
Konjac Glucomannan Attenuated Triglyceride Metabolism during Rice Gruel Tolerance Test

 ,
,
Abstract
1. Introduction
2. Materials and Methods
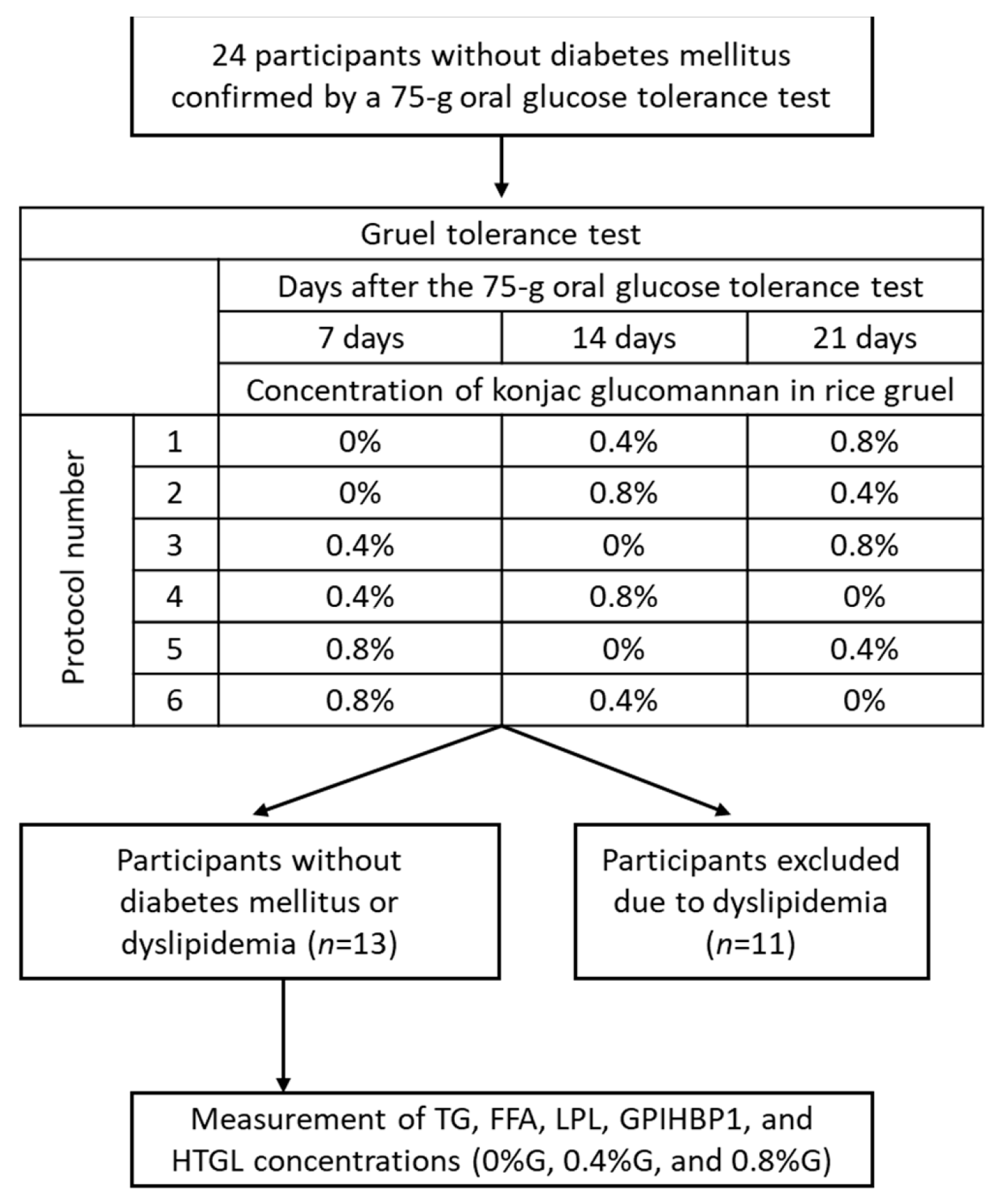
2.1. Participants
2.2. Gruel Test
2.3. Study Design
2.4. Laboratory Assays
2.5. Statistical Analysis
3. Results
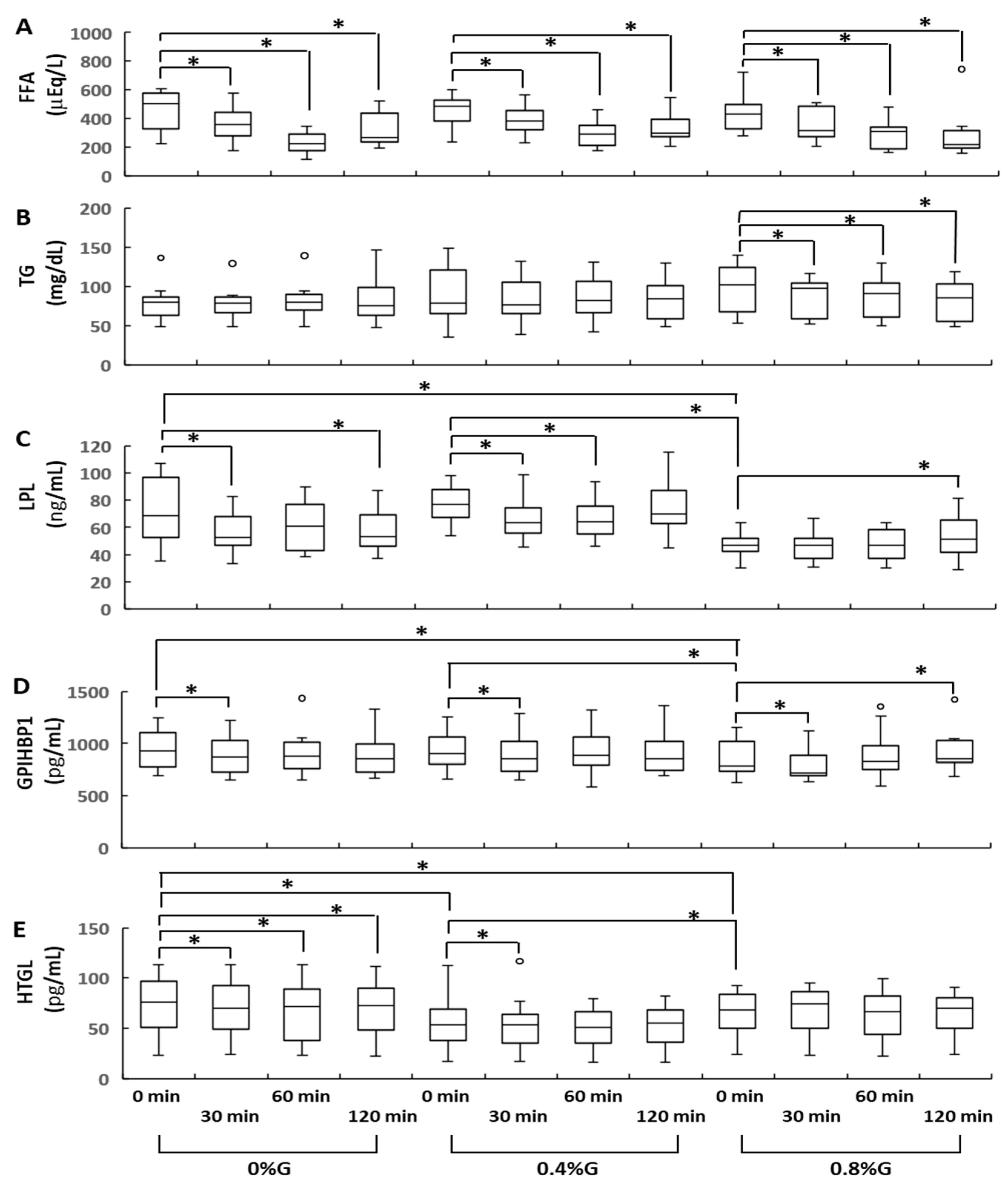
3.1. Changes in Circulating TG Levels
3.2. Changes in Circulating LPL, GPIHBP1, and HTGL Levels
3.3. Circulating Levels of Fasting HTGL and FFA Were Significantly Correlated
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BMI | body mass index |
| ELISA | enzyme-linked immunosorbent assay |
| FFA | free fatty acid |
| GPIHBP1 | glycosylphosphatidylinositol-anchored high-density lipoprotein-binding protein 1 |
| HbA1c | hemoglobin A1c |
| HDL-C | high-density lipoprotein cholesterol |
| HTGL | hepatic triglyceride lipase |
| KGM | konjac glucomannan |
| LDL-C | low-density lipoprotein cholesterol |
| LPL | lipoprotein lipase |
| TG | triglycerides |
| TRL | triglyceride-rich lipoprotein |
| VLDL | very low-density lipoprotein |
| 0%G | rice gruel containing no konjac glucomannan powder |
| 0.4%G | rice gruel supplemented with 0.4% konjac glucomannan powder |
| 0.8%G | rice gruel supplemented with 0.8% konjac glucomannan powder |
References
- Xiong, G.; Cheng, W.; Ye, L.; Du, X.; Zhou, M.; Lin, R.; Geng, S.; Chen, M.; Corke, H.; Cai, Y.Z. Effects of konjac glucomannan on physicochemical properties of myofibrillar protein and surimi gels from grass carp (Ctenopharyngodon idella). Food Chem. 2009, 116, 413–418. [Google Scholar] [CrossRef]
- Behera, S.S.; Ray, R.C. Nutritional and potential health benefits of konjac glucomannan, a promising polysaccharide of elephant foot yam, Amorphophallus konjac K. Koch: A review. Food Rev. Int. 2017, 33, 22–43. [Google Scholar] [CrossRef]
- Zhou, Y.; Cao, H.; Hou, M.; Nirasawa, S.; Tatsumi, E.; Foster, T.J.; Cheng, Y. Effect of konjac glucomannan on physical and sensory properties of noodles made fromlow-protein wheat flour. Food Res. Int. 2013, 51, 879–885. [Google Scholar] [CrossRef]
- Yang, D.; Yuan, Y.; Wang, L.; Wang, X.; Mu, R.; Pang, J.; Xiao, J.; Zheng, Y. A review on konjac glucomannan gels: Microstructure and application. Int. J. Mol. Sci. 2017, 18, 2250. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.S.; Ray, R.C. Konjac glucomannan, a promising polysaccharide of Amorphophallus konjac K. Koch in health care. Int. J. Biol. Macromol. 2016, 92, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Xie, B.J.; Gan, X. Advance in the applications of konjac glucomannan and its derivatives. Carbohydr. Polym. 2005, 60, 27–31. [Google Scholar] [CrossRef]
- Devaraj, R.D.; Reddy, C.K.; Xu, B. Health-promoting effects of konjac glucomannan and its practical applications: A critical review. Int. J. Biol. Macromol. 2019, 126, 273–281. [Google Scholar] [CrossRef]
- Yoshida, A.; Kimura, T.; Tsunekawa, K.; Araki, O.; Ushiki, K.; Ishigaki, H.; Shoho, Y.; Suda, I.; Hiramoto, S.; Murakami, M. Glucomannan inhibits rice gruel-induced increases in plasma glucose and insulin levels. Ann. Nutr. Metab. 2020, 76, 259–267. [Google Scholar] [CrossRef]
- Havel, R.J. Triglyceride-rich lipoproteins and plasma lipid transport. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 9–19. [Google Scholar] [CrossRef]
- Khera, A.V.; Won, H.H.; Peloso, G.M.; O’Dushlaine, C.; Liu, D.; Stitziel, N.O.; Natarajan, P.; Nomura, A.; Emdin, C.A.; Gupta, N.; et al. Association of rare and common variation in the lipoprotein lipase gene with coronary artery disease. JAMA 2017, 317, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Stitziel, N.O.; Stirrups, K.E.; Masca, N.G.; Erdmann, J.; Ferrario, P.G.; König, I.R.; Weeke, P.E.; Webb, T.R.; Auer, P.L.; Schick, U.M.; et al. Coding variation in ANGPTL4, LPL, and SVEP1 and the risk of coronary disease, myocardial infarction genetics and CARDIoGRAM Exome Consortia Investigators. N. Engl. J. Med. 2016, 374, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.S.; Beigneux, A.P.; Barnes, R.H., II; Tu, Y.; Gin, P.; Weinstein, M.M.; Nobumori, C.; Nyrén, R.; Goldberg, I.; Olivecrona, G.; et al. GPIHBP1 is responsible for the entry of lipoprotein lipase into capillaries. Cell Metab. 2010, 12, 42–52. [Google Scholar] [CrossRef]
- Goulbourne, C.N.; Gin, P.; Tatar, A.; Nobumori, C.; Hoenger, A.; Jiang, H.; Grovenor, C.R.; Adeyo, O.; Esko, J.D.; Goldberg, I.J.; et al. The GPIHBP1-LPL complex is responsible for the margination of triglyceride-rich lipoproteins in capillaries. Cell Metab. 2014, 19, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Young, S.G.; Zechner, R. Biochemistry and pathophysiology of intravascular and intracellular lipolysis. Genes Dev. 2013, 27, 459–484. [Google Scholar] [CrossRef]
- Fong, L.G.; Young, S.G.; Beigneux, A.P.; Bensadoun, A.; Oberer, M.; Jiang, H.; Ploug, M. GPIHBP1 and plasma triglyceride metabolism. Trends Endocrinol. Metab. 2016, 27, 455–469. [Google Scholar] [CrossRef]
- Tsuzaki, K.; Kotani, K.; Yamada, K.; Sakane, N. Fasting lipoprotein lipase protein levels can predict a postmeal increment of triglyceride levels in fasting normohypertriglyceridemic subjects. J. Clin. Lab. Anal. 2016, 30, 404–407. [Google Scholar] [CrossRef]
- Ishiyama, N.; Sakamaki, K.; Shimomura, Y.; Kotani, K.; Tsuzaki, K.; Sakane, N.; Miyashita, K.; Fukamachi, I.; Kobayashi, J.; Stanhope, K.L.; et al. Lipoprotein lipase does not increase significantly in the postprandial plasma. Clin. Chim. Acta 2017, 464, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Committee of the Japan Diabetes Society on the Diagnostic Criteria of Diabetes Mellitus; Seino, Y.; Nanjo, K.; Tajima, N.; Kadowaki, T.; Kashiwagi, A.; Araki, E.; Ito, C.; Inagaki, N.; Iwamoto, Y.; et al. Report of the Committee on the classification and diagnostic criteria of diabetes mellitus. J. Diabetes Investig. 2010, 1, 212–228. [Google Scholar] [CrossRef]
- Matsumoto, R.; Tsunekawa, K.; Shoho, Y.; Yanagawa, Y.; Kotajima, N.; Matsumoto, S.; Araki, O.; Kimura, T.; Nakajima, K.; Murakami, M. Association between skeletal muscle mass and serum concentrations of lipoprotein lipase, GPIHBP1, and hepatic triglyceride lipase in young Japanese men. Lipids Health Dis. 2019, 18, 84. [Google Scholar] [CrossRef]
- Vuksan, V.; Sievenpiper, J.L.; Xu, Z.; Wong, E.Y.Y.; Jenkins, A.L.; Beljan-Zdravkovic, U.; Leiter, L.A.; Josse, R.G.; Stavro, M.P. Konjac-mannan and American ginsing: Emerging alternative therapies for type 2 diabetes mellitus. J. Am. Coll. Nutr. 2001, 20, 370S–381S; discussion 381S. [Google Scholar] [CrossRef]
- Ebihara, K.; Schneeman, B.O. Interaction of bile acids, phospholipids, cholesterol and triglyceride with dietary fibers in the small intestine of rats. J. Nutr. 1989, 119, 1100–1106. [Google Scholar] [CrossRef]
- Kiriyama, S.; Enishi, A.; Yura, K. Inhibitory effect of konjac mannan on bile acid transport in the everted sacs from rat ileum. J. Nutr. 1974, 104, 69–78. [Google Scholar] [CrossRef]
- Shen, Q.; Zhao, L.; Tuohy, K.M. High-level dietary fibre up-regulates colonic fermentation and relative abundance of saccharolytic bacteria within the human faecal microbiota in vitro. Eur. J. Nutr. 2012, 51, 693–705. [Google Scholar] [CrossRef]
- Hozumi, T.; Yoshida, M.; Ishida, Y.; Mimoto, H.; Sawa, J.; Doi, K.; Kazumi, T. Long-term effects of dietary fiber supplementation on serum glucose and lipoprotein levels in diabetic rats fed a high cholesterol diet. Endocr. J. 1995, 42, 187–192. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Anderson, J.W. Dietary fiber, lipids and atherosclerosis. Am. J. Cardiol. 1987, 60, 17G–22G. [Google Scholar] [CrossRef]
- Anderson, J.W.; Akanji, A.O. Dietary fiber—An overview. Diabetes Care 1991, 14, 1126–1131. [Google Scholar] [CrossRef]
- Chen, W.J.; Anderson, J.W.; Jennings, D. Propionate may mediate the hypocholesterolemic effects of certain soluble plant fibers in cholesterol-fed rats. Proc. Soc. Exp. Biol. Med. 1984, 175, 215–218. [Google Scholar] [CrossRef]
- Young, S.G.; Fong, L.G.; Beigneux, A.P.; Allan, C.M.; He, C.; Jiang, H.; Nakajima, K.; Meiyappan, M.; Birrane, G.; Ploug, M. GPIHBP1 and lipoprotein lipase, partners in plasma triglyceride metabolism. Cell Metab. 2019, 30, 51–65. [Google Scholar] [CrossRef]
- Shirakawa, T.; Nakajima, K.; Shimomura, Y.; Kobayashi, J.; Stanhope, K.; Havel, P.; Machida, T.; Sumino, H.; Murakami, M. Comparison of the effect of post-heparin and pre-heparin lipoprotein lipase and hepatic triglyceride lipase on remnant lipoprotein metabolism. Clin. Chim. Acta 2015, 440, 193–200. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shirakawa, T.; Nakajima, K.; Yatsuzuka, S.; Shimomura, Y.; Kobayashi, J.; Machida, T.; Sumino, H.; Murakami, M. The role of circulating lipoprotein lipase and adiponectin on the particle size of remnant lipoproteins in patients with diabetes mellitus and metabolic syndrome. Clin. Chim. Acta 2015, 440, 123–132. [Google Scholar] [CrossRef]
- Muraba, Y.; Koga, T.; Shimomura, Y.; Ito, Y.; Hirao, Y.; Kobayashi, J.; Kimura, T.; Nakajima, K.; Murakami, M. The role of plasma lipoprotein lipase, hepatic lipase and GPIHBP1 in the metabolism of remnant lipoproteins and small dense LDL in patients with coronary artery disease. Clin. Chim. Acta 2018, 476, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Machida, T.; Imamura, S.; Kawase, D.; Miyashita, K.; Fukamachi, I.; Maeda, M.; Muraba, Y.; Koga, T.; Kobayashi, J.; et al. An automated method for measuring lipoprotein lipase and hepatic triglyceride lipase activities in post-heparin plasma. Clin. Chim. Acta 2018, 487, 54–59. [Google Scholar] [CrossRef]
- DiMarco, N.M.; Beitz, D.C.; Whitehurst, G.B. Effect of fasting on free fatty acid, glycerol and cholesterol concentrations in blood plasma and lipoprotein lipase activity in adipose tissue of cattle. J. Anim. Sci. 1981, 52, 75–82. [Google Scholar] [CrossRef]
- Nelson, R.H.; Edgerton, D.S.; Basu, R.; Roesner, J.C.; Cherrington, A.D.; Miles, J.M. Triglyceride uptake and lipoprotein lipase-generated fatty acid spillover in the splanchnic bed of dogs. Diabetes 2007, 56, 1850–1855. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kagawa, K.; Matsutaka, H.; Fukuhama, C.; Watanabe, Y.; Fujino, H. Globin digest, acidic protease hydrolysate, inhibits dietary hypertriglyceridemia and Val-Val-Tyr-Pro, one of its constituents, possesses most superior effect. Life Sci. 1996, 58, 1745–1755. [Google Scholar] [CrossRef]
- Peterson, J.; Bihain, B.E.; Bengtsson-Olivecrona, G.; Deckelbaum, R.J.; Carpentier, Y.A.; Olivecrona, T. Fatty acid control of lipoprotein lipase: A link between energy metabolism and lipid transport. Proc. Natl. Acad. Sci. USA 1990, 87, 909–913. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| All Participants (n = 13) | |
|---|---|
| Age (years) | 47 (41–50) |
| Weight (kg) | 73.1 (70.3–80.0) |
| BMI (kg/m2) | 24.9 (23.2–26.5) |
| Systolic blood pressure (mmHg) | 125.0 (120–134.8) |
| Diastolic blood pressure (mmHg) | 77.0 (73.5–80.5) |
| Fasting plasma glucose (mg/dL) | 105.0 (100–107.8) |
| Fasting plasma insulin (U/mL) | 5.0 (4.3–6.0) |
| HbA1c (%) | 5.5 (5.3–5.6) |
| LDL-C (mg/dL) | 114 (89.3–134.8) |
| HDL-C (mg/dL) | 50.0 (41.0–56.0) |
| TG (mg/dL) | 73.0 (61.3–87.5) |
| FFA (μEq/L) | 506.0 (330–559) |
| LPL (ng/mL) | 69 (52.9–94.0) |
| GPIHBP1 (pg/mL) | 930.0 (778.3–1020.6) |
| HTGL (pg/mL) | 76.0 (51.5–96.6) |
| FFA | TG | LPL | GPIHBP1 | HTGL | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | r | p | |
| FFA | −0.094 | 0.569 | 0.193 | 0.240 | 0.149 | 0.364 | −0.358 | 0.025 * | ||
| TG | −0.094 | 0.569 | −0.238 | 0.144 | −0.269 | 0.098 | 0.069 | 0.677 | ||
| LPL | 0.193 | 0.240 | −0.238 | 0.144 | 0.142 | 0.389 | −0.115 | 0.487 | ||
| GPIHBP1 | 0.149 | 0.364 | −0.269 | 0.098 | 0.142 | 0.389 | 0.015 | 0.930 | ||
| HTGL | −0.358 | 0.025 * | 0.069 | 0.677 | −0.115 | 0.487 | 0.014 | 0.930 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagasawa, T.; Kimura, T.; Yoshida, A.; Tsunekawa, K.; Araki, O.; Ushiki, K.; Ishigaki, H.; Shoho, Y.; Suda, I.; Hiramoto, S.; et al. Konjac Glucomannan Attenuated Triglyceride Metabolism during Rice Gruel Tolerance Test. Nutrients 2021, 13, 2191. https://doi.org/10.3390/nu13072191
Nagasawa T, Kimura T, Yoshida A, Tsunekawa K, Araki O, Ushiki K, Ishigaki H, Shoho Y, Suda I, Hiramoto S, et al. Konjac Glucomannan Attenuated Triglyceride Metabolism during Rice Gruel Tolerance Test. Nutrients. 2021; 13(7):2191. https://doi.org/10.3390/nu13072191
Chicago/Turabian StyleNagasawa, Takumi, Takao Kimura, Akihiro Yoshida, Katsuhiko Tsunekawa, Osamu Araki, Kazumi Ushiki, Hirotaka Ishigaki, Yoshifumi Shoho, Itsumi Suda, Suguru Hiramoto, and et al. 2021. "Konjac Glucomannan Attenuated Triglyceride Metabolism during Rice Gruel Tolerance Test" Nutrients 13, no. 7: 2191. https://doi.org/10.3390/nu13072191
APA StyleNagasawa, T., Kimura, T., Yoshida, A., Tsunekawa, K., Araki, O., Ushiki, K., Ishigaki, H., Shoho, Y., Suda, I., Hiramoto, S., & Murakami, M. (2021). Konjac Glucomannan Attenuated Triglyceride Metabolism during Rice Gruel Tolerance Test. Nutrients, 13(7), 2191. https://doi.org/10.3390/nu13072191