.; Caimari, A.; Priego, T.; Palou, A. The intake of physiological doses of leptin during lactation in rats prevents obesity in later life. Int. J. Obes. 2007, 31, 1199–1209. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, J.; Priego, T.; Palou, M.; Tobaruela, A.; Palou, A.; Pico, C. Oral Supplementation with Physiological Doses of Leptin During Lactation in Rats Improves Insulin Sensitivity and Affects Food Preferences Later in Life. Endocrinology 2008, 149, 733–740. [Google Scholar] [CrossRef] [Green Version]
- LeDuc, C.A.; Skowronski, A.A.; Rosenbaum, M. The Role of Leptin in the Development of Energy Homeostatic Systems and the Maintenance of Body Weight. Front. Physiol. 2021, 12, 789519. [Google Scholar] [CrossRef]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Frederich, R.C.; Hamann, A.; Anderson, S.; Löllmann, B.; Lowell, B.B.; Flier, J.S. Leptin levels reflect body lipid content in mice: Evidence for diet-induced resistance to leptin action. Nat. Med. 1995, 1, 1311–1314. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lee, J.; Hernandez, M.A.S.; Mazitschek, R.; Ozcan, U. Treatment of Obesity with Celastrol. Cell 2015, 161, 999–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Geng, C.; Liu, X.; Li, M.; Gao, M.; Liu, X.; Fang, F.; Chang, Y. Celastrol ameliorates liver metabolic damage caused by a high-fat diet through Sirt1. Mol. Metab. 2017, 6, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shi, C.; Yang, X.; Yang, M.; Sun, H.; Wang, C. Celastrol suppresses obesity process via increasing antioxidant capacity and improving lipid metabolism. Eur. J. Pharmacol. 2014, 744, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Bel-Serrat, S.; Mouratidou, T.; Huybrechts, I.; Labayen, I.; Cuenca-García, M.; Palacios, G.; Breidenassel, C.; Molnár, D.; Roccaldo, R.; Widhalm, K.; et al. Associations between macronutrient intake and serum lipid profile depend on body fat in European adolescents: The Healthy Lifestyle in Europe by Nutrition in Adolescence (HELENA) study. Br. J. Nutr. 2014, 112, 2049–2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labayen, I.; Ruiz, J.R.; Ortega, F.B.; Huybrechts, I.; Rodríguez, G.; Jiménez-Pavón, D.; Roccaldo, R.; Nova, E.; Widhalm, K.; Kafatos, A.; et al. High fat diets are associated with higher abdominal adiposity regardless of physical activity in adolescents; the HELENA study. Clin. Nutr. 2014, 33, 859–866. [Google Scholar] [CrossRef]
- Choi, J.; Se-Young, O.; Lee, D.; Tak, S.; Hong, M.; Park, S.; Cho, B.; Park, M. Characteristics of diet patterns in metabolically obese, normal weight adults (Korean National Health and Nutrition Examination Survey III, 2005). Nutr. Metab. Cardiovasc. Dis. 2012, 22, 567–574. [Google Scholar] [CrossRef]
- Suliga, E.; Kozieł, D.; Cieśla, E.; Głuszek, S. Association between dietary patterns and metabolic syndrome in individuals with normal weight: A cross-sectional study. Nutr. J. 2015, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Ruderman, N.; Chisholm, D.; Pi-Sunyer, X.; Schneider, S. The metabolically obese, normal-weight individual revisited. Diabetes 1998, 47, 699–713. [Google Scholar] [CrossRef]
- Conus, F.; Allison, D.B.; Rabasa-Lhoret, R.; St-Onge, M.; St-Pierre, D.H.; Tremblay-Lebeau, A.; Poehlman, E.T. Metabolic and Behavioral Characteristics of Metabolically Obese but Normal-Weight Women. J. Clin. Endocrinol. Metab. 2004, 89, 5013–5020. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Miranda, J.; Perez-Martinez, P. It is time to define metabolically obese but normal-weight (MONW) individuals. Clin. Endocrinol. 2013, 79, 314–315. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhuang, R.; Luo, X.; Yin, L.; Pang, C.; Feng, T.; You, H.; Zhai, Y.; Ren, Y.; Zhang, L.; et al. Prevalence of Metabolically Healthy Obese and Metabolically Obese but Normal Weight in Adults Worldwide: A Meta-Analysis. Horm. Metab. Res. 2015, 47, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.; Oliver, P.; Miralles, O.; Ceresi, E.; Pico, C.; Palou, A. Leptin Orally Supplied to Neonate Rats Is Directly Uptaken by the Immature Stomach and May Regulate Short-Term Feeding. Endocrinology 2005, 146, 2575–2582. [Google Scholar] [CrossRef] [PubMed]
- Beig, A.; Agbaria, R.; Dahan, A. The use of captisol (SBE7-β-CD) in oral solubility-enabling formulations: Comparison to HPβCD and the solubility-permeability interplay. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2015, 77, 73–78. [Google Scholar] [CrossRef]
- Díaz-Rúa, R.; Keijer, J.; Caimari, A.; Van Schothorst, E.M.; Palou, A.; Oliver, P. Peripheral blood mononuclear cells as a source to detect markers of homeostatic alterations caused by the intake of diets with an unbalanced macronutrient composition. J. Nutr. Biochem. 2015, 26, 398–407. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Caimari, A.; Oliver, P.; Palou, A. Adipose triglyceride lipase expression and fasting regulation are differently affected by cold exposure in adipose tissues of lean and obese Zucker rats. J. Nutr. Biochem. 2012, 23, 1041–1050. [Google Scholar] [CrossRef]
- Reynés, B.; García-Ruiz, E.; Díaz-Rúa, R.; Palou, A.; Oliver, P. Reversion to a control balanced diet is able to restore body weight and to recover altered metabolic parameters in adult rats long-term fed on a cafeteria diet. Food Res. Int. 2014, 64, 839–848. [Google Scholar] [CrossRef]
- Reynés, B.; García-Ruiz, E.; Oliver, P.; Palou, A. Gene expression of peripheral blood mononuclear cells is affected by cold exposure. Am. J. Physiol. Integr. Comp. Physiol. 2015, 309, R824–R834. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the (-Delta Delta C(T)) 2 Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Caimari, A.; Oliver, P.; Keijer, J.; Palou, A. Peripheral Blood Mononuclear Cells as a Model to Study the Response of Energy Homeostasis-Related Genes to Acute Changes in Feeding Conditions. OMICS: A J. Integr. Biol. 2010, 14, 129–141. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, E.; Reynés, B.; Díaz-Rúa, R.; Ceresi, E.; Oliver, P.; Palou, A. The intake of high-fat diets induces the acquisition of brown adipocyte gene expression features in white adipose tissue. Int. J. Obes. 2015, 39, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Pluta, W.; Dudzińska, W.; Lubkowska, A. Metabolic Obesity in People with Normal Body Weight (MONW)-Review of Diagnostic Criteria. Int. J. Environ. Res. Public Health 2022, 19, 624. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Chan, Z.; Magkos, F. Lean, but not healthy: The 'metabolically obese, normal-weight' phenotype. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Rúa, R.; van Schothorst, E.M.; Keijer, J.; Palou, A.; Oliver, P. Isocaloric high-fat feeding directs hepatic metabolism to handling of nutrient imbalance promoting liver fat deposition. Int. J. Obes. 2016, 40, 1250–1259. [Google Scholar] [CrossRef]
- Díaz-Rúa, R.; García-Ruiz, E.; Caimari, A.; Palou, A.; Oliver, P. Sustained exposure to diets with an unbalanced macronutrient proportion alters key genes involved in energy homeostasis and obesity-related metabolic parameters in rats. Food Funct. 2014, 5, 3117–3131. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Loh, K.H.; Wu, M.; Morgan, D.A.; Schneeberger, M.; Yu, X.; Chi, J.; Kosse, C.; Kim, D.; Rahmouni, K.; et al. A leptin–BDNF pathway regulating sympathetic innervation of adipose tissue. Nature 2020, 583, 839–844. [Google Scholar] [CrossRef]
- Luo, D.; Guo, Y.; Cheng, Y.; Zhao, J.; Wang, Y.; Rong, J. Natural product celastrol suppressed macrophage M1 polarization against inflammation in diet-induced obese mice via regulating Nrf2/HO-1, MAP kinase and NF-κB pathways. Aging 2017, 9, 2069–2082. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Lee, M.H.; Nam, D.H.; Song, H.K.; Kang, Y.S.; Lee, J.E.; Kim, H.W.; Cha, J.J.; Hyun, Y.Y.; Han, S.Y.; et al. Celastrol, an NF-κB Inhibitor, Improves Insulin Resistance and Attenuates Renal Injury in db/db Mice. PLoS ONE 2013, 8, e62068. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Xu, L.; Alberobello, A.T.; Gavrilova, O.; Bagattin, A.; Skarulis, M.; Liu, J.; Finkel, T.; Mueller, E. Celastrol Protects against Obesity and Metabolic Dysfunction through Activation of a HSF1-PGC1α Transcriptional Axis. Cell Metab. 2015, 22, 695–708. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight and Adiposity Parameters | |||||
| NF | HF Pair-Fed | ||||
| Control | Control | Leptin | Celastrol | Lep+Cel | |
| Weight (g) | 409 ± 13 | 416 ± 6 | 410 ± 6 | 420 ± 6 | 427 ± 5 |
| Fat mass (%) | 19.4 ± 0.5 | 23.9 ± 1.8 # (a) | 22.9 ± 1.2 # (a) | 27.3 ± 0.8 # (b) | 25.6 ± 0.7 # (a,b) |
| Adiposity index (%) | 8.36 ± 0.26 | 11.2 ± 0.6 # (a,b) | 10.4 ± 0.4 # (a) | 11.9 ± 0.3 # (b) | 11.0 ± 0.3 # (a,b) |
| iWAT (g) | 9.73 ± 0.86 | 13.9 ± 1.2 # | 13.5 ± 0.9 # | 16.2 ± 0.7 # | 14.9 ± 0.8 # |
| eWAT (g) | 9.59 ± 0.66 | 13.7 ± 0.5 # (a) | 11.1 ± 0.5 (b) * | 13.1 ± 0.7 #(a) | 12.2 ± 0.6 # (a,b) |
| mWAT (g) | 4.04 ± 0.26 | 5.46 ± 0.4 # (a,b) | 4.83 ± 0.30 # (a) | 4.91 ± 0.36 #(a) | 6.04 ± 0.24 # (b) |
| rWAT (g) | 11.0 ± 0.9 | 13.1 ± 0.4 (a) | 13.2 ± 0.7 (a) | 15.8 ± 0.8 # (b) * | 13.7 ± 0.7 (a) |
| BAT (g) | 0.65 ± 0.05 | 0.71 ± 0.04 | 0.68 ± 0.04 | 0.80 ± 0.09 | 0.85 ± 0.06 # |
| Serum Parameters | |||||
| NF | HF Pair-Fed | ||||
| Control | Control | Leptin | Celastrol | Lep+Cel | |
| Glucose Fasting(mg/dL) | 101 ± 3 | 120 ± 7 # | 114 ± 4 | 113 ± 5 | 122 ± 4 # |
| Glucose Ad libitum (mg/dL) | 110 ± 2 | 111 ± 2 (a) | 116 ± 2 (a) | 120 ± 3 # (a,b) * | 127 ± 5 # (b) * |
| Insulin Fasting (µg/L) | 0.218 ± 0.046 | 0.330 ± 0.053 | 0.243 ± 0.027 | 0.408 ± 0.073 # | 0.341 ± 0.071 |
| HOMA-IR | 1.06 ± 0.16 | 2.45 ± 0.52 # | 1.69 ± 0.24 | 2.69 ± 0.48 # | 2.49 ± 0.54 # |
| Leptin (mg/mL) | 12.8 ± 1.0 | 18.1 ± 2.3 # (a) | 12.5 ± 1.3 (b) * | 18.7 ± 0.5 # (a) | 16.6 ± 1.3 # (a,b) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reynés, B.; Cifre, M.; Palou, A.; Oliver, P. Perinatal Treatment with Leptin, but Not Celastrol, Protects from Metabolically Obese, Normal-Weight Phenotype in Rats. Nutrients 2022, 14, 2277. https://doi.org/10.3390/nu14112277
Reynés B, Cifre M, Palou A, Oliver P. Perinatal Treatment with Leptin, but Not Celastrol, Protects from Metabolically Obese, Normal-Weight Phenotype in Rats. Nutrients. 2022; 14(11):2277. https://doi.org/10.3390/nu14112277
Chicago/Turabian StyleReynés, Bàrbara, Margalida Cifre, Andreu Palou, and Paula Oliver. 2022. "Perinatal Treatment with Leptin, but Not Celastrol, Protects from Metabolically Obese, Normal-Weight Phenotype in Rats" Nutrients 14, no. 11: 2277. https://doi.org/10.3390/nu14112277
APA StyleReynés, B., Cifre, M., Palou, A., & Oliver, P. (2022). Perinatal Treatment with Leptin, but Not Celastrol, Protects from Metabolically Obese, Normal-Weight Phenotype in Rats. Nutrients, 14(11), 2277. https://doi.org/10.3390/nu14112277