
Dietary Avian Proteins Are Comparable to Soybean Proteins on the Atherosclerosis Development and Fatty Liver Disease in Apoe-Deficient Mice

 , ,
, ,  and
and
Abstract

1. Introduction
2. Material and Methods
2.1. Animals and Diets
2.2. Dietary Characterization
2.3. Plasma Determinations
2.4. Reactive Oxygen Species (ROS) Content in Lipoproteins
2.5. Evaluation of Atherosclerotic Lesions
2.6. Hepatic Histological Analyses
2.7. Statistical Analyses
3. Results
3.1. Composition of Experimental Diets
3.2. Somatometric Analyses
3.3. Plasma Parameters
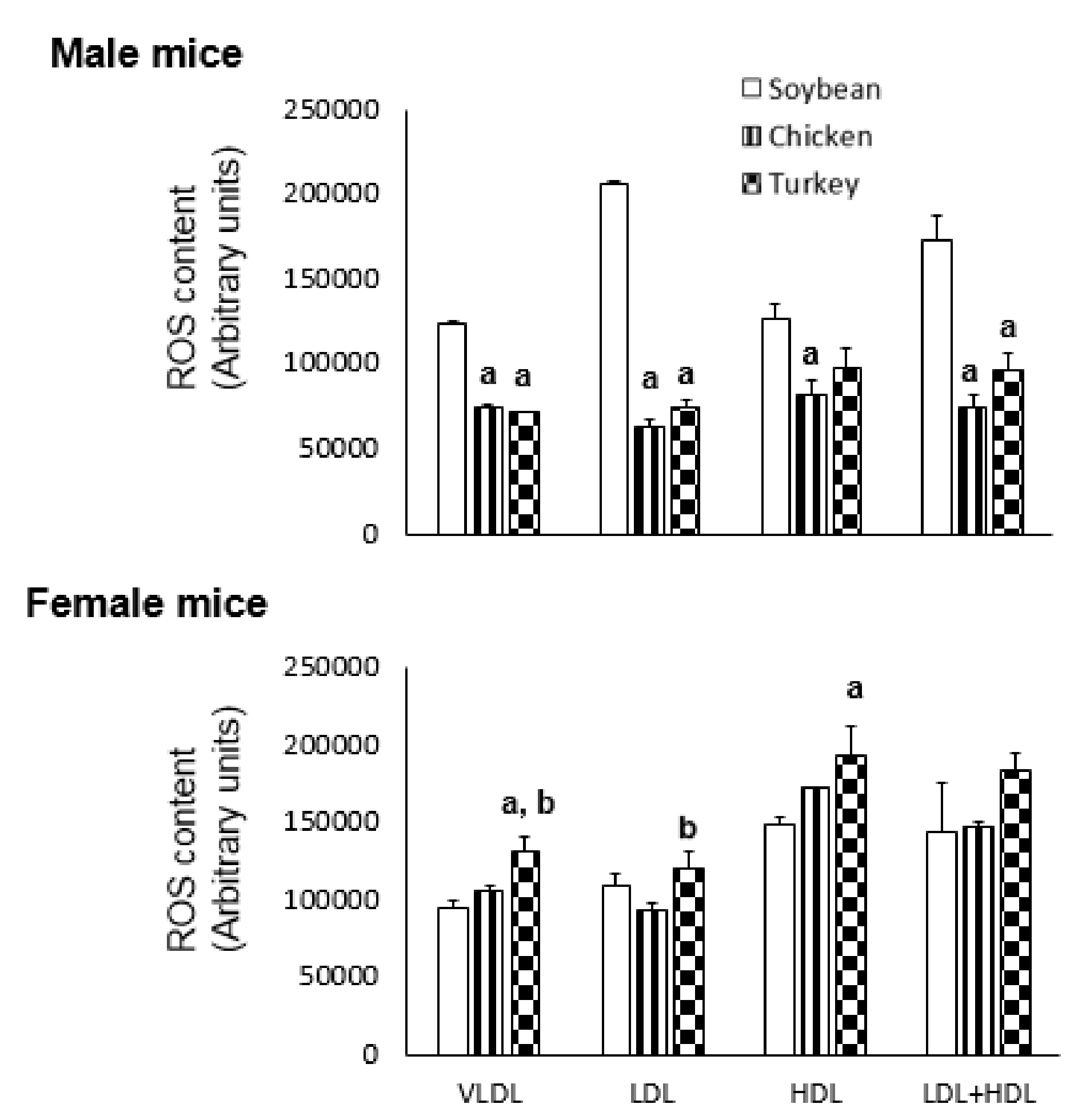
3.4. ROS
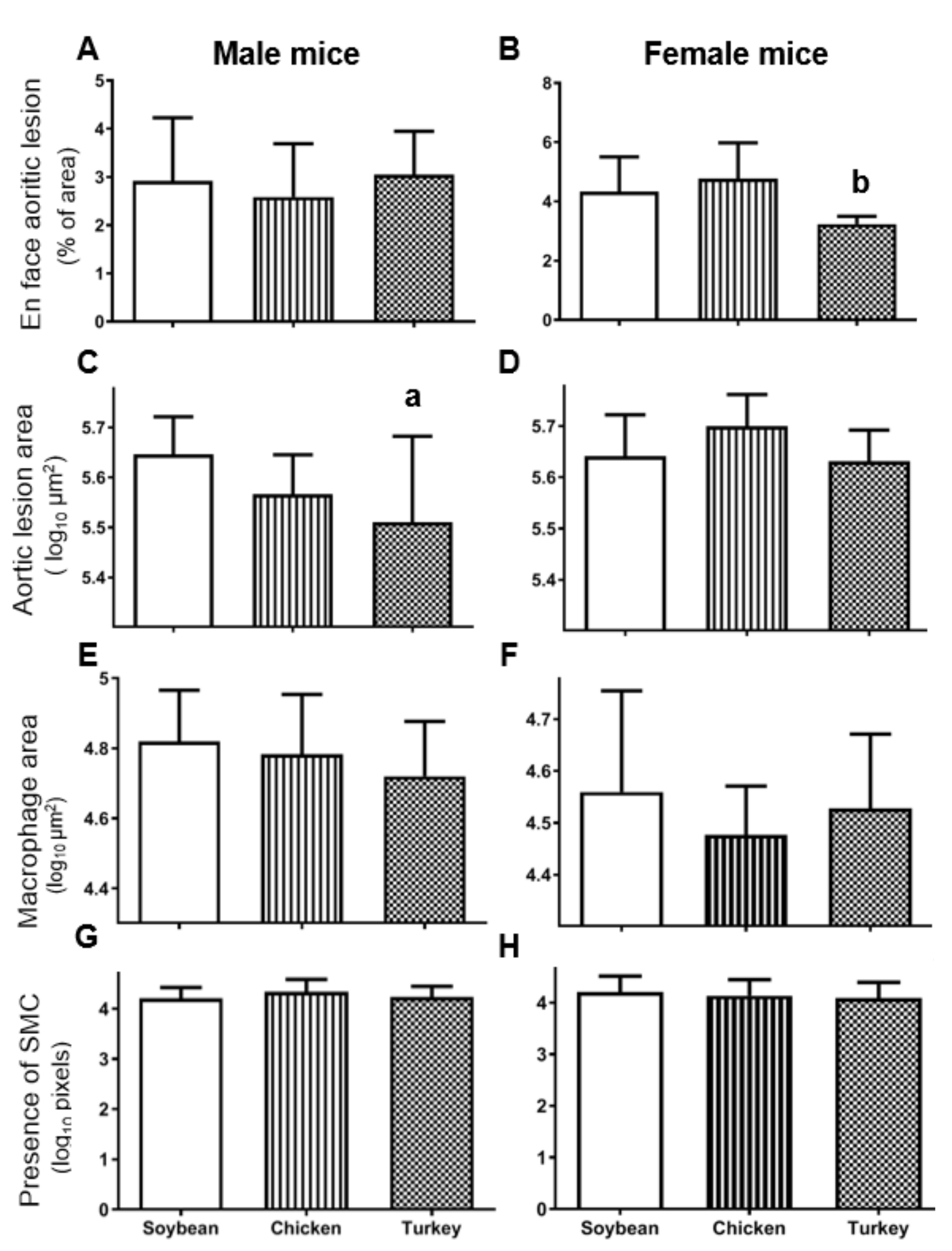
3.5. Aortic Atherosclerotic Lesions
3.6. Hepatic Steatosis
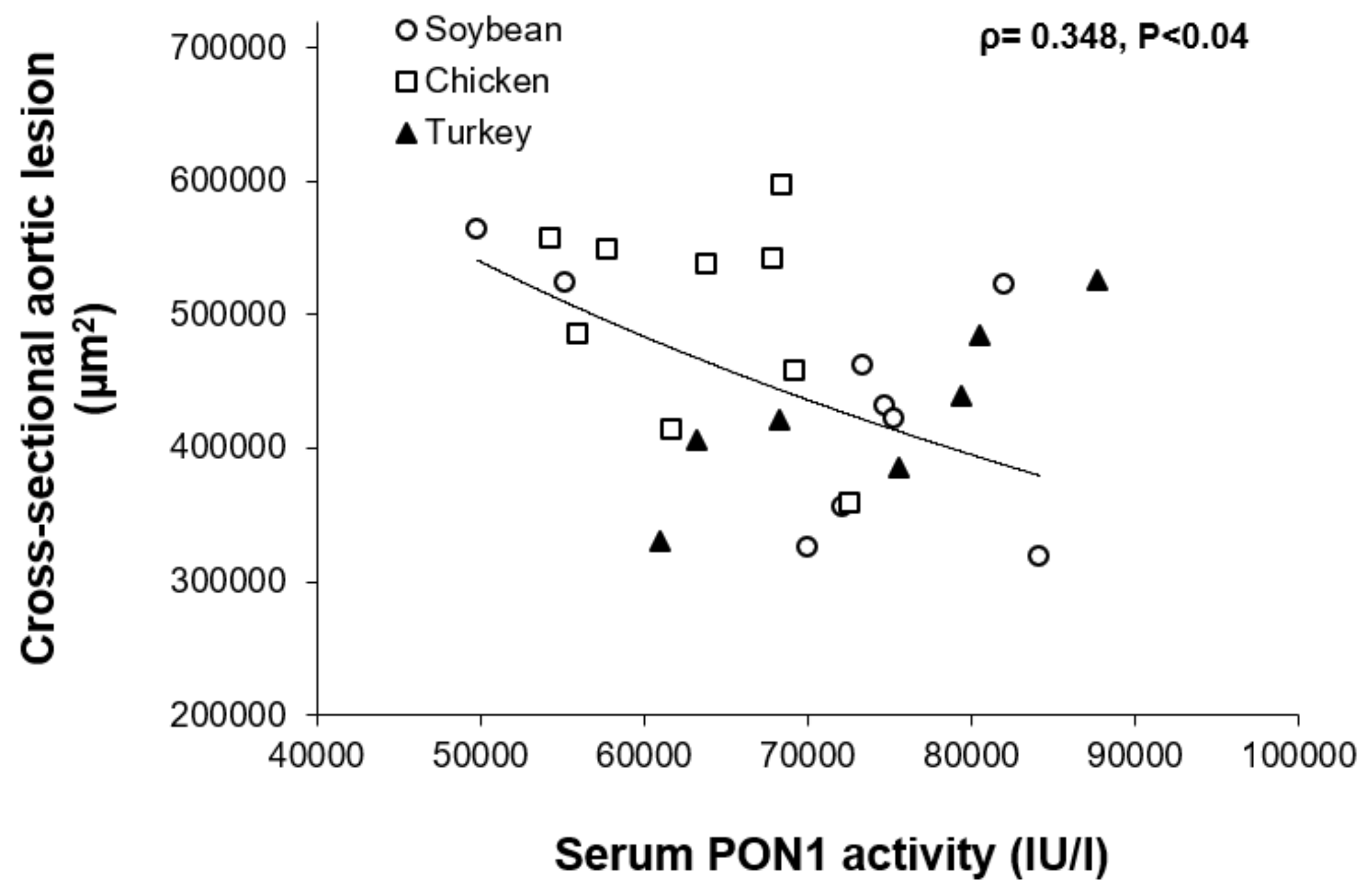
3.7. Association Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APOA1 | apolipoprotein A1 |
| BCAA | branched-chain amino acids |
| FPLC | fast performance liquid chromatography |
| HDL | high density lipoproteins |
| LDL | low density lipoproteins |
| NEFA | non-esterified fatty acids |
| PON1 | paraoxonase |
| ROS | reactive oxygen species |
| VLDL | very low density lipoproteins |
References
- United States Department of Agriculture; U.S. Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025, 9th ed. Available online: https://www.dietaryguidelines.gov (accessed on 3 March 2021).
- FAO. Dietary protein quality evaluation in human nutrition. Report of an FAQ Expert Consultation. FAO Food Nutr. Pap. 2013, 92, 1–66. [Google Scholar]
- Millward, D.J. Identifying recommended dietary allowances for protein and amino acids: A critique of the 2007 WHO/FAO/UNU report. Br. J. Nutr. 2012, 108 (Suppl. S2), S3–S21. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Laboratory Animals, 4th ed.; The National Academies Press: Washington, DC, USA, 1995; p. 192. [Google Scholar]
- Wolfson, R.L.; Sabatini, D.M. The Dawn of the Age of Amino Acid Sensors for the mTORC1 Pathway. Cell Metab. 2017, 26, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.Y.; Jiang, H.J.; Li, L.; Chen, F.W.; Liu, Y.X.; Zhou, M.Y.; Wang, J.; Jiang, J.J.; Li, X.Y.; Fan, X.M.; et al. Branched-Chain Amino Acid Catabolism Promotes Thrombosis Risk by Enhancing Tropomodulin-3 Propionylation in Platelets. Circulation 2020, 142, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Zhenyukh, O.; González-Amor, M.; Rodrigues-Diez, R.R.; Esteban, V.; Ruiz-Ortega, M.; Salaices, M.; Mas, S.; Briones, A.M.; Egido, J. Branched-chain amino acids promote endothelial dysfunction through increased reactive oxygen species generation and inflammation. J. Cell. Mol. Med. 2018, 22, 4948–4962. [Google Scholar] [CrossRef] [PubMed]
- Lichtman, A.H.; Clinton, S.K.; Iiyama, K.; Connelly, P.W.; Libby, P.; Cybulsky, M.I. Hyperlipidemia and atherosclerotic lesion development in LDL receptor-deficient mice fed defined semipurified diets with and without cholate. Arter. Thromb. Vasc. Biol. 1999, 19, 1938–1944. [Google Scholar] [CrossRef]
- Osada, J.; Joven, J.; Maeda, N. The value of apolipoprotein E knockout mice for studying the effects of dietary fat and cholesterol on atherogenesis. Curr. Opin. Lipidol. 2000, 11, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Marcos, L.V.; Sancho-Knapik, S.; Gabas-Rivera, C.; Barranquero, C.; Gascon, S.; Romanos, E.; Martinez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Arnal, C.; et al. Pgc1a is responsible for the sex differences in hepatic Cidec/Fsp27beta mRNA expression in hepatic steatosis of mice fed a Western diet. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E249–E261. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Tsuda, Y.; Sakono, M.; Imaizumi, K. Dietary Soy Protein Isolate, Compared with Casein, Reduces Atherosclerotic Lesion Area in Apolipoprotein E–Deficient Mice. J. Nutr. 1998, 128, 1884–1889. [Google Scholar] [CrossRef]
- Adams, M.R.; Golden, D.L.; Anthony, M.S.; Register, T.C.; Williams, J.K. The inhibitory effect of soy protein isolate on atherosclerosis in mice does not require the presence of LDL receptors or alteration of plasma lipoproteins. J. Nutr. 2002, 132, 43–49. [Google Scholar] [CrossRef]
- Adams, M.R.; Golden, D.L.; Franke, A.A.; Potter, S.M.; Smith, H.S.; Anthony, M.S. Inhibitory effects of dietary 7S globulin on atherosclerosis in mice exceed those of whole soy protein isolate and are independent of LDL receptors and plasma lipoproteins. J. Nutr. 2004, 134, 1273s. [Google Scholar]
- Adams, M.R.; Golden, D.L.; Williams, J.K.; Franke, A.A.; Register, T.C.; Kaplan, J.R. Soy protein containing isoflavones reduces the size of atherosclerotic plaques without affecting coronary artery reactivity in adult male monkeys. J. Nutr. 2005, 135, 2852–2856. [Google Scholar] [CrossRef]
- Sato, M.; Sato, H.; Ogawa, A.; Nomura, R.; Takashima, S.; Bang, H.J.; Matsuoka, H.; Imaizumi, K. Antiatherogenic effect of isoflavones in ovariectomized apolipoprotein e-deficient mice. J. Agric. Food Chem. 2007, 55, 8967–8971. [Google Scholar] [CrossRef]
- Sawashita, N.; Naemura, A.; Shimizu, M.; Morimatsu, F.; Ijiri, Y.; Yamamoto, J. Effect of dietary vegetable and animal proteins on atherothrombosis in mice. Nutrition 2006, 22, 661–667. [Google Scholar] [CrossRef]
- Zhang, Y.; Kouguchi, T.; Shimizu, K.; Sato, M.; Takahata, Y.; Morimatsu, F. Chicken collagen hydrolysate reduces proinflammatory cytokine production in C57BL/6.KOR-ApoEshl mice. J. Nutr. Sci. Vitaminol. 2010, 56, 208–210. [Google Scholar] [CrossRef]
- Ishihara, K.; Fukuchi, Y.; Mizunoya, W.; Mita, Y.; Fukuya, Y.; Fushiki, T.; Yasumoto, K. Amino acid composition of soybean protein increased postprandial carbohydrate oxidation in diabetic mice. Biosci. Biotechnol. Biochem. 2003, 67, 2505–2511. [Google Scholar] [CrossRef]
- Zhao, G.P.; Cui, H.X.; Liu, R.R.; Zheng, M.Q.; Chen, J.L.; Wen, J. Comparison of breast muscle meat quality in 2 broiler breeds. Poult. Sci. 2011, 90, 2355–2359. [Google Scholar] [CrossRef]
- Chen, Y.; Qiao, Y.; Xiao, Y.; Chen, H.; Zhao, L.; Huang, M.; Zhou, G. Differences in Physicochemical and Nutritional Properties of Breast and Thigh Meat from Crossbred Chickens, Commercial Broilers, and Spent Hens. Asian-Australas. J. Anim. Sci. 2016, 29, 855–864. [Google Scholar] [CrossRef]
- Domínguez, R.; Echegaray, N.; Gálvez, F.; Pérez-Santaescolática, C.; Pateiro, M.; Franco, D.; Lorenzo, J.M. Effect of sex on amino acid composition of turkey meat. In Proceedings of the 63rd International Congress of Meat Science and Technology, Cork, Ireland, 13–18 August 2017; pp. 846–847. [Google Scholar]
- Reeves, P.G.; Rossow, K.L.; Lindlauf, J. Development and Testing of the Ain-93 Purified Diets for Rodents—Results on Growth, Kidney Calcification and Bone Mineralization in Rats and Mice. J. Nutr. 1993, 123, 1923–1931. [Google Scholar] [CrossRef]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern. Z. Anal. Chem. 1883, 22, 366–383. [Google Scholar] [CrossRef]
- Commission, E. Commission Regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union 2009, 52, 1. [Google Scholar]
- Navarro, M.A.; Carpintero, R.; Acin, S.; Arbones-Mainar, J.M.; Calleja, L.; Carnicer, R.; Surra, J.C.; Guzman-Garcia, M.A.; Gonzalez-Ramon, N.; Iturralde, M.; et al. Immune-regulation of the apolipoprotein A-I/C-III/A-IV gene cluster in experimental inflammation. Cytokine 2005, 31, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Gabas-Rivera, C.; Barranquero, C.; Martinez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Osada, J. Dietary squalene increases high density lipoprotein-cholesterol and paraoxonase 1 and decreases oxidative stress in mice. PLoS ONE 2015, 9, e104224. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Beamonte, R.; Navarro, M.A.; Acin, S.; Guillen, N.; Barranquero, C.; Arnal, C.; Surra, J.; Osada, J. Postprandial changes in high density lipoproteins in rats subjected to gavage administration of virgin olive oil. PLoS ONE 2013, 8, e55231. [Google Scholar] [CrossRef]
- Navab, M.; Hama, S.Y.; Hough, G.P.; Subbanagounder, G.; Reddy, S.T.; Fogelman, A.M. A cell-free assay for detecting HDL that is dysfunctional in preventing the formation of or inactivating oxidized phospholipids. J. Lipid Res. 2001, 42, 1308–1317. [Google Scholar] [CrossRef]
- Arbones-Mainar, J.M.; Navarro, M.A.; Guzman, M.A.; Arnal, C.; Surra, J.C.; Acin, S.; Carnicer, R.; Osada, J.; Roche, H.M. Selective effect of conjugated linoleic acid isomers on atherosclerotic lesion development in apolipoprotein E knockout mice. Atherosclerosis 2006, 189, 318–327. [Google Scholar] [CrossRef]
- Guevara, N.V.; Kim, H.S.; Antonova, E.I.; Chan, L. The absence of p53 accelerates atherosclerosis by increasing cell proliferation in vivo. Nat. Med. 1999, 5, 335–339. [Google Scholar] [CrossRef]
- Guillen, N.; Acin, S.; Navarro, M.A.; Perona, J.S.; Arbones-Mainar, J.M.; Arnal, C.; Sarria, A.J.; Surra, J.C.; Carnicer, R.; Orman, I.; et al. Squalene in a sex-dependent manner modulates atherosclerotic lesion which correlates with hepatic fat content in apoE-knockout male mice. Atherosclerosis 2008, 197, 72–83. [Google Scholar] [CrossRef]
- Paigen, B.; Morrow, A.; Holmes, P.A.; Mitchell, D.; Williams, R.A. Quantitative assessment of atherosclerotic lesions in mice. Atherosclerosis 1987, 68, 231–240. [Google Scholar] [CrossRef]
- John, A.M.; Bell, J.M. Amino-Acid Requirements of Growing Mouse. J. Nutr. 1976, 106, 1361–1367. [Google Scholar] [CrossRef]
- White, P.J.; Lapworth, A.L.; McGarrah, R.W.; Kwee, L.C.; Crown, S.B.; Ilkayeva, O.; An, J.; Carson, M.W.; Christopher, B.A.; Ball, J.R.; et al. Muscle-Liver Trafficking of BCAA-Derived Nitrogen Underlies Obesity-Related Glycine Depletion. Cell Rep. 2020, 33, 108375. [Google Scholar] [CrossRef]
- Dhot, J.; Prat, V.; Ferron, M.; Aillerie, V.; Erraud, A.; Rozec, B.; Waard, M.D.; Gauthier, C.; Lauzier, B. Implications of a Soy-Based Diet for Animal Models. Int. J. Mol. Sci. 2021, 22, 774. [Google Scholar] [CrossRef]
- Tall, A.R. Plasma cholesteryl ester transfer protein. J. Lipid Res. 1993, 34, 1255–1274. [Google Scholar] [CrossRef]
- Peng, H.; Chiu, T.-Y.; Liang, Y.-J.; Lee, C.-J.; Liu, C.-S.; Suen, C.-S.; Yen, J.J.-Y.; Chen, H.-T.; Hwang, M.-J.; Hussain, M.M.; et al. PRAP1 is a novel lipid binding protein that promotes lipid absorption by facilitating MTTP-mediated lipid transport. J. Biol. Chem. 2020, 296, 100052. [Google Scholar] [CrossRef]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef]
- Acin, S.; Navarro, M.A.; Carnicer, R.; Arbones-Mainar, J.M.; Guzman, M.A.; Arnal, C.; Beltran, G.; Uceda, M.; Maeda, N.; Osada, J. Dietary cholesterol suppresses the ability of olive oil to delay the development of atherosclerotic lesions in apolipoprotein E knockout mice. Atherosclerosis 2005, 182, 17–28. [Google Scholar] [CrossRef]
- Libby, P. The changing landscape of atherosclerosis. Nature 2021, 592, 524–533. [Google Scholar] [CrossRef]
- Yahia, S.; Benomar, S.; Dehiba, F.; Allaoui, A.; Guillen, N.; Rodriguez-Yoldi Maria, J.; Osada, J.; Boualga, A. Hypocholesterolaemic and antioxidant efficiency of chickpea (Cicer arietinum) protein hydrolysates depend on its degree of hydrolysis in cholesterol-fed rat. Nutr. Food Sci. 2017, 47, 254–269. [Google Scholar] [CrossRef]
- Allaoui, A.; Barranquero, C.; Yahia, S.; Herrera-Marcos Luis, V.; Benomar, S.; Jridi, M.; Navarro María, Á.; Rodriguez-Yoldi Maria, J.; Nasri, M.; Osada, J.; et al. Fenugreek proteins and their hydrolysates prevent hypercholesterolemia and enhance the HDL antioxidant properties in rats. Nutr. Food Sci. 2018, 48, 973–989. [Google Scholar] [CrossRef]
- Halim, H.; Pinkaew, D.; Chunhacha, P.; Sinthujaroen, P.; Thiagarajan, P.; Fujise, K. Ticagrelor induces paraoxonase-1 (PON1) and better protects hypercholesterolemic mice against atherosclerosis compared to clopidogrel. PLoS ONE 2019, 14, e0218934. [Google Scholar] [CrossRef]
- Lopez, P.; Rodriguez-Carrio, J.; Martinez-Zapico, A.; Perez-Alvarez, A.I.; Lopez-Mejias, R.; Benavente, L.; Mozo, L.; Caminal-Montero, L.; Gonzalez-Gay, M.A.; Suarez, A. Serum Levels of Anti-PON1 and Anti-HDL Antibodies as Potential Biomarkers of Premature Atherosclerosis in Systemic Lupus Erythematosus. Thromb. Haemost. 2017, 117, 2194–2206. [Google Scholar] [CrossRef] [PubMed]
- Lou-Bonafonte, J.M.; Gabas-Rivera, C.; Navarro, M.A.; Osada, J. PON1 and Mediterranean Diet. Nutrients 2015, 7, 4068–4092. [Google Scholar] [CrossRef] [PubMed]
- Kratzer, A.; Giral, H.; Landmesser, U. High-density lipoproteins as modulators of endothelial cell functions: Alterations in patients with coronary artery disease. Cardiovasc. Res. 2014, 103, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Hegele, R.A.; Boren, J.; Ginsberg, H.N.; Arca, M.; Averna, M.; Binder, C.J.; Calabresi, L.; Chapman, M.J.; Cuchel, M.; von Eckardstein, A.; et al. Rare dyslipidaemias, from phenotype to genotype to management: A European Atherosclerosis Society task force consensus statement. Lancet Diabetes Endocrinol. 2020, 8, 50–67. [Google Scholar] [CrossRef]
- Arbones-Mainar, J.M.; Navarro, M.A.; Carnicer, R.; Guillen, N.; Surra, J.C.; Acin, S.; Guzman, M.A.; Sarria, A.J.; Arnal, C.; Aguilera, M.P.; et al. Accelerated atherosclerosis in apolipoprotein E-deficient mice fed Western diets containing palm oil compared with extra virgin olive oils: A role for small, dense high-density lipoproteins. Atherosclerosis 2007, 194, 372–382. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Soybean (%, w/w) | Chicken (%, w/w) | Turkey (%, w/w) |
|---|---|---|---|
| Corn starch a | 37.2 | 37.2 | 37.2 |
| Soybean protein | 11.2 | ||
| Chicken protein | 11.2 | ||
| Turkey protein | 11.2 | ||
| Maltodextrin a | 12.4 | 12.4 | 12.4 |
| Sucrose b | 8.0 | 8.0 | 8.0 |
| Soybean oil c | 3.2 | 3.2 | 3.2 |
| Cellulose d | 4.0 | 4.0 | 4.0 |
| Mineral mix e | 2.8 | 2.8 | 2.8 |
| Vitamin mix f | 0.8 | 0.8 | 0.8 |
| Choline g | 0.2 | 0.2 | 0.2 |
| L-Cystine g | 0.1 | 0.1 | 0.1 |
| Cholesterol g | 0.15 | 0.15 | 0.15 |
| Palm oil h | 20 | 20 | 20 |
| Soybean | Chicken | Turkey | |
|---|---|---|---|
| Protein (%, w/w dry) | 10.9 ± 0.2 | 10.6 ± 0.1 | 10.7 ± 0.1 |
| Amino acids (%, w/w) * | |||
| Non-essential | |||
| Alanine (Ala) | 0.40 | 0.61 | 0.62 |
| Arginine (Arg) | 0.64 | 0.59 | 0.61 |
| Aspartic acid (Asp) | 1.00 | 0.93 | 1.00 |
| Cysteine (Cys) | 0.16 | 0.14 | 0.14 |
| Glutamic acid (Glu) | 1.66 | 1.46 | 1.47 |
| Glycine (Gly) | 0.36 | 0.44 | 0.43 |
| Proline (Pro) | 0.46 | 0.38 | 0.36 |
| Serine (Ser) | 0.46 | 0.41 | 0.39 |
| Tyrosine (Tyr) | 0.29 | 0.27 | 0.30 |
| Total non-essential | 5.43 | 5.23 | 5.32 |
| Essential | |||
| Histidine (His) | 0.20 | 0.28 | 0.29 |
| Isoleucine (Ileu) | 0.39 | 0.49 | 0.48 |
| Leucine (Leu) | 0.66 | 0.81 | 0.79 |
| Lysine (Lys) | 0.58 | 0.89 | 0.90 |
| Methionine (Met) | 0.10 | 0.22 | 0.22 |
| Phenylalanine (Phe) | 0.44 | 0.32 | 0.31 |
| Threonine (Thr) | 0.34 | 0.44 | 0.44 |
| Tryptophan (Trp) | 0.15 | 0.13 | 0.11 |
| Valine (Val) | 0.45 | 0.55 | 0.55 |
| Total essential | 3.31 | 4.13 | 4.09 |
| Essential/Non-essential | 0.6 | 0.8 | 0.8 |
| Soybean Group | Chicken Group | Turkey Group | |
|---|---|---|---|
| Males (n) | 14 | 15 | 16 |
| Triglycerides (mg/dL) * | 150 ± 35 | 88 ± 17 a | 115 ± 44 a,b |
| Total cholesterol (mg/dL) | 808 ± 128 | 765 ± 166 | 670 ± 230 |
| Glucose (mg/dL) | 277 ± 41 | 220 ± 44 | 342 ± 182 |
| Non-esterified fatty acids (mg/dL) | 37 ± 6 | 44 ± 8 | 38 ± 13 |
| Apolipoprotein A1 (arbitrary units) | 4.1 ± 0.6 | 3.9 ± 0.5 | 4.2 ± 0.6 |
| Paraoxonase 1 (IU/L) * | 43,249 ± 5704 | 62,541 ± 10,589 a | 74,940 ± 17,834 a |
| Females (n) | 10 | 10 | 10 |
| Triglycerides (mg/dL) | 168 ± 35 | 177 ± 44 | 212 ± 71 |
| Total cholesterol (mg/dL) | 510 ± 163 | 553 ± 93 | 502 ± 58 |
| Glucose (mg/dL) | 270 ± 71 | 320 ± 95 | 341 ± 79 |
| Non-esterified fatty acids (mg/dL) | 43 ± 8 | 44 ± 7 | 44 ± 10 |
| Apolipoprotein A1 (arbitrary units) | 5.0 ± 0.6 | 4.6 ± 0.4 | 4.7 ± 0.5 |
| Paraoxonase 1 (IU/L) | 70,076 ± 10,891 | 63,461 ± 6493 | 74,854 ± 11,778 b |
| Soybean Group | Chicken Group | Turkey Group | |
|---|---|---|---|
| Males (n) | 14 | 15 | 16 |
| Total cholesterol (mg/dL) | |||
| VLDL | 510 ± 92 | 447 ± 129 | 465 ± 200 |
| LDL | 272 ± 49 | 239 ± 70 | 248 ± 107 |
| HDL | 48 ± 9 | 42 ± 12 | 44 ± 14 |
| Esterified cholesterol (mg/dL) | |||
| VLDL | 121 ± 22 | 106 ± 31 | 79 ± 34 a,b |
| LDL | 96 ± 17 | 84 ± 24 | 82 ± 35 |
| HDL | 34 ± 6 | 30 ± 8 | 33 ± 14 |
| Non-esterified cholesterol (mg/dL) | |||
| VLDL | 396 ± 71 | 347 ± 100 | 384 ± 161 |
| LDL | 166 ± 30 | 146 ± 42 | 161 ± 69 |
| HDL | 16 ± 3 | 14 ± 4 | 15 ± 7 |
| Females (n) | 10 | 10 | 10 |
| Total cholesterol (mg/dL) | |||
| VLDL | 337 ± 58 | 352 ± 59 | 279 ± 55 a,b |
| LDL | 179 ± 31 | 175 ± 29 | 180 ± 35 |
| HDL | 15 ± 3 | 25 ± 4 a | 18 ± 4 b |
| Esterified cholesterol (mg/dL) | |||
| VLDL | 224 ± 39 | 92 ± 16 a | 109 ± 21 a |
| LDL | 144 ± 25 | 80 ± 13 a | 92 ± 18 a |
| HDL | 8 ± 1 | 13 ± 2 a | 9 ± 2 b |
| Non-esterified cholesterol (mg/dL) | |||
| VLDL | 112 ± 19 | 261 ± 44 a | 191 ± 37 a,b |
| LDL | 36 ± 6 | 94 ± 16 a | 69 ± 14 a,b |
| HDL | 7 ± 1 | 10 ± 2 a | 7 ± 1 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Beamonte, R.; Sánchez-Marco, J.; Lázaro, G.; Barco, M.; Herrero-Continente, T.; Serrano-Megías, M.; Botaya, D.; Arnal, C.; Barranquero, C.; Surra, J.C.; et al. Dietary Avian Proteins Are Comparable to Soybean Proteins on the Atherosclerosis Development and Fatty Liver Disease in Apoe-Deficient Mice. Nutrients 2021, 13, 1838. https://doi.org/10.3390/nu13061838
Martínez-Beamonte R, Sánchez-Marco J, Lázaro G, Barco M, Herrero-Continente T, Serrano-Megías M, Botaya D, Arnal C, Barranquero C, Surra JC, et al. Dietary Avian Proteins Are Comparable to Soybean Proteins on the Atherosclerosis Development and Fatty Liver Disease in Apoe-Deficient Mice. Nutrients. 2021; 13(6):1838. https://doi.org/10.3390/nu13061838
Chicago/Turabian StyleMartínez-Beamonte, Roberto, Javier Sánchez-Marco, Gonzalo Lázaro, María Barco, Tania Herrero-Continente, Marta Serrano-Megías, David Botaya, Carmen Arnal, Cristina Barranquero, Joaquín C. Surra, and et al. 2021. "Dietary Avian Proteins Are Comparable to Soybean Proteins on the Atherosclerosis Development and Fatty Liver Disease in Apoe-Deficient Mice" Nutrients 13, no. 6: 1838. https://doi.org/10.3390/nu13061838
APA StyleMartínez-Beamonte, R., Sánchez-Marco, J., Lázaro, G., Barco, M., Herrero-Continente, T., Serrano-Megías, M., Botaya, D., Arnal, C., Barranquero, C., Surra, J. C., Osada, J., & Navarro, M. A. (2021). Dietary Avian Proteins Are Comparable to Soybean Proteins on the Atherosclerosis Development and Fatty Liver Disease in Apoe-Deficient Mice. Nutrients, 13(6), 1838. https://doi.org/10.3390/nu13061838