
Intestinal Mucosal Barrier Improvement with Prebiotics: Histological Evaluation of Longish Glucomannan Hydrolysates-Induced Innate T Lymphocyte Activities in Mice

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
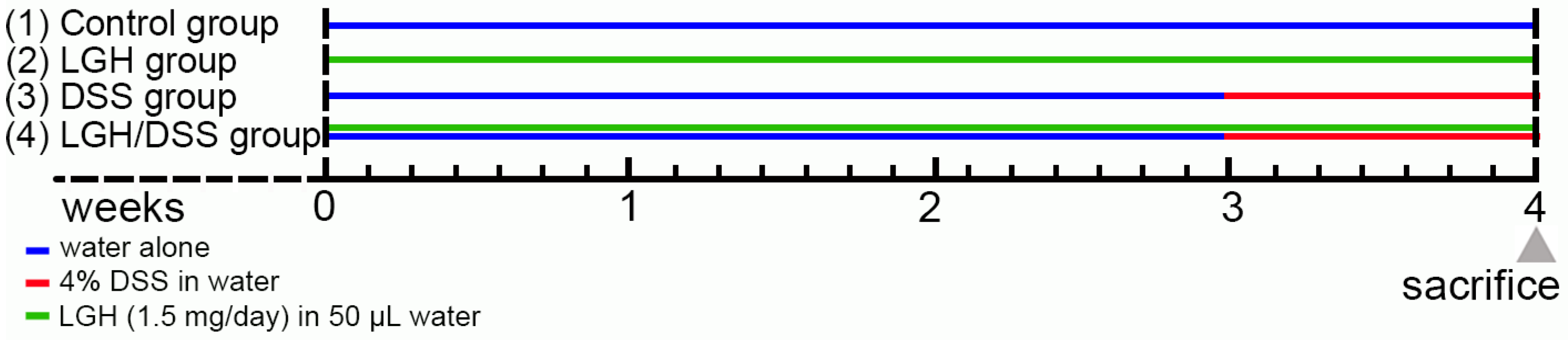
2.1. Mouse Model of DSS-Induced Colitis and LGH Supplementation
2.2. Inflammation Characteristics of Colon Tissue in Mice with Induced Colitis
2.3. Immunohistochemical Evaluations of Cytokine Expressions
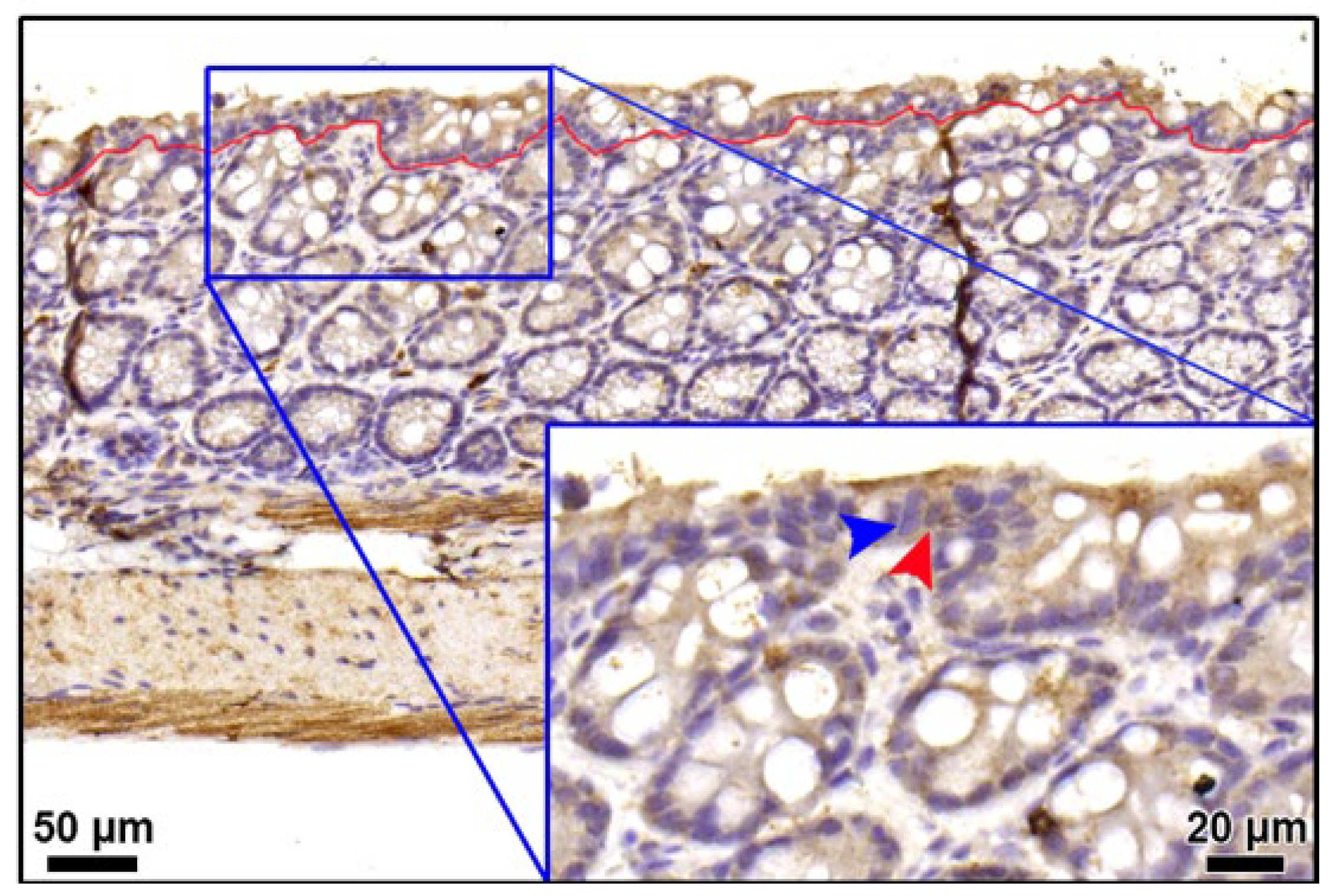
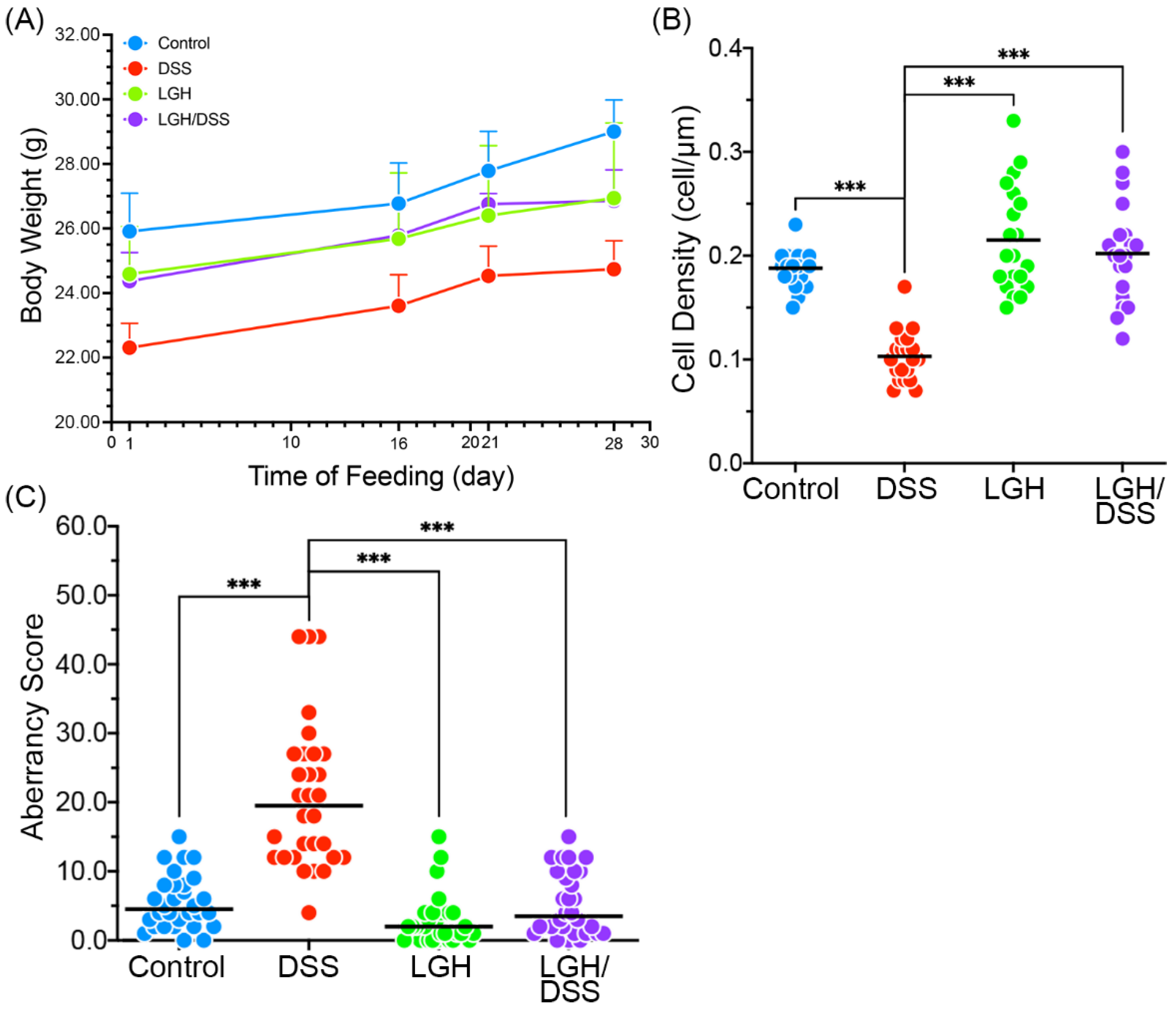
2.4. Calculation of Epithelial Cell Density
2.5. Assessment of Inflammatory Scores and Analysis of Aberrancy Score
2.6. Statistical Analysis
3. Results
3.1. Bodyweight Variance
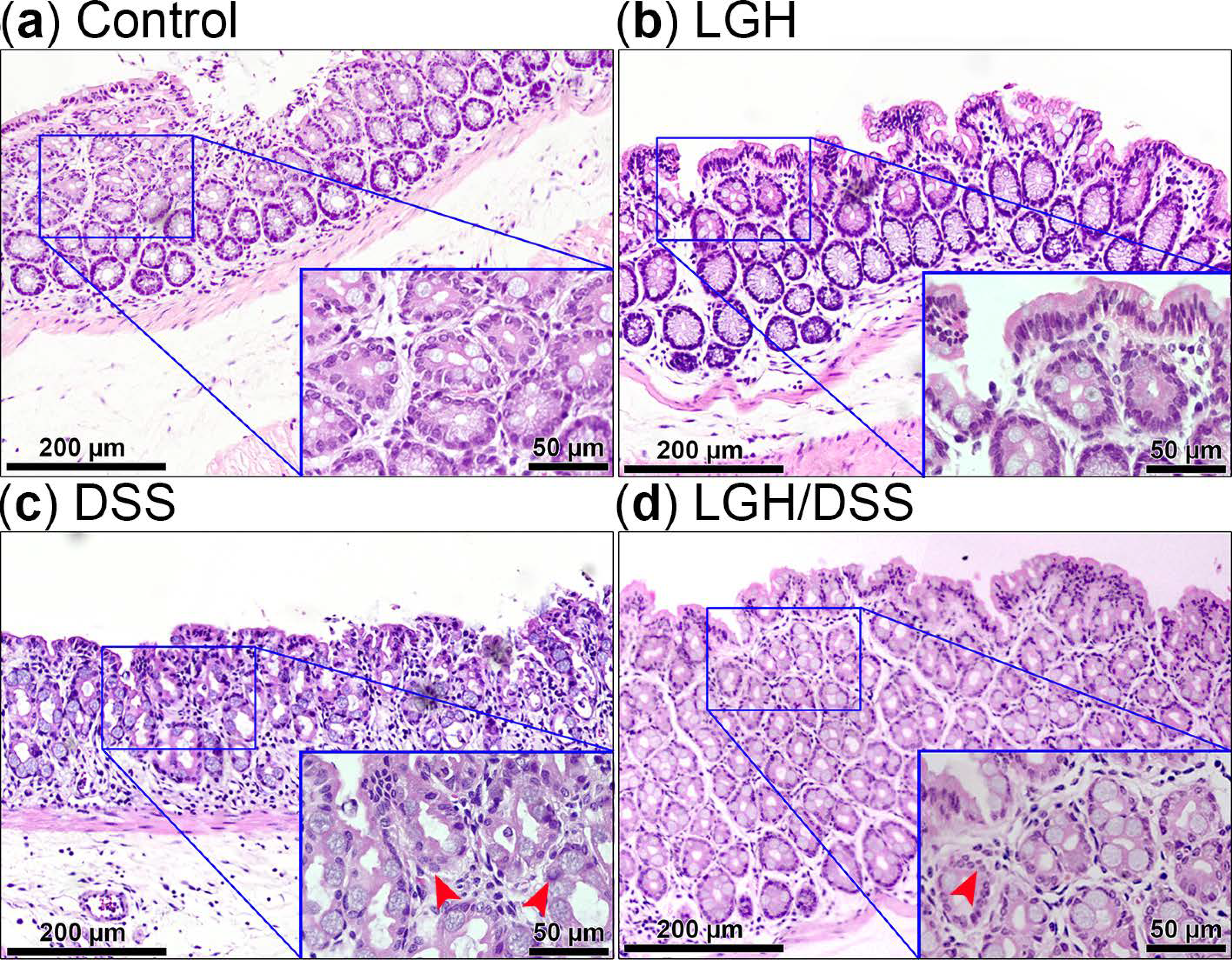
3.2. Integrity of Colonic Epithelium
3.3. LGH Manipulates DSS-Induced Inflammatory Responses
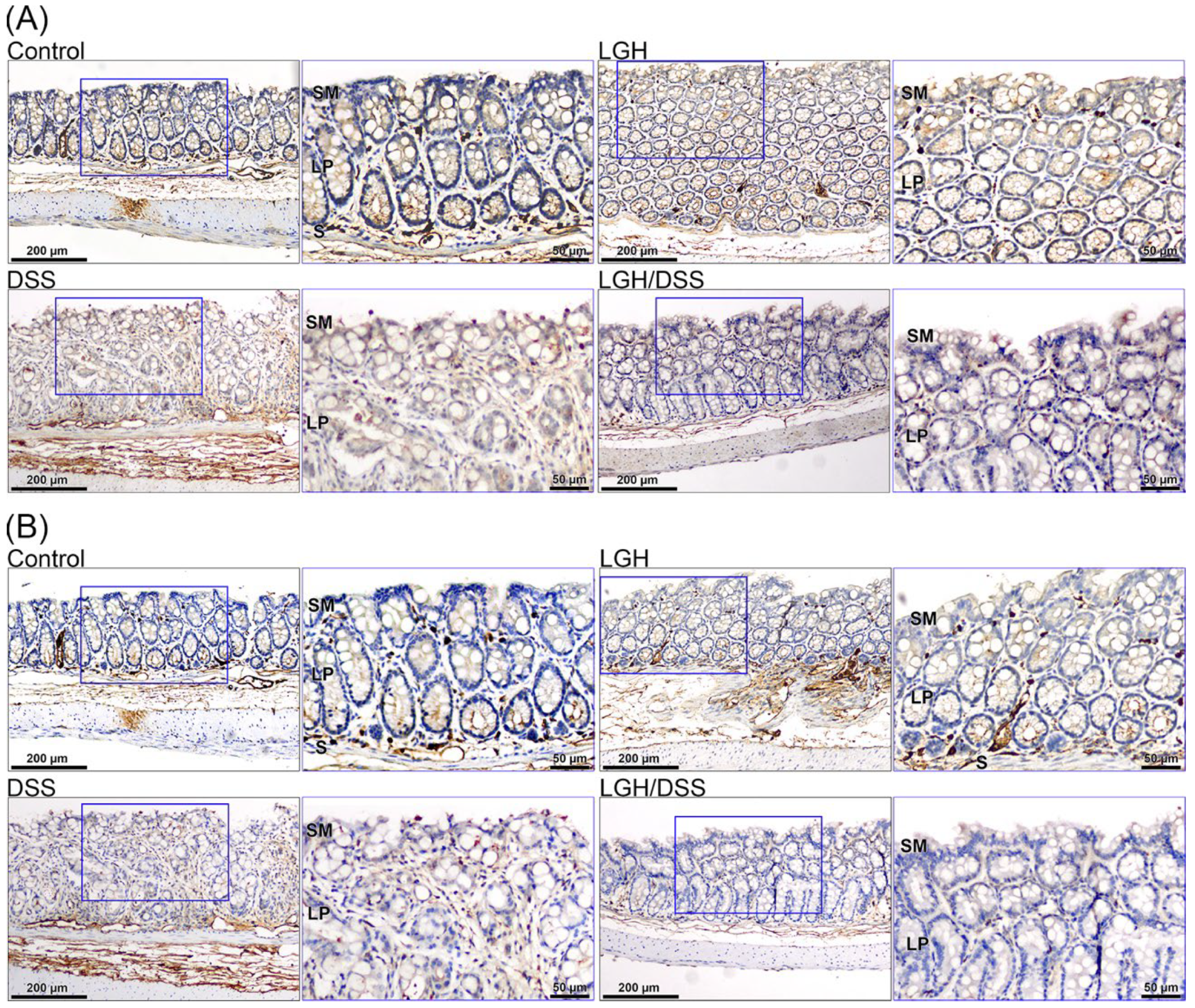
3.4. Lamina Propria T Lymphocyte Activation
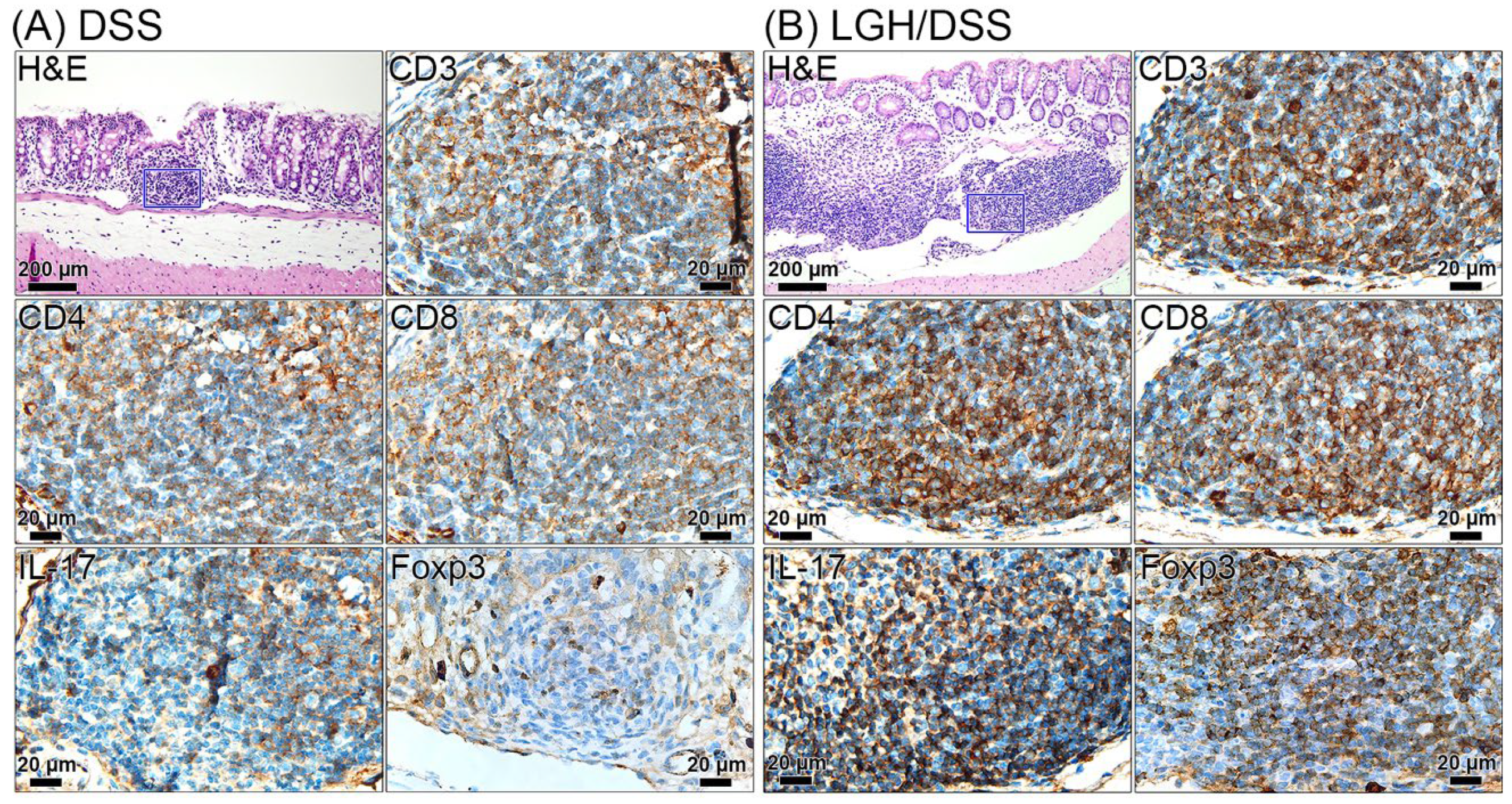
3.5. Lymphoid Aggregate Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut Biogeography of the Bacterial Microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Whitley, C.S.; Haribabu, B.; Jala, V.R. Regulation of Intestinal Barrier Function by Microbial Metabolites. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 1463–1482. [Google Scholar] [CrossRef] [PubMed]
- Fong, W.; Li, Q.; Yu, J. Gut Microbiota Modulation: A Novel Strategy for Prevention and Treatment of Colorectal Cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef]
- Brosseau, C.; Selle, A.; Palmer, D.J.; Prescott, S.L.; Barbarot, S.; Bodinier, M. Prebiotics: Mechanisms and Preventive Effects in Allergy. Nutrients 2019, 11, 1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D. Dietary Fiber and Prebiotics and the Gastrointestinal Microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Okumura, R.; Takeda, K. Maintenance of Intestinal Homeostasis by Mucosal Barriers. Inflamm. Regen. 2018, 38, 5. [Google Scholar] [CrossRef]
- Mörbe, U.M.; Jørgensen, P.B.; Fenton, T.M.; von Burg, N.; Riis, L.B.; Spencer, J.; Agace, W.W. Human Gut-Associated Lymphoid Tissues (GALT); Diversity, Structure, and Function. Mucosal Immunol. 2021, 14, 793–802. [Google Scholar] [CrossRef]
- Fenton, T.M.; Jørgensen, P.B.; Niss, K.; Rubin, S.J.S.; Mörbe, U.M.; Riis, L.B.; Da Silva, C.; Plumb, A.; Vandamme, J.; Jakobsen, H.L.; et al. Immune Profiling of Human Gut-Associated Lymphoid Tissue Identifies a Role for Isolated Lymphoid Follicles in Priming of Region-Specific Immunity. Immunity 2020, 52, 557–570.e6. [Google Scholar] [CrossRef]
- Yan, J.; Luo, M.; Chen, Z.; He, B. The Function and Role of the Th17/Treg Cell Balance in Inflammatory Bowel Disease. J. Immunol. Res. 2020, 2020, 8813558. [Google Scholar] [CrossRef]
- Tokuhara, D.; Kurashima, Y.; Kamioka, M.; Nakayama, T.; Ernst, P.; Kiyono, H. A Comprehensive Understanding of the Gut Mucosal Immune System in Allergic Inflammation. Allergol. Int. 2019, 68, 17–25. [Google Scholar] [CrossRef]
- Simon, E.; Călinoiu, L.F.; Mitrea, L.; Vodnar, D.C. Probiotics, Prebiotics, and Synbiotics: Implications and Beneficial Effects against Irritable Bowel Syndrome. Nutrients 2021, 13, 2112. [Google Scholar] [CrossRef]
- Vodnar, D.-C.; Mitrea, L.; Teleky, B.-E.; Szabo, K.; Călinoiu, L.-F.; Nemeş, S.-A.; Martău, G.-A. Coronavirus Disease (COVID-19) Caused by (SARS-CoV-2) Infections: A Real Challenge for Human Gut Microbiota. Front. Cell. Infect. Microbiol. 2020, 10, 575559. [Google Scholar] [CrossRef]
- Changchien, C.H.; Han, Y.C.; Chen, H.L. Konjac Glucomannan Polysaccharide and Inulin Oligosaccharide Enhance the Colonic Mucosal Barrier Function and Modulate Gut-Associated Lymphoid Tissue Immunity in C57BL/6J Mice. Br. J. Nutr. 2020, 123, 319–327. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, J.; Yang, F.-Q. Konjac Glucomannan, a Promising Polysaccharide for OCDDS. Carbohydr. Polym. 2014, 104, 175–181. [Google Scholar] [CrossRef]
- Yin, J.Y.; Ma, L.Y.; Xie, M.Y.; Nie, S.P.; Wu, J.Y. Molecular Properties and Gut Health Benefits of Enzyme-Hydrolyzed Konjac Glucomannans. Carbohydr. Polym. 2020, 237, 116117. [Google Scholar] [CrossRef]
- Zhu, F. Modifications of Konjac Glucomannan for Diverse Applications. Food Chem. 2018, 256, 419–426. [Google Scholar] [CrossRef]
- Cochran, K.E.; Lamson, N.G.; Whitehead, K.A. Expanding the Utility of the Dextran Sulfate Sodium (DSS) Mouse Model to Induce a Clinically Relevant Loss of Intestinal Barrier Function. PeerJ 2020, 8, e8681. [Google Scholar] [CrossRef]
- Jackson, P.; Blythe, D. Immunohistochemical Techniques. In Bancroft’s Theory and Practice of Histological Techniques; Elsevier: Amsterdam, The Netherlands, 2013; pp. 381–426. [Google Scholar]
- Bancroft, J.D.; Layton, C. The Hematoxylins and Eosin. In Bancroft’s Theory and Practice of Histological Techniques; Elsevier: Amsterdam, The Netherlands, 2013; pp. 173–186. [Google Scholar]
- Singh, J.; Thachil, T.; Eapen, M.S.; Lim, A.; Sufyan, W.; Rawson, R.; Duncan, H.; Ieso, P.D.E.; Sohal, S.S. Immunohistochemical Investigation of Cytokine Expression Levels as Biomarkers in Transrectal Ultrasound - Guided Needle Biopsy Specimens of Prostate Adenocarcinoma. Mol. Clin. Oncol. 2021, 15, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Dieleman, L.A.; Palmen, M.J.H.J.; Akol, H.; Bloemena, E.; Peña, A.S.; Meuwissen, S.G.M.; Van Rees, E.P. Chronic Experimental Colitis Induced by Dextran Sulphate Sodium (DSS) Is Characterized by Th1 and Th2 Cytokines. Clin. Exp. Immunol. 1998, 114, 385–391. [Google Scholar] [CrossRef]
- Kihara, N. Vanilloid Receptor-1 Containing Primary Sensory Neurones Mediate Dextran Sulphate Sodium Induced Colitis in Rats. Gut 2003, 52, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.; Jun, Y. Measurement of Pain-like Response to Various NICU Stimulants for High-Risk Infants. Early Hum. Dev. 2007, 83, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Klose, C.S.N.; Artis, D. Innate Lymphoid Cells Control Signaling Circuits to Regulate Tissue-Specific Immunity. Cell Res. 2020, 30, 475–491. [Google Scholar] [CrossRef]
- Hao, Z.; Wang, X.; Yang, H.; Tu, T.; Zhang, J.; Luo, H.; Huang, H.; Su, X. PUL-Mediated Plant Cell Wall Polysaccharide Utilization in the Gut Bacteroidetes. Int. J. Mol. Sci. 2021, 22, 3077. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Yokoi, T.; Sugiyama, M.; Osawa, R.; Mukai, T.; Okada, N. Roles of the Cell Surface Architecture of Bacteroides and Bifidobacterium in the Gut Colonization. Front. Microbiol. 2021, 12, 754819. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Burgueño, J.F.; Abreu, M.T. Epithelial Toll-like Receptors and Their Role in Gut Homeostasis and Disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 263–278. [Google Scholar] [CrossRef]
- La Rosa, S.L.; Leth, M.L.; Michalak, L.; Hansen, M.E.; Pudlo, N.A.; Glowacki, R.; Pereira, G.; Workman, C.T.; Arntzen, M.Ø.; Pope, P.B.; et al. The Human Gut Firmicute Roseburia Intestinalis Is a Primary Degrader of Dietary β-Mannans. Nat. Commun. 2019, 10, 905. [Google Scholar] [CrossRef]
- Lindstad, L.J.; Lo, G.; Leivers, S.; Lu, Z.; Michalak, L.; Pereira, G.V.; Røhr, Å.K.; Martens, E.C.; McKee, L.S.; Louis, P.; et al. Human Gut Faecalibacterium Prausnitzii Deploys a Highly Efficient Conserved System To Cross-Feed on β-Mannan-Derived Oligosaccharides. MBio 2021, 12, e03628–e03720. [Google Scholar] [CrossRef]
- Michalak, L.; Gaby, J.C.; Lagos, L.; La Rosa, S.L.; Hvidsten, T.R.; Tétard-Jones, C.; Willats, W.G.T.; Terrapon, N.; Lombard, V.; Henrissat, B.; et al. Microbiota-Directed Fibre Activates Both Targeted and Secondary Metabolic Shifts in the Distal Gut. Nat. Commun. 2020, 11, 5773. [Google Scholar] [CrossRef]
- Spencer, C.N.; McQuade, J.L.; Gopalakrishnan, V.; McCulloch, J.A.; Vetizou, M.; Cogdill, A.P.; Khan, M.A.W.; Zhang, X.; White, M.G.; Peterson, C.B.; et al. Dietary Fiber and Probiotics Influence the Gut Microbiome and Melanoma Immunotherapy Response. Science 2021, 374, 1632–1640. [Google Scholar] [CrossRef]
- Cabrita, R.; Lauss, M.; Sanna, A.; Donia, M.; Skaarup Larsen, M.; Mitra, S.; Johansson, I.; Phung, B.; Harbst, K.; Vallon-Christersson, J.; et al. Tertiary Lymphoid Structures Improve Immunotherapy and Survival in Melanoma. Nature 2020, 577, 561–565. [Google Scholar] [CrossRef]
- Goc, J.; Lv, M.; Bessman, N.J.; Flamar, A.L.; Sahota, S.; Suzuki, H.; Teng, F.; Putzel, G.G.; Eberl, G.; Withers, D.R.; et al. Dysregulation of ILC3s Unleashes Progression and Immunotherapy Resistance in Colon Cancer. Cell 2021, 184, 5015–5030.e16. [Google Scholar] [CrossRef]
- Lochner, M.; Ohnmacht, C.; Presley, L.; Bruhns, P.; Si-Tahar, M.; Sawa, S.; Eberl, G. Microbiota-Induced Tertiary Lymphoid Tissues Aggravate Inflammatory Disease in the Absence of RORγt and LTi Cells. J. Exp. Med. 2011, 208, 125–134. [Google Scholar] [CrossRef]
- Kuhn, K.A.; Schulz, H.M.; Regner, E.H.; Severs, E.L.; Hendrickson, J.D.; Mehta, G.; Whitney, A.K.; Ir, D.; Ohri, N.; Robertson, C.E.; et al. Bacteroidales Recruit IL-6-Producing Intraepithelial Lymphocytes in the Colon to Promote Barrier Integrity. Mucosal Immunol. 2018, 11, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Sano, T.; Kageyama, T.; Fang, V.; Kedmi, R.; Martinez, C.S.; Talbot, J.; Chen, A.; Cabrera, I.; Gorshko, O.; Kurakake, R.; et al. Redundant Cytokine Requirement for Intestinal Microbiota-Induced Th17 Cell Differentiation in Draining Lymph Nodes. Cell Rep. 2021, 36, 109608. [Google Scholar] [CrossRef]
- Jump, R.L.; Levine, A.D. Mechanisms of Natural Tolerance in the Intestine. Inflamm. Bowel Dis. 2004, 10, 462–478. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | β | SE | χ2 | p |
|---|---|---|---|---|
| Time | ||||
| Pre-test | Ref | |||
| Post-test | 0.66 | 0.07 | 90.3 | *** |
| Group | ||||
| Control | Ref | |||
| DSS | −3.26 | 0.63 | 26.7 | *** |
| LGH | −1.38 | 1.12 | 1.53 | 0.22 |
| LGH/DSS | −1.03 | 0.48 | 4.60 | * |
| Group × Time | ||||
| Control × Pre-test | Ref | |||
| Control × Post-test | Ref | |||
| DSS × Pre-test | Ref | |||
| DSS × Post-test | −0.45 | 0.08 | 33.7 | *** |
| LGH × Pre-test | Ref | |||
| LGH × Post-test | −0.12 | 0.12 | 1.01 | 0.32 |
| LGH/DSS × Pre-test | Ref | |||
| LGH/DSS × Post-test | −0.55 | 0.33 | 2.72 | 0.10 |
| Intercept | 27.8 | 0.46 | 3724 | *** |
| Feature | Group (G) | Inflammatory Score | Multiple Comparisons | |||
|---|---|---|---|---|---|---|
| Mean | SE | (G)–Control | (G)–LGH | (G)–DSS | ||
| Inflammatory | Control | 0.6 | 0.2 | 0.0 | 0.2 | −3.5 *** |
| severity | DSS | 4.1 | 0.4 | 3.5 *** | 3.7 *** | 0.0 |
| LGH | 0.4 | 0.2 | −0.2 | 0.0 | −3.7 *** | |
| LGH/DSS | 0.8 | 0.2 | 0.2 | 0.4 | −3.3 *** | |
| Inflammatory | Control | 0.9 | 0.3 | 0.0 | 0.5 | −2.4 *** |
| extent | DSS | 3.3 | 0.3 | 2.4 *** | 2.9 *** | 0.0 |
| LGH | 0.4 | 0.2 | −0.5 | 0.0 | −2.9 *** | |
| LGH/DSS | 1.2 | 0.3 | 0.3 | 0.8 | −2.1 *** | |
| Epithelium | Control | 0.3 | 0.1 | 0.0 | 0.0 | −2.9 *** |
| regeneration | DSS | 3.2 | 0.5 | 2.9 *** | 2.9 *** | 0.0 |
| LGH | 0.3 | 0.1 | 0.0 | 0.0 | −2.9 *** | |
| LGH/DSS | 0.2 | 0.1 | −0.1 | −0.1 | −3.0 *** | |
| Crypt | Control | 0.4 | 0.1 | 0.0 | 0.2 | −2.6 *** |
| damage | DSS | 3.0 | 0.5 | 2.6 *** | 2.8 *** | 0.0 |
| LGH | 0.2 | 0.1 | −0.2 | 0.0 | −2.8 *** | |
| LGH/DSS | 0.2 | 0.1 | −0.2 | 0.0 | −2.8 *** | |
| Location | Group (G) | Grade | Multiple Comparisons | |||
|---|---|---|---|---|---|---|
| Mean | SE | G–Control | G–LGH | G–DSS | ||
| Superficial | Control | 0.8 | 0.1 | 0.0 | 0.2 | −1.7 *** |
| mucosa (SM) | DSS | 2.5 | 0.2 | 1.7 *** | 1.9 *** | 0.0 |
| LGH | 0.6 | 0.1 | −0.2 | 0.0 | −1.9 *** | |
| LGH/DSS | 0.3 | 0.1 | −0.5 * | −0.3 | −2.2 *** | |
| Lamina | Control | 1.7 | 0.1 | 0.0 | 0.3 | −0.5 |
| propria (LP) | DSS | 2.2 | 0.2 | 0.5 | 0.8 *** | 0.0 |
| LGH | 1.4 | 0.1 | −0.3 | 0.0 | −0.8 *** | |
| LGH/DSS | 1.4 | 0.1 | −0.3 | 0.0 | −0.8 *** | |
| Submucosa (S) | Control | 2.0 | 0.2 | 0.0 | 0.5 | −1.3 *** |
| DSS | 3.3 | 0.1 | 1.3 *** | 0.8 *** | 0.0 | |
| LGH | 1.5 | 0.2 | −0.5 | 0.0 | −0.8 *** | |
| LGH/DSS | 2.3 | 0.1 | 0.3 | 0.8 *** | −1.0 *** | |
| Location | Group (G) | Grade | Multiple Comparisons | |||
|---|---|---|---|---|---|---|
| Mean | SE | G–Control | G–LGH | G–DSS | ||
| Superficial | Control | 1.2 | 0.1 | 0.0 | −0.4 | −2.1 *** |
| mucosa (SM) | DSS | 3.3 | 0.2 | 2.1 *** | 1.7 *** | 0.0 |
| LGH | 1.6 | 0.1 | 0.4 | 0.0 | −1.7 *** | |
| LGH/DSS | 1.4 | 0.2 | 0.2 | −0.2 | −1.9 *** | |
| Lamina | Control | 1.1 | 0.1 | 0.0 | −0.2 | −2.0 *** |
| propria (LP) | DSS | 3.1 | 0.1 | 2.0 *** | 1.7*** | 0.0 |
| LGH | 1.3 | 0.1 | 0.2 | 0.0 | −1.7 *** | |
| LGH/DSS | 1.3 | 0.1 | 0.2 | 0.0 | −1.7 *** | |
| Submucosa (S) | Control | 0.2 | 0.1 | 0.0 | 0.1 | −2.4 *** |
| DSS | 2.6 | 0.2 | 2.4 *** | 2.5 *** | 0.0 | |
| LGH | 0.1 | 0.1 | −0.1 | 0.0 | −2.5 *** | |
| LGH/DSS | 0.7 | 0.1 | 0.5 | 0.6 | −1.9 *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, S.-C.; Chiang, H.-H.; Liu, C.-Y.; Li, Y.-J.; Lu, C.-L.; Lee, Y.-P.; Huang, C.-J.; Lai, C.-L. Intestinal Mucosal Barrier Improvement with Prebiotics: Histological Evaluation of Longish Glucomannan Hydrolysates-Induced Innate T Lymphocyte Activities in Mice. Nutrients 2022, 14, 2220. https://doi.org/10.3390/nu14112220
Chang S-C, Chiang H-H, Liu C-Y, Li Y-J, Lu C-L, Lee Y-P, Huang C-J, Lai C-L. Intestinal Mucosal Barrier Improvement with Prebiotics: Histological Evaluation of Longish Glucomannan Hydrolysates-Induced Innate T Lymphocyte Activities in Mice. Nutrients. 2022; 14(11):2220. https://doi.org/10.3390/nu14112220
Chicago/Turabian StyleChang, Shih-Chang, Hui-Hsun Chiang, Chih-Yi Liu, Yu-Ju Li, Chung-Lun Lu, Yung-Pin Lee, Chi-Jung Huang, and Ching-Long Lai. 2022. "Intestinal Mucosal Barrier Improvement with Prebiotics: Histological Evaluation of Longish Glucomannan Hydrolysates-Induced Innate T Lymphocyte Activities in Mice" Nutrients 14, no. 11: 2220. https://doi.org/10.3390/nu14112220
APA StyleChang, S.-C., Chiang, H.-H., Liu, C.-Y., Li, Y.-J., Lu, C.-L., Lee, Y.-P., Huang, C.-J., & Lai, C.-L. (2022). Intestinal Mucosal Barrier Improvement with Prebiotics: Histological Evaluation of Longish Glucomannan Hydrolysates-Induced Innate T Lymphocyte Activities in Mice. Nutrients, 14(11), 2220. https://doi.org/10.3390/nu14112220