
Docosahexaenoic Acid Suppresses Expression of Adipogenic Tetranectin through Sterol Regulatory Element-Binding Protein and Forkhead Box O Protein in Pigs

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Study
2.2. Cell Culture and Adipocyte Differentiation from Porcine Adipose Stem Cells
2.3. Cloning Porcine TN cDNA
2.4. Cloning the Putative Porcine TN Promoter
2.5. Luciferase Reporter Assay
2.6. Analysis of Quantitative Real-Time PCR (qPCR)
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. Cloning of Porcine TN
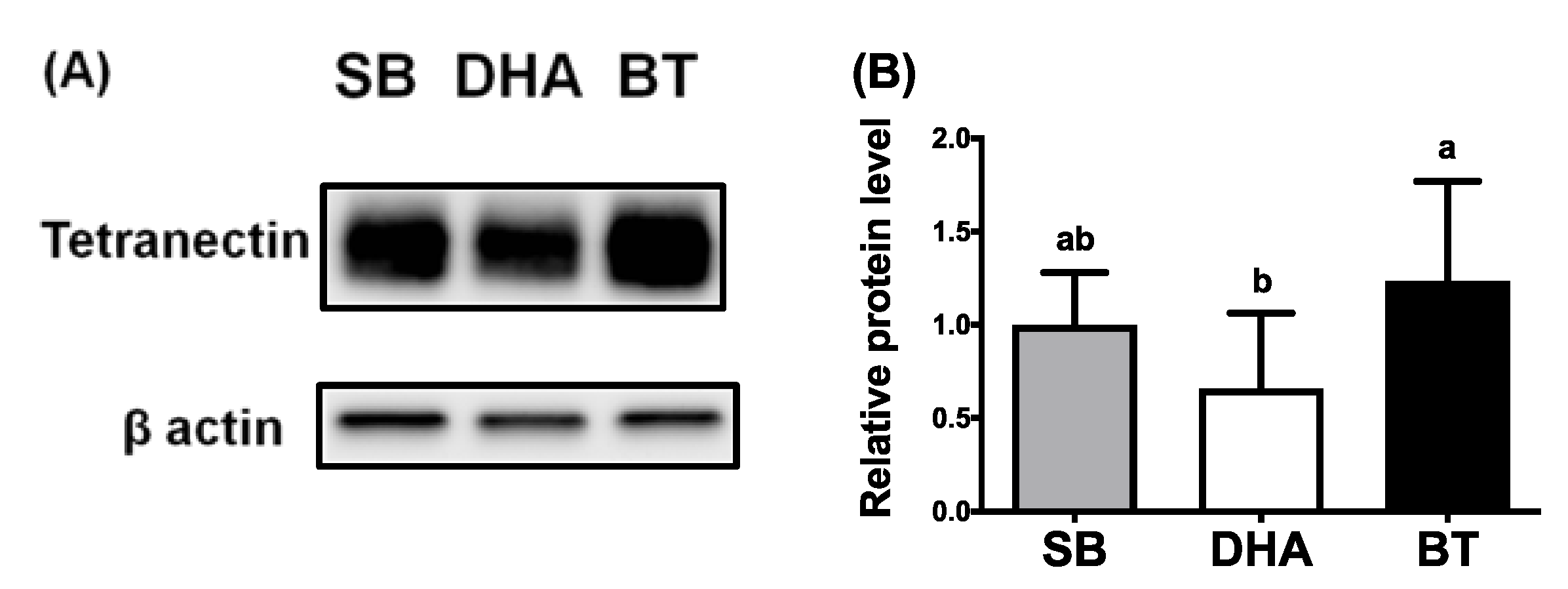
3.2. DHA Reduced Expression of TN in Porcine Adipose, Lung and Liver Tissues
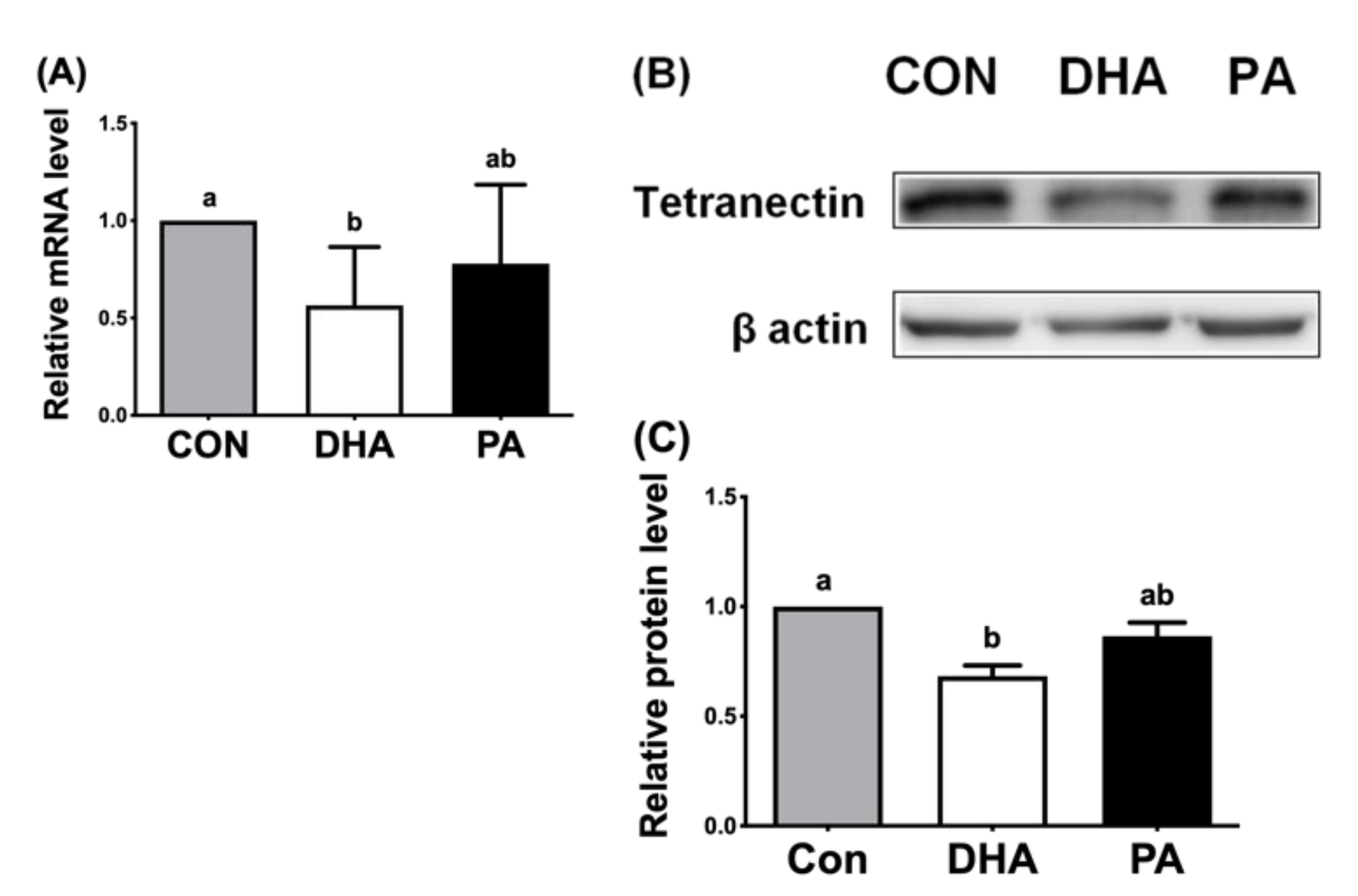
3.3. DHA Reduced Expression of TN in Porcine Adipocytes
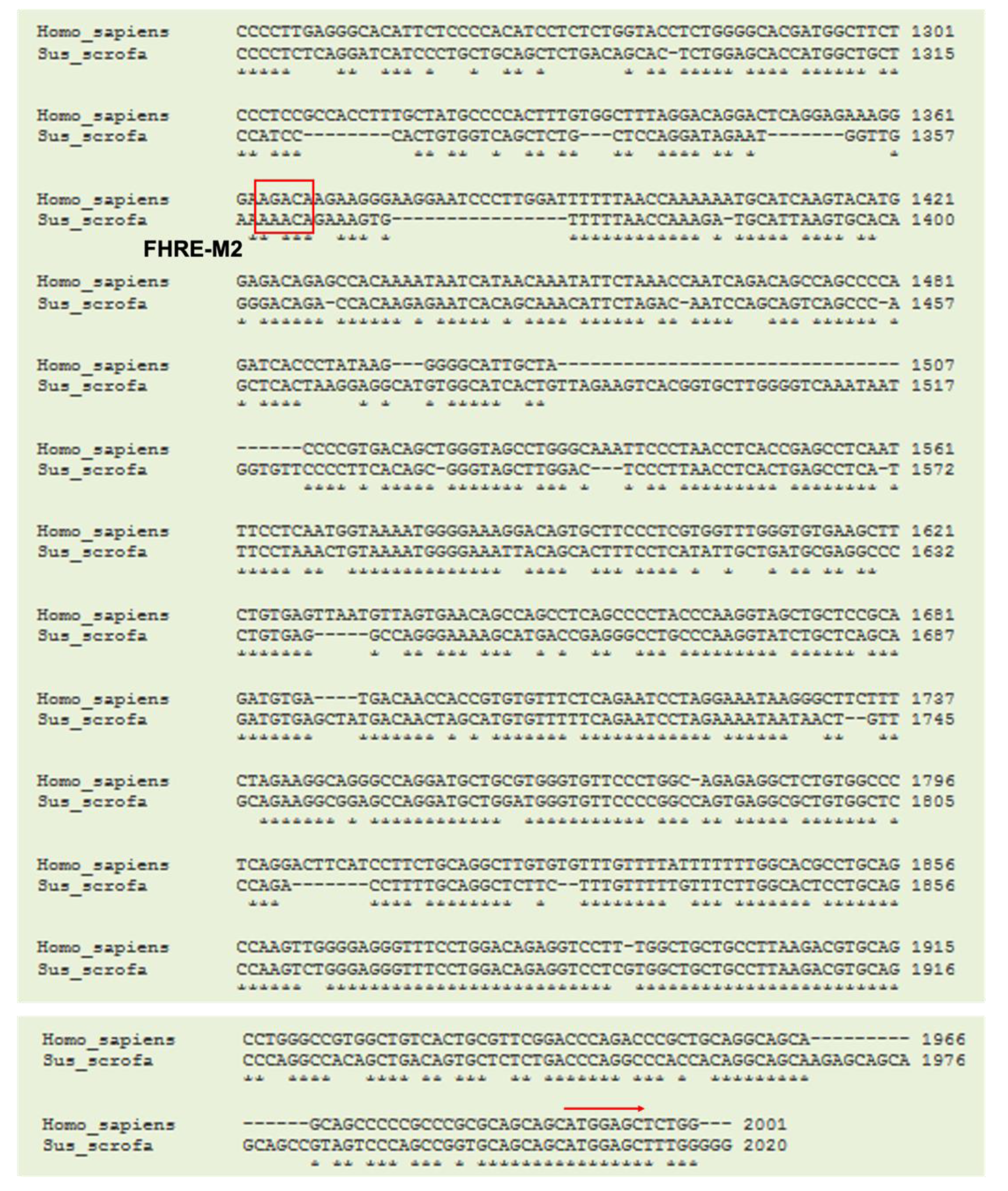
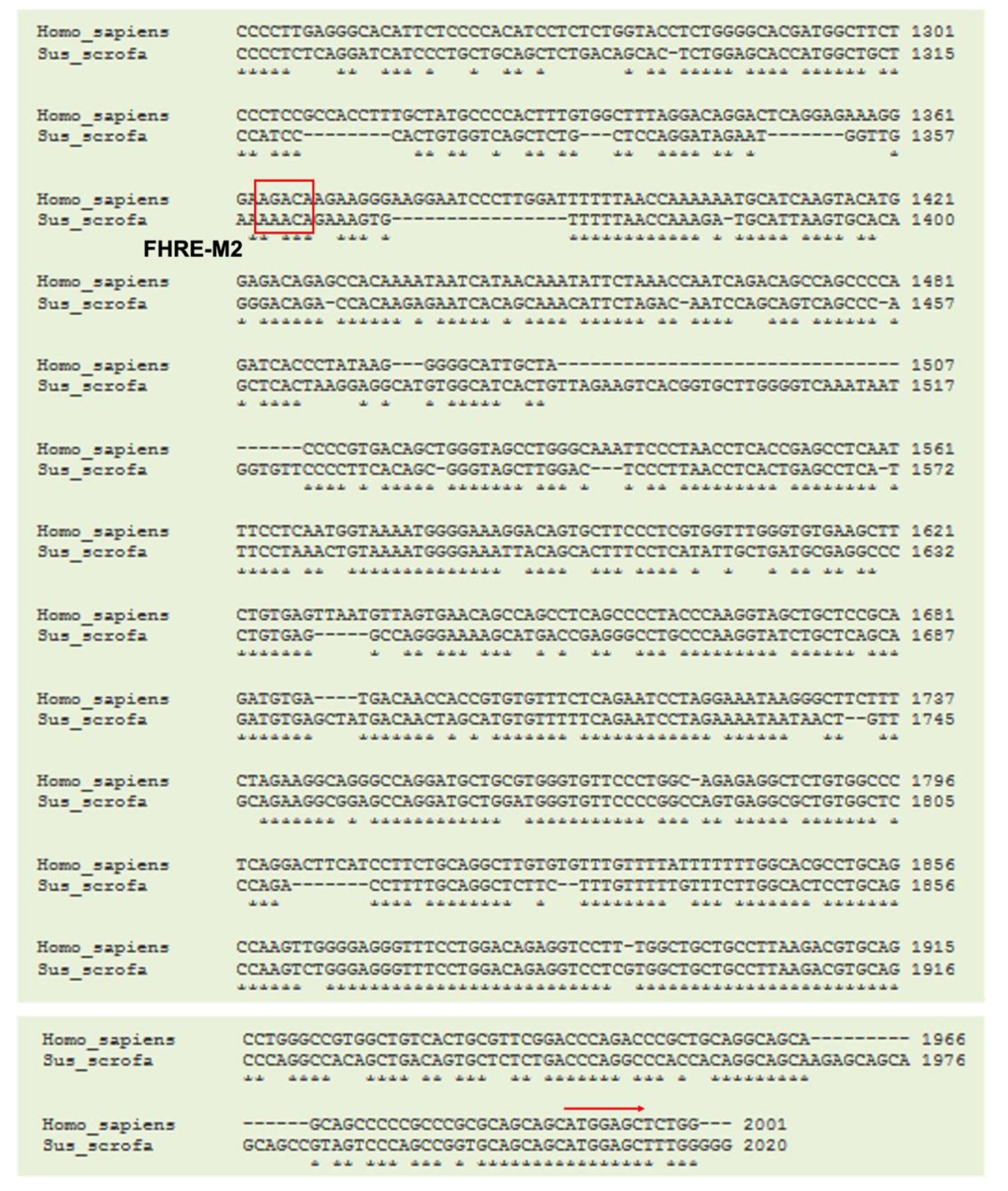
3.4. Cloning of Porcine TN Promoter and Identification of Putative Binding Sites for Transcription Factors on Porcine TN Promoter
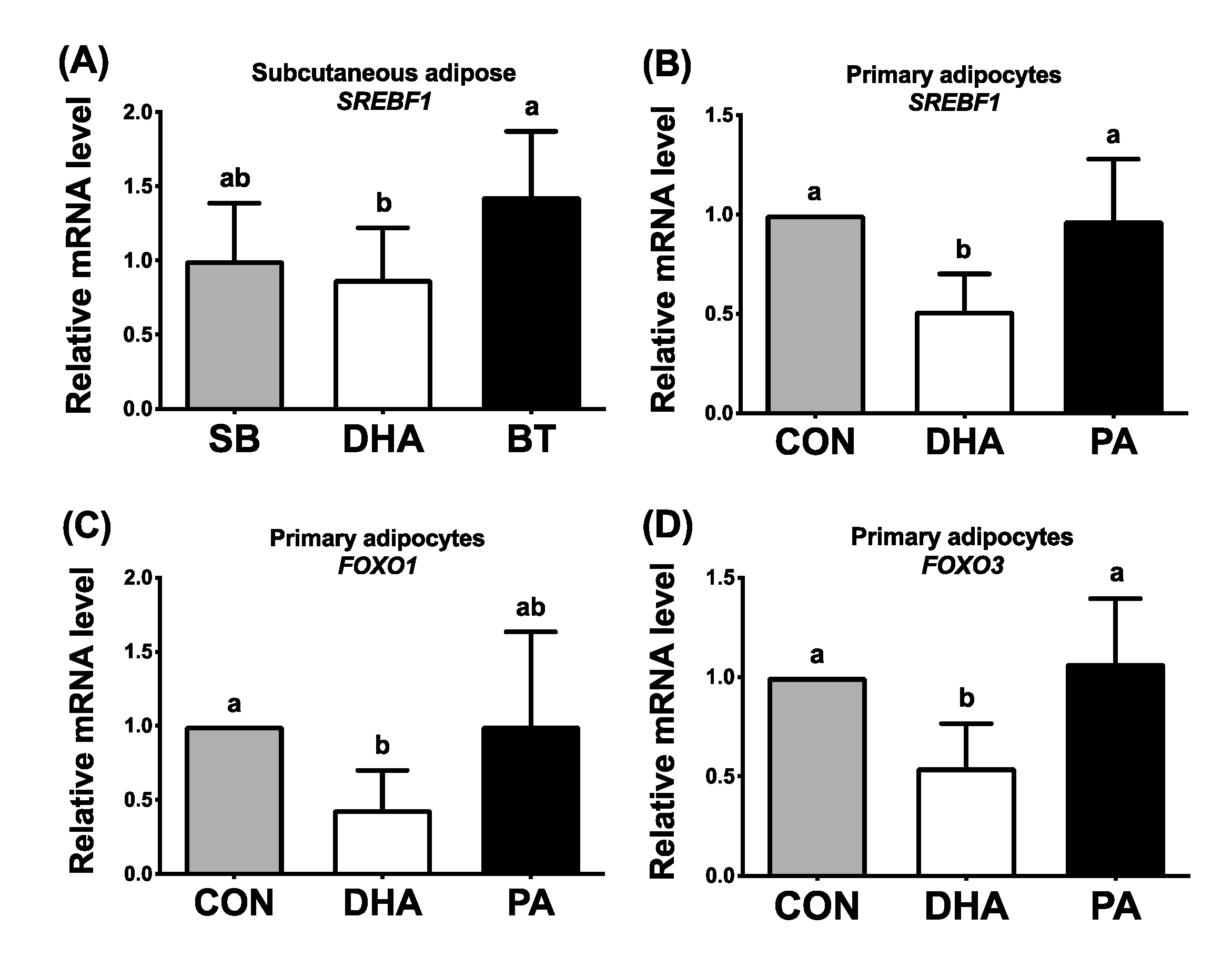
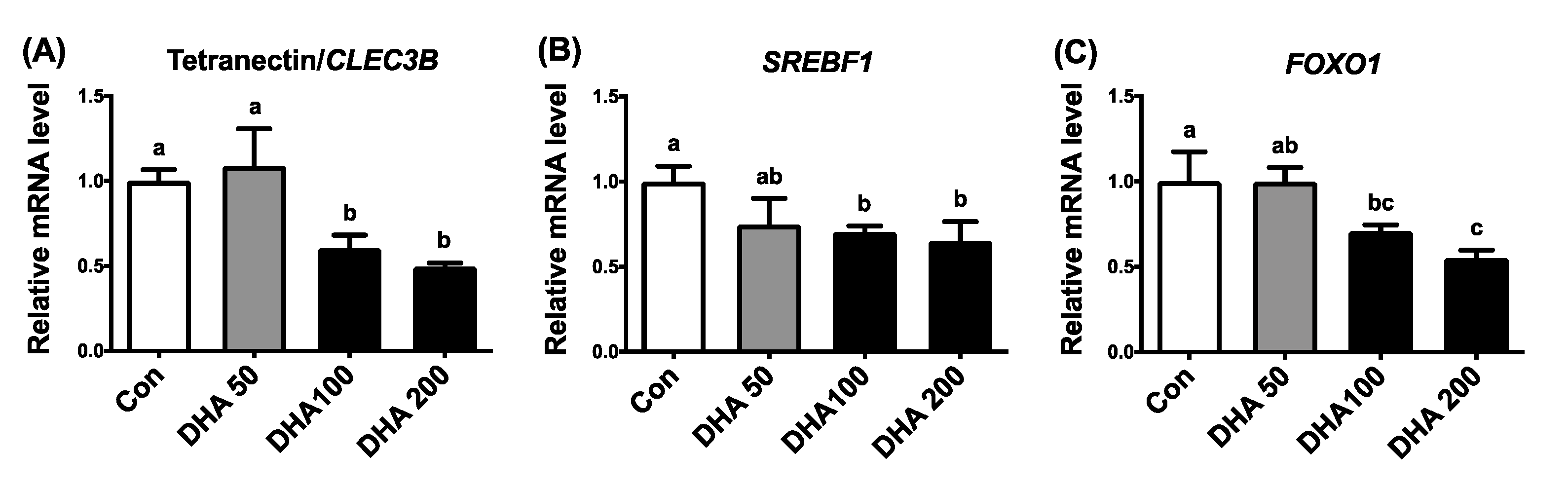
3.5. DHA Repressed Expression of FOXO and SREBP-1c in Porcine Adipose Tissues and Adipocytes
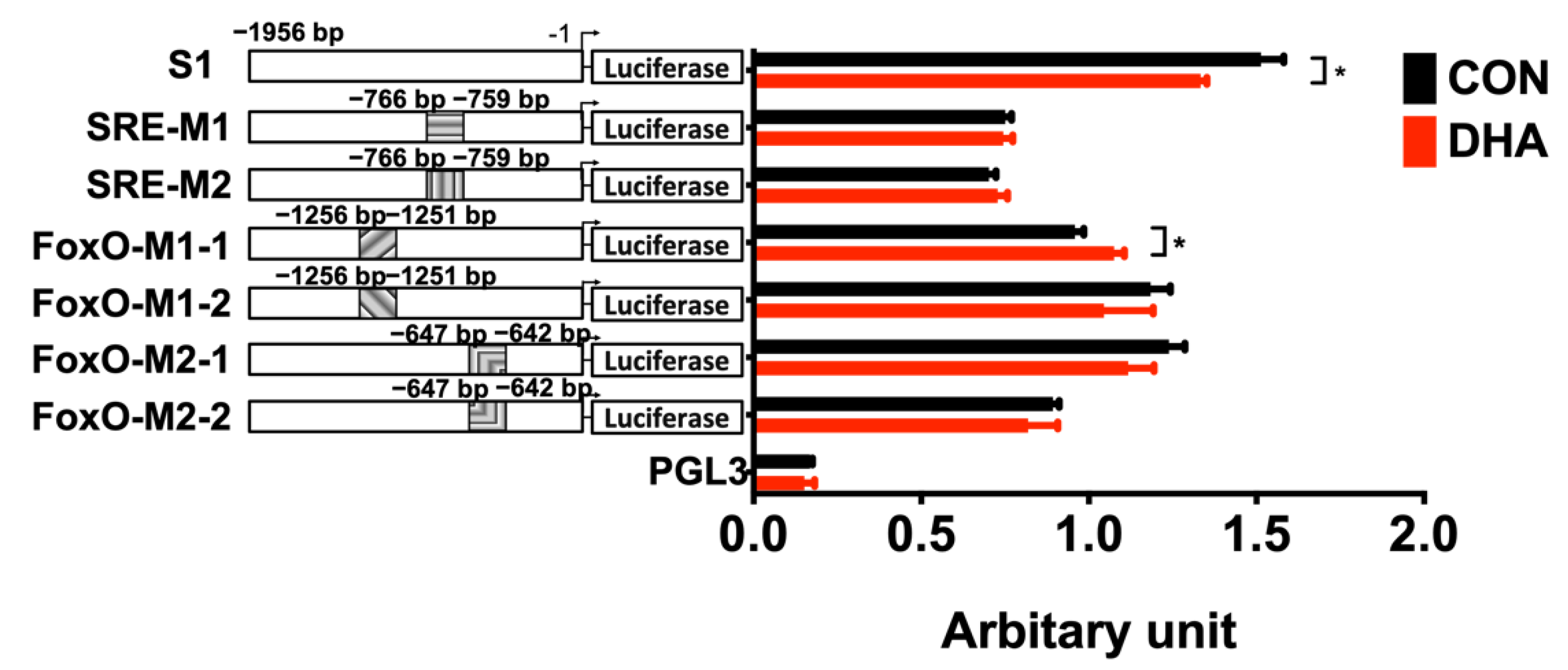
3.6. Transcriptional Suppression of TN Expression by DHA Through SREBP-1c and FoxO in HEK293T Cells
4. Discussion
4.1. Negative Regulation of TN by DHA
4.2. TN Promoter Regulation by DHA
4.3. Implication of TN in Adipogenesis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Popkin, B.M.; Du, S.; Green, W.D.; Beck, M.A.; Algaith, T.; Herbst, C.H.; Alsukait, R.F.; Alluhidan, M.; Alazemi, N.; Shekar, M. Individuals with obesity and COVID-19: A global perspective on the epidemiology and biological relationships. Obes. Rev. 2020, 21, e13128. [Google Scholar] [CrossRef] [PubMed]
- Fryar, C.D.; Carroll, M.D.; Ogden, C.L. Prevalence of Overweight, Obesity, and Extreme Obesity Among Adults Aged 20 and Over: United States, 1960–1962 Through 2013–2014; US National Center for Health Statistics: Atlanta, GA, USA, 2016; pp. 1–6. [Google Scholar]
- Flegal, K.M.; Kruszon-Moran, D.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Trends in Obesity Among Adults in the United States, 2005 to 2014. JAMA 2016, 315, 2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundy, S.M. Metabolic syndrome update. Trends Cardiovasc. Med. 2016, 26, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, I.; Petersen, L.C.; Kluft, C. Purification and characterization of a novel, oligomeric, plasminogen kringle 4 binding protein from human plasma: Tetranectin. Eur. J. Biochem. 1986, 156, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Holtet, T.L.; Graversen, J.H.; Clemmensen, I.; Thogersen, H.C.; Etzerodt, M. Tetranectin, a trimeric plasminogen-binding C-type lectin. Protein Sci. 1997, 6, 1511–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westergaard, U.B.; Andersen, M.H.; Heegaard, C.W.; Fedosov, S.N.; Petersen, T.E. Tetranectin binds hepatocyte growth factor and tissue-type plasminogen activator. Eur. J. Biochem. 2003, 270, 1850–1854. [Google Scholar] [CrossRef]
- Iba, K.; Durkin, M.E.; Johnsen, L.; Hunziker, E.; Damgaard-Pedersen, K.; Zhang, H.; Engvall, E.; Albrechtsen, R.; Wewer, U.M. Mice with a targeted deletion of the tetranectin gene exhibit a spinal deformity. Mol. Cell. Biol. 2001, 21, 7817–7825. [Google Scholar] [CrossRef] [Green Version]
- Wewer, U.M.; Ibaraki, K.; Schjørring, P.; Durkin, M.E.; Young, M.F.; Albrechtsen, R. A potential role for tetranectin in mineralization during osteogenesis. J. Cell Biol. 1994, 127, 1767–1775. [Google Scholar] [CrossRef]
- Jensen, B.A.; Clemmensen, I. Plasma tetranectin is reduced in cancer and related to metastasia. Cancer 1988, 62, 869–872. [Google Scholar] [CrossRef]
- Obrist, P.; Spizzo, G.; Ensinger, C.; Fong, D.; Brunhuber, T.; Schäfer, G.; Varga, M.; Margreiter, R.; Amberger, A.; Gastl, G.; et al. Aberrant tetranectin expression in human breast carcinomas as a predictor of survival. J. Clin. Pathol. 2004, 57, 417–421. [Google Scholar] [CrossRef] [Green Version]
- Høgdall, C.K.; Høgdall, E.V.S.; Hørding, U.; Daugaard, S.; Clemmensen, I.; Nørgaard-Pedersen, B.; Toftager-Larsen, K. Plasma tetranectin and ovarian neoplasms. Gynecol. Oncol. 1991, 43, 103–107. [Google Scholar] [CrossRef]
- H⊘gdall, C.K.; Christensen, L.; Clemmensen, I. The prognostic value of tetranectin immunoreactivity and plasma tetranectin in patients with ovarian cancer. Cancer 1993, 72, 2415–2422. [Google Scholar] [CrossRef]
- Høgdall, C.K.; Sölétormos, G.; Nielsen, D.; Nørgaard-Pedersen, B.; Dombernowsky, P.; Clemmensen, I. Prognostic Value of Serum Tetranectin in Patients with Metastatic Breast Cancer. Acta Oncol. 1993, 32, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Berglund, L.; Petersen, T.E. The gene structure of tetranectin, a plasminogen binding protein. FEBS Lett. 1992, 309, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.Y.; Olefsky, J.M. Omega 3 Fatty Acids and GPR120. Cell Metab. 2012, 15, 564–565. [Google Scholar] [CrossRef] [Green Version]
- Madsen, L.; Petersen, R.K.; Kristiansen, K. Regulation of adipocyte differentiation and function by polyunsaturated fatty acids. Biochim. Biophys. Acta 2005, 1740, 266–286. [Google Scholar] [CrossRef] [Green Version]
- Morrison, R.F.; Farmer, S.R. Hormonal signaling and transcriptional control of adipocyte differentiation. J. Nutr. 2000, 130, 3116s–3121s. [Google Scholar] [CrossRef]
- Chen, Y.J.; Chen, C.C.; Li, T.K.; Wang, P.H.; Liu, L.R.; Chang, F.Y.; Wang, Y.C.; Yu, Y.H.; Lin, S.P.; Mersmann, H.J.; et al. Docosahexaenoic acid suppresses the expression of FoxO and its target genes. J. Nutr. Biochem. 2012, 23, 1609–1616. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Ding, S.-T. Tetranectin Promotes Adipogenesis and Lipogenesis and is Negatively Regulated by Polyunsaturated Fatty Acids in Human Adipocytes. FASEB J. 2012, 26, lb303. [Google Scholar] [CrossRef]
- Chen, Y.J.; Liu, H.Y.; Chang, Y.T.; Cheng, Y.H.; Mersmann, H.J.; Kuo, W.H.; Ding, S.T. Isolation and Differentiation of Adipose-Derived Stem Cells from Porcine Subcutaneous Adipose Tissues. J. Vis. Exp. 2016, 109, e53886. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-W.; Chen, Y.-J.; Yang, J.-T.; Chen, C.-Y.; Ajuwon, K.M.; Chen, S.-E.; Su, N.-W.; Chen, Y.-S.; Mersmann, H.J.; Ding, S.-T. Docosahexaenoic acid increases accumulation of adipocyte triacylglycerol through up-regulation of lipogenic gene expression in pigs. Lipids Health Dis. 2017, 16, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Choi, Y.S.; Lim, S.; Yea, K.; Yoon, J.H.; Jun, D.J.; Ha, S.H.; Kim, J.W.; Kim, J.H.; Suh, P.G.; et al. Comparative analysis of the secretory proteome of human adipose stromal vascular fraction cells during adipogenesis. Proteomics 2010, 10, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.M.; Wang, P.H.; Liu, B.H.; Ding, S.T. The effect of dietary docosahexaenoic acid on the expression of porcine lipid metabolism-related genes. J. Anim. Sci. 2004, 82, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.C.; Chen, C.Y.; Lee, H.S.; Wang, Y.C.; Li, T.K.; Mersamm, H.J.; Ding, S.T.; Wang, P.H. Docosahexaenoic Acid Enhances Hepatic Serum Amyloid A Expression via Protein Kinase A-dependent Mechanism. J. Biol. Chem. 2009, 284, 32239–32247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, S.; Cooper, J.A. Effect of dietary fatty acid composition on substrate utilization and body weight maintenance in humans. Eur. J. Nutr. 2014, 53, 691–710. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Kuo, C.F.; Wang, Y.C.; Ding, S.T. Effect of docosahexaenoic acid and arachidonic acid on the expression of adipocyte determination and differentiation-dependent factor 1 in differentiating porcine adipocytes. J. Anim. Sci. 2005, 83, 1516–1525. [Google Scholar] [CrossRef]
- Fernandez-Alvarez, A.; Alvarez, M.S.; Gonzalez, R.; Cucarella, C.; Muntane, J.; Casado, M. Human SREBP1c expression in liver is directly regulated by peroxisome proliferator-activated receptor alpha (PPARalpha). J. Biol. Chem. 2011, 286, 21466–21477. [Google Scholar] [CrossRef] [Green Version]
- Le Lay, S.; Lefrere, I.; Trautwein, C.; Dugail, I.; Krief, S. Insulin and sterol-regulatory element-binding protein-1c (SREBP-1C) regulation of gene expression in 3T3-L1 adipocytes. Identification of CCAAT/enhancer-binding protein beta as an SREBP-1C target. J. Biol. Chem. 2002, 277, 35625–35634. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Takahashi, M.; Ezaki, O. Fish oil feeding decreases mature sterol regulatory element-binding protein 1 (SREBP-1) by down-regulation of SREBP-1c mRNA in mouse liver. A possible mechanism for down-regulation of lipogenic enzyme mRNAs. J. Biol. Chem. 1999, 274, 25892–25898. [Google Scholar] [CrossRef] [Green Version]
- Begg, D.P.; Woods, S.C. The central insulin system and energy balance. Appet. Control. 2012, 111–129. [Google Scholar] [CrossRef]
- Shimano, H. Sterol regulatory element-binding proteins (SREBPs): Transcriptional regulators of lipid synthetic genes. Prog. Lipid Res. 2001, 40, 439–452. [Google Scholar] [CrossRef]
- Nakae, J.; Biggs, W.H.; Kitamura, T.; Cavenee, W.K.; Wright, C.V.; Arden, K.C.; Accili, D. Regulation of insulin action and pancreatic beta-cell function by mutated alleles of the gene encoding forkhead transcription factor Foxo1. Nat. Genet. 2002, 32, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Nakae, J.; Kitamura, T.; Kitamura, Y.; Biggs, W.H.; Arden, K.C.; Accili, D. The forkhead transcription factor Foxo1 regulates adipocyte differentiation. Dev. Cell 2003, 4, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Kobayashi, M.; Sasaki, T.; Kikuchi, O.; Amano, K.; Kitazumi, T.; Lee, Y.S.; Yokota-Hashimoto, H.; Susanti, V.Y.; Kitamura, Y.I.; et al. Overexpression of FoxO1 in the hypothalamus and pancreas causes obesity and glucose intolerance. Endocrinology 2012, 153, 659–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.J.; Li, P.; Huntley, J.; Chang, J.P.; Arden, K.C.; Olefsky, J.M. FoxO1 haploinsufficiency protects against high-fat diet-induced insulin resistance with enhanced peroxisome proliferator-activated receptor gamma activation in adipose tissue. Diabetes 2009, 58, 1275–1282. [Google Scholar] [CrossRef] [Green Version]
- Nakae, J.; Cao, Y.; Oki, M.; Orba, Y.; Sawa, H.; Kiyonari, H.; Iskandar, K.; Suga, K.; Lombes, M.; Hayashi, Y. Forkhead transcription factor FoxO1 in adipose tissue regulates energy storage and expenditure. Diabetes 2008, 57, 563–576. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; White, M.F. Targeting Forkhead box O1 from the concept to metabolic diseases: Lessons from mouse models. Antioxid. Redox Signal. 2011, 14, 649–661. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Han, S.; Kitamura, T.; Accili, D. Dual role of transcription factor FoxO1 in controlling hepatic insulin sensitivity and lipid metabolism. J. Clin. investig. 2006, 116, 2464–2472. [Google Scholar] [CrossRef] [Green Version]
- Rosenow, A.; Arrey, T.N.; Bouwman, F.G.; Noben, J.-P.; Wabitsch, M.; Mariman, E.C.M.; Karas, M.; Renes, J. Identification of Novel Human Adipocyte Secreted Proteins by Using SGBS Cells. J. Proteome Res. 2010, 9, 5389–5401. [Google Scholar] [CrossRef]
- Lim, J.-M.; Sherling, D.; Teo, C.F.; Hausman, D.B.; Lin, D.; Wells, L. Defining the Regulated Secreted Proteome of Rodent Adipocytes upon the Induction of Insulin Resistance. J. Proteome Res. 2008, 7, 1251–1263. [Google Scholar] [CrossRef]
- Park, J.; Park, J.; Jeong, J.; Cho, K.-H.; Choi, I.; Kim, J. Identification of tetranectin as adipogenic serum protein. Biochem. Biophys. Res. Commun. 2015, 460, 583–588. [Google Scholar] [CrossRef]
- Park, J.; Ryu, D.; Rahman, S.; Kim, J. Adipogenic function of mouse tetranectin and identification of its functional domain. Biochem. Biophys. Res. Commun. 2019, 519, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hunt, D.; Cushman, S.W.; Hess, S. Proteomic characterization of thiazolidinedione regulation of obese adipose secretome in Zucker obese rats. PROTEOMICS–Clin. Appl. 2009, 3, 1099–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bujalska, I.J.; Quinkler, M.; Tomlinson, J.W.; Montague, C.T.; Smith, D.M.; Stewart, P.M. Expression profiling of 11β-hydroxysteroid dehydrogenase type-1 and glucocorticoid-target genes in subcutaneous and omental human preadipocytes. J. Mol. Endocrinol. 2006, 37, 327–340. [Google Scholar] [CrossRef]
- Tokunaga, M.; Inoue, M.; Jiang, Y.; Barnes, R.H.; Buchner, D.A.; Chun, T.-H. Fat depot-specific gene signature and ECM remodeling of Sca1high adipose-derived stem cells. Matrix Biol. 2014, 36, 28–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Jansson, P.A.; Nagaev, I.; Jack, M.M.; Carvalho, E.; Sunnerhagen, K.S.; Cam, M.C.; Cushman, S.W.; Smith, U. Evidence of impaired adipogenesis in insulin resistance. Biochem. Biophys. Res. Commun. 2004, 317, 1045–1051. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense (5′ → 3′) | Antisense (5′ → 3′) | Temp | Accession Number |
|---|---|---|---|---|
| CLEC3B, clone 1 (536–737) | CACCAAGGTGCACATGAAGT | CTCATCCGCCATATCGTTG | 60 ℃ | XM_005669451.3 |
| CLEC3B, clone 2 (377–602) | TGAGACACCTACCCCCAAGA | CTCGCTCGCCTCGTGGAAAGTC | 58 ℃ | XM_005669451.3 |
| CLEC3B, clone 3 (166–408) | GTGCTCTCTGACCCAGGCCCA | CTCGCTCGCCTCGTGGAAAGTC | 58 ℃ | XM_005669451.3 |
| Promoter | Sense (5′ → 3′) | Antisense (5′ → 3′) |
|---|---|---|
| TN-S1 (wild-type) | GCGCGGTACCCTGTCACCACCTCGGACTTT | GCGCAAGCTTGCTGCTGCACCGGCTGGGA |
| TN-SRE site 1-mutant 1 (SRE-M1), first half part | GCGCGGTACCCTGTCACCACCTCGGACTTT | CAGCCTGGTTTTGGGTGTCAGGA |
| TN-SRE site 1-mutant 1 (SRE-M1), second half part | TCCTGACACCCAAAACCAGGCTG | GCGCAAGCTTGCTGCTGCACCGGCTGGGA |
| TN-SRE site 1-mutant 2 (SRE-M2), first half part | GCGCGGTACCCTGTCACCACCTCGGACTTT | CAGCCTGGTGTGGTTTGTCAGGA |
| TN-SRE site 1-mutant 2 (SRE-M2), second half part | TCCTGACAAACCACACCAGGCTG | GCGCAAGCTTGCTGCTGCACCGGCTGGGA |
| TN-FHRE/FoxO site 1-mutant 1 (FoxO-M1-1), first half part | GCGCGGTACCCTGTCACCACCTCGGACTTT | GCAGAAATTGAGCCCAGGACAAGTTGCCTTGCCAG |
| TN-FHRE/FoxO site 1-mutant 1 (FoxO-M1-1), second half part | CTGGCAAGGCAACTTGTCCTGGGCTCAATTTCTGC | GCGCAAGCTTGCTGCTGCACCGGCTGGGA |
| TN-FHRE/FoxO site 1-mutant 2 (FoxO-M1-2), first half part | GCGCGGTACCCTGTCACCACCTCGGACTTT | GCAGAAATTGAGCCCAGGACAAGTCGTCTTGCCAG |
| TN-FHRE/FoxO site 1-mutant 2 (FoxO-M1-2), second half part | CTGGCAAGACGACTTGTCCTGGGCTCAATTTCTGC | GCGCAAGCTTGCTGCTGCACCGGCTGGGA |
| TN-FHRE/FoxO site 2-mutant 1 (FoxO-M2-1), first half part | GCGCGGTACCCTGTCACCACCTCGGACTTT | CACTTTCTGCCTTTCAACCATTCTATCCTGGAGC |
| TN-FHRE/FoxO site 2-mutant 1 (FoxO-M2-1), second half part | GCTCCAGGATAGAATGGTTGAAAGGCAGAAAGTG | GCGCAAGCTTGCTGCTGCACCGGCTGGGA |
| TN-FHRE/FoxO site 2-clone 2 (FoxO-M2-2), first half part | GCGCGGTACCCTGTCACCACCTCGGACTTT | CACTTTCTATCTTTCAACCATTCTATCCTGGAGC |
| TN-FHRE/FoxO site 2-clone 2 (FoxO-M2-2), second half part | GCTCCAGGATAGAATGGTTGAAAGATAGAAAGTG | GCGCAAGCTTGCTGCTGCACCGGCTGGGA |
| Gene | Sense (5′ → 3′) | Antisense (5′ → 3′) | Accession Number |
|---|---|---|---|
| porcine CLEC3B (tetranectin, TN) | GAGGTGGCCCTGCTGAAGGA | CTCGCTCGCCTCGTGGAAAGTC | XM_005669451.2 |
| porcine ACTB (β-actin) | ATCGTGCGGGACATCAAG | AGGAAGGAGGGCTGGAAG | AY550069.1 |
| porcine SREBF1 (SREBP-1c) | GCGACGGTGCCTCTGGTAGT | CGCAAGACGGCGGATTTA | AF102873.1 |
| porcine FOXO1 | AAGAGCGTGCCCTACTTCAA | TTCCTTCATTCTGCACACGA | NM_214014.2 |
| porcine FOXO3 | GGCTGGAAGAACTCTATC | GTACTTGTTGCTGTTGTC | NM_001135959.1 |
| human CLEC3B (tetranectin, TN) | GAGCTCTGGGGGGCCTAC | CTTGGCATTTACAATCTTCTTGG | NM_003278.2 |
| human ACTB (β-actin) | AAGATCAAGATCATTGCTCCTCC | CGGACTCGTCATACTCCTG | NM_001101.3 |
| human SREBF1 (SREBP-1c) | CTGTTGGTGCTCGTCTCCTTGG | CAGCAGGTGACGGATGAGGTTC | NM_001005291.2 |
| human FOXO1 | CGGAATGACCTCATGGATGGA | TAAGTGTAACCTGCTCACTAACCC | NM_002015.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.-T.; Chen, Y.-J.; Huang, C.-W.; Wang, Y.-C.; Mersmann, H.J.; Wang, P.-H.; Ding, S.-T. Docosahexaenoic Acid Suppresses Expression of Adipogenic Tetranectin through Sterol Regulatory Element-Binding Protein and Forkhead Box O Protein in Pigs. Nutrients 2021, 13, 2315. https://doi.org/10.3390/nu13072315
Yang J-T, Chen Y-J, Huang C-W, Wang Y-C, Mersmann HJ, Wang P-H, Ding S-T. Docosahexaenoic Acid Suppresses Expression of Adipogenic Tetranectin through Sterol Regulatory Element-Binding Protein and Forkhead Box O Protein in Pigs. Nutrients. 2021; 13(7):2315. https://doi.org/10.3390/nu13072315
Chicago/Turabian StyleYang, Jui-Ting, Yu-Jen Chen, Chao-Wei Huang, Ya-Chin Wang, Harry J. Mersmann, Pei-Hwa Wang, and Shih-Torng Ding. 2021. "Docosahexaenoic Acid Suppresses Expression of Adipogenic Tetranectin through Sterol Regulatory Element-Binding Protein and Forkhead Box O Protein in Pigs" Nutrients 13, no. 7: 2315. https://doi.org/10.3390/nu13072315
APA StyleYang, J.-T., Chen, Y.-J., Huang, C.-W., Wang, Y.-C., Mersmann, H. J., Wang, P.-H., & Ding, S.-T. (2021). Docosahexaenoic Acid Suppresses Expression of Adipogenic Tetranectin through Sterol Regulatory Element-Binding Protein and Forkhead Box O Protein in Pigs. Nutrients, 13(7), 2315. https://doi.org/10.3390/nu13072315