
A Low-Carbohydrate Ketogenic Diet and Treadmill Training Enhanced Fatty Acid Oxidation Capacity but Did Not Enhance Maximal Exercise Capacity in Mice




Abstract
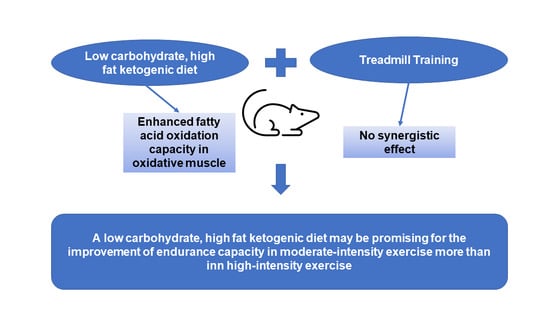
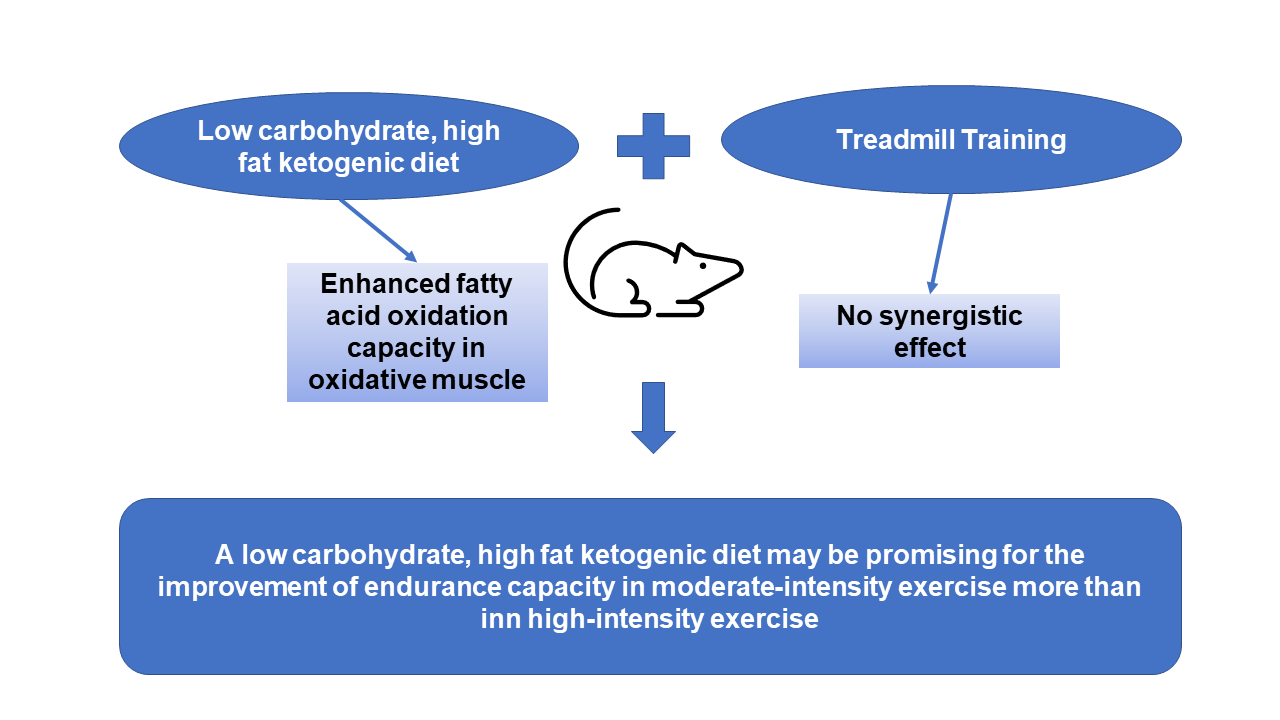
1. Introduction
2. Materials and Methods
2.1. Mice and Diets
2.2. Training Procedures, Forelimb Grip Strength Test, and Maximal Workload
2.3. Sampling and Plasma Biochemical Assessment
2.4. Real-Time PCR
2.5. Plasma Fibroblast Growth Factor 21 (FGF21) and Testosterone Assessment
2.6. Muscle Glycogen Assessment
2.7. Statistical Analysis
3. Results
3.1. Weight Change and Relative Tissue or Organ Weight
3.2. Grip Power and Maximal Exercise Capacity Test
3.3. Analysis of Plasma Biochemical Indexes
3.4. Plasma FGF21 and Testestorone Concentrations
3.5. Muscle Glygocen Content
3.6. Gene Expression in Tissues and Organs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Poti, J.M.; Braga, B.; Qin, B. Ultra-processed Food Intake and Obesity: What Really Matters for Health—Processing or Nutrient Content? Curr. Obes. Rep. 2017, 6, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Almoosawi, S.; Vingeliene, S.; Gachon, F.; Voortman, T.; Palla, L.; Johnston, J.D.; Van Dam, R.M.; Darimont, C.; Karagounis, L.G. Chronotype: Implications for Epidemiologic Studies on Chrono-Nutrition and Cardiometabolic Health. Adv. Nutr. 2019, 10, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Harvie, M.; Howell, A. Potential Benefits and Harms of Intermittent Energy Restriction and Intermittent Fasting Amongst Obese, Overweight and Normal Weight Subjects—A Narrative Review of Human and Animal Evidence. Behav. Sci. 2017, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, S.; Shibata, S. The role of circadian rhythms in muscular and osseous physiology and their regulation by nutrition and exercise. Front. Neurosci. 2017, 11, 63. [Google Scholar] [CrossRef]
- Bialostosky, K.; Wright, J.D.; Kennedy-Stephenson, J.; McDowell, M.; Johnson, C.L. Dietary intake of macronutrients, micronutrients, and other dietary constituents: United States, 1988–94: Data from the National Health Examination Survey, the National Health and Nutrition Examination Surveys, and the Hispanic Health and Nutrition Examination Survey. Vital Health Stat. 2002, 11, 245. [Google Scholar]
- Kossoff, E.H.; Dorward, J.L. The modified Atkins diet. Epilepsia 2008, 49, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Zopf, Y.; Reljic, D.; Dieterich, W. Dietary Effects on Microbiota—New Trends with Gluten-Free or Paleo Diet. Med. Sci. 2018, 6, 92. [Google Scholar] [CrossRef]
- McGrice, M.; Porter, J. The effect of low carbohydrate diets on fertility hormones and outcomes in overweight and obese women: A systematic review. Nutrients 2017, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Suzuki, K. Keto-Adaptation and Endurance Exercise Capacity, Fatigue Recovery, and Exercise-Induced Muscle and Organ Damage Prevention: A Narrative Review. Sports 2019, 7, 40. [Google Scholar] [CrossRef]
- Erickson, N.; Boscheri, A.; Linke, B.; Huebner, J.J.M.O. Systematic review: Isocaloric ketogenic dietary regimes for cancer patients. Med. Oncol. 2017, 5, 72. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Suzuki, K. Potential Application of Ketogenic Diet to Metabolic Status and Exercise Performance: A Review. EC Nutr. 2018, 13, 496–499. [Google Scholar]
- Leal, L.G.; Lopes, M.A.; Batista, M.L., Jr. Physical exercise-induced myokines and muscle-adipose tissue crosstalk: A review of current knowledge and the implications for health and metabolic diseases. Front. Physiol. 2018, 9, 1307. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adiponectin and inflammation: Consensus and controversy. J. Allergy Clin. Immunol. 2008, 2, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R.; Pissios, P.; Otu, H.; Xue, B.; Asakura, K.; Furukawa, N.; Marino, F.E.; Liu, F.F.; Kahn, B.B.; Libermann, T.A.; et al. A high-fat, ketogenic diet induces a unique metabolic state in mice. Am. J. Physiol. Endocrinol. Metabol. 2007, 6, E1724–E1739. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; Adams, A.C. Inventing new medicines: The FGF21 story. Mol. Metabol. 2014, 3, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Urban, R.J.; Bodenburg, Y.H.; Gilkison, C.; Foxworth, J.; Coggan, A.R.; Wolfe, R.R.; Ferrando, A. Testosterone administration to elderly men increases skeletal muscle strength and protein synthesis. Am. J. Physiol. Endocrinol. 1995, 5, E820–E826. [Google Scholar] [CrossRef] [PubMed]
- Basciani, S.; Camajani, E.; Contini, S.; Persichetti, A.; Risi, R.; Bertoldi, L.; Strigari, L.; Prossomarti, G.; Watanabe, M.; Mariani, S.; et al. Very-low-calorie ketogenic diets with whey, vegetable, or animal protein in patients with obesity: A randomized pilot study. J. Clin. Endocrinol. Metabol. 2020, 9, 2939–2949. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Huang, Q.; Yada, K.; Liu, C.; Suzuki, K. An 8-Week Ketogenic Low Carbohydrate, High Fat Diet Enhanced Exhaustive Exercise Capacity in Mice. Nutrients 2018, 10, 673. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Huang, Q.; Tominaga, T.; Liu, C.; Suzuki, K. An 8-Week Ketogenic Diet Alternated Interleukin-6, Ketolytic and Lipolytic Gene Expression, and Enhanced Exercise Capacity in Mice. Nutrients 2018, 10, 1696. [Google Scholar] [CrossRef] [PubMed]
- Avila, J.J.; Kim, S.K.; Massett, M.P. Differences in Exercise Capacity and Responses to Training in 24 Inbred Mouse Strains. Front. Physiol. 2017, 8, 974. [Google Scholar] [CrossRef] [PubMed]
- Massett, M.P.; Fan, R.; Berk, B.C. Quantitative trait loci for exercise training responses in FVB/NJ and C57BL/6J mice. Physiol. Genom. 2009, 1, 15–22. [Google Scholar] [CrossRef]
- Nymo, S.; Coutinho, S.R.; Jørgensen, J.; Rehfeld, J.F.; Truby, H.; Kulseng, B.; Martins, C. Timeline of changes in appetite during weight loss with a ketogenic diet. Int. J. Obes. 2017, 8, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.I.; Appel, S. Energy expenditure and body composition changes after an isocaloric ketogenic diet in overweight and obese men: A secondary analysis of energy expenditure and physical activity. PLoS ONE 2019, 14, e0222971. [Google Scholar] [CrossRef] [PubMed]
- Bueno, N.B.; de Melo, I.S.; de Oliveira, S.L.; da Rocha Ataide, T. Verylow-carbohydrate ketogenic diet v. low-fat diet for long-term weight loss: A meta-analysis of randomised controlled trials. Br. J. Nutr. 2013, 110, 1178–1187. [Google Scholar] [CrossRef]
- Naude, C.E.; Schoonees, A.; Senekal, M.; Young, T.; Garner, P.; Volmink, J. Low carbohydrate versus isoenergetic balanced diets for reducing weight and cardiovascular risk: A systematic review and meta-analysis. PLoS ONE 2014, 7, e100652. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, J. Interest in the Ketogenic Diet Grows for Weight Loss and Type 2 Diabetes. JAMA 2018, 319, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Westman, E.C.; Yancy, W.S., Jr. Using a low-carbohydrate diet to treat obesity and type 2 diabetes mellitus. Curr. Opin. Endocrinol. Diabetes Obes. 2020, 5, 255–260. [Google Scholar] [CrossRef]
- Meng, Y.; Bai, H.; Wang, S.; Li, Z.; Wang, Q.; Chen, L. Efficacy of low carbohydrate diet for type 2 diabetes mellitus management: A systematic review and meta-analysis of randomized controlled trials. Diabetes Res. Clin. Pract. 2017, 131, 124–131. [Google Scholar] [CrossRef]
- Paoli, A. Ketogenic Diet for Obesity: Friend or Foe? Int. J. Environ. Res. Public Health 2014, 11, 2092–2107. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metabol. 2020, 2, 817–828. [Google Scholar] [CrossRef]
- Paoli, A.; Grimaldi, K.; D’Agostino, D.; Cenci, L.; Moro, T.; Bianco, A.; Palma, A. Ketogenic diet does not affect strength performance in elite artistic gymnasts. J. Int. Soc. Sports Nutr. 2012, 1, 34. [Google Scholar] [CrossRef]
- Barnhart, E. The Effect of Nutritional Ketosis on Strength and Power in Tactical Athletes. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2018. [Google Scholar]
- Zajac, A.; Poprzecki, S.; Maszczyk, A.; Czuba, M.; Michalczyk, M.; Zydek, G. The effects of a ketogenic diet on exercise metabolism and physical performance in off-road cyclists. Nutrients 2014, 6, 2493–2508. [Google Scholar] [CrossRef] [PubMed]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. Endocrinol. Metabol. 1993, 3, E380–E391. [Google Scholar] [CrossRef]
- Garbow, J.R.; Doherty, J.M.; Schugar, R.C.; Travers, S.; Weber, M.L.; Wentz, A.E.; Ezenwajiaku, N.; Cotter, D.G.; Brunt, E.M.; Crawford, P.A. Hepatic steatosis, inflammation, and ER stress in mice maintained long term on a very low-carbohydrate ketogenic diet. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G956–G967. [Google Scholar] [CrossRef]
- Milder, J.B.; Liang, L.-P.; Patel, M. Acute oxidative stress and systemic Nrf2 activation by the ketogenic diet. Neurobiol. Dis. 2010, 40, 238–244. [Google Scholar] [CrossRef]
- Schugar, R.C.; Crawford, P.A. Low-carbohydrate ketogenic diets, glucose homeostasis, and nonalcoholic fatty liver disease. Curr. Opin. Clin. Nutr. Metab. Care 2020, 4, 374. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Ma, S.; Tominaga, T.; Suzuki, K.; Liu, C. An 8-Week, Low carbohydrate, high fat, ketogenic diet enhanced exhaustive exercise capacity in mice Part 2: Effect on fatigue recovery, post-exercise biomarkers and anti-oxidation capacity. Nutrients 2018, 10, 1339. [Google Scholar] [CrossRef] [PubMed]
- Fisher, F.M.; Maratos-Flier, E. Understanding the physiology of FGF21. Annu. Rev. Physiol. 2016, 78, 223–241. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, S.H.; Min, Y.K.; Yang, H.M.; Lee, J.B.; Lee, M.S. Acute exercise induces FGF21 expression in mice and in healthy humans. PLoS ONE 2013, 8, e63517. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.I.; Goodyear, L.J. Muscle-Adipose Tissue Cross Talk. Cold Spring Harb. Perspect. Med. 2018, 8, a029801. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Palmieri, A.; Mazzarella, G.; Costagliola, C.; Bianco, A.; Daniele, A. New Insight into Adiponectin Role in Obesity and Obesity-Related Diseases. BioMed Res. Int. 2014, 2014, 658913. [Google Scholar] [CrossRef]
- Ruskin, D.N.; Kawamura, M.; Masino, S.A. Reduced Pain and Inflammation in Juvenile and Adult Rats Fed a Ketogenic Diet. PLoS ONE 2009, 4, e8349. [Google Scholar] [CrossRef]
- Diop, S.B.; Bisharat-Kernizan, J.; Birse, R.T.; Oldham, S.; Ocorr, K.; Bodmer, R. PGC-1/Spargel Counteracts High-Fat-Diet-Induced Obesity and Cardiac Lipotoxicity Downstream of TOR and Brummer ATGL Lipase. Cell Rep. 2015, 10, 1572–1584. [Google Scholar] [CrossRef]
- Kawanishi, N.; Mizokami, T.; Yano, H.; Suzuki, K. Exercise attenuates M1 macrophages and CD8+ T cells in the adipose tissue of obese mice. Med. Sci. Sports Exerc. 2013, 45, 1684–1693. [Google Scholar] [CrossRef]
- Razzoli, M.; Emmett, M.J.; Lazar, M.A.; Bartolomucci, A. β-Adrenergic receptors control brown adipose UCP-1 tone and cold response without affecting its circadian rhythmicity. FASEB J. 2018, 32, 5640–5646. [Google Scholar] [CrossRef] [PubMed]
- Nakao, R.; Shimba, S.; Oishi, K. Ketogenic diet induces expression of the muscle circadian gene Slc25a25 via neural pathway that might be involved in muscle thermogenesis. Sci. Rep. 2017, 7, 2885. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.Y.; Goldsmith, F.R.; Fuller, S.E.; Simon, J.; Batdorf, H.M.; Scott, M.C.; Essajee, N.M.; Brown, J.M.; Burk, D.H.; Morrison, C.D.; et al. Response of Liver Metabolic Pathways to Ketogenic Diet and Exercise Are Not Additive. Med. Sci. Sports Exerc. 2020, 52, 37–48. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Con | LCKD | |
|---|---|---|
| Protein (%) | 14.2 | 4.8 |
| Fat (%) | 10.1 | 93.2 |
| Carbohydrate (%) | 75.7 | 1.9 |
| Kcal/g | 3.6 | 7.3 |
| Forward | Reverse | |
|---|---|---|
| 18s | CGGCTACCACATCCAAGGA | AGCTGGAATTACCGCGGC |
| Adiponectin | AGAGTCGTTGACGTTATCTGCATA | GGGCTCTGTGCTGCTCCATCT |
| Atgl | GAGCCCCGGGGTGGAACAAGAT | AAAAGGTGGTGGGCAGGAGTAAGG |
| Cd36 | TGGCCTTACTTGGGATTGG | CCAGTGTATATGTAGGCTCATCCA |
| F480 | CTTTGGCTATGGGCTTCCAGTC | GCAAGGAGGACAGAGTTTATCGTG |
| Il-6 | AACGATGATGCACTTGCAGA | TGGTACTCCAGAAGACCAGAGG |
| Klotho | TGTTCTGCTGCGAGCTGTTAC | TACCGGACTCACGTACTGTTT |
| Leptin | GCTTTGGTCCTATCTGTCTTATGTT | CAATGGTCTTGATGAGGGTTTT |
| Prdm16 | CCACCAGCGAGGACTTCA | GGAGGACTCTCGTAGCTCGAA |
| Pgc1α | GACTGGAGGAAGACTAAACGGCCA | GCCAGTCACAGGAGGCATCTTT |
| Pparγ | CCACCAGCGAGGACTTCAC | GGAGGACTCTCGTAGCTCGAA |
| Cidea | TCCTCGGCTGTCTCAATG | GGCTGCTCTTCTGTATCG |
| Ucp-1 | TGGTTGGTTTTATTCGTGGT | AGGGTTTGTGGCTTCTTTTC |
| Cox4 | TGGGAGTGTTGTGAAGAGTGA | GCAGTGAAGCCGATGAAGAAC |
| Cs | GCAGCCAAGAACTCATCCTG | TCTGGGCCTGCTCCTTAGGTA |
| Cytochrome c | CACGCTTTACCCTTCGTTCT | CTCATTTCCCTGCCATTCTC |
| Gp | TGGCAGAAGTGGTGAACAATGAC | CCGTGGAGATCTGCTCCGATA |
| Gs | ACTGCTTGGGCGTTATCTCTGTG | ATGCCCGCTCCATGCGTA |
| Hadh | ACTACATCAAAATGGGCTCTCAG | AGCAGAAATGGAATGCGGACC |
| Hbdh | AGTTTGGGGTCGAGGCTTTC | TGGTGGCCGCTATGAAGTTG |
| Hk2 | CTGTCTACAAGAAACATCCCCATTT | CACCGCCGTCACCATAGC |
| Mct1 | GCCTGAGCAAGTCAAGCTAG | TCAGACCTCGGATCCAGTAC |
| Oxct1 | CCAAGGAAGTAAATGAAGATCTCCTA | ACGTGTATGTTACAAGAAATGGCTTACC |
| Sirt1 | GCAACAGCATCTTGCCTGAT | GTGCTACTGGTCTCACTT |
| Tfam | TTCCCAAGACTTCATTTCATTGTC | GATGATTCGGCTCAGGGAAA |
| Acat1 | CCGAGACAACTACCCAAGGA | CACACACAGGACCAGGACAC |
| Cpt1α | CCAGGCTACAGTGGGACATT | GAACTTGCCCATGTCCTTGT |
| Mcad | GCTCGTGAGCACATTGAAAA | CATTGTCCAAAAGCCAAACC |
| Con | Con + T | LCKD | LCKD + T | Significance | |||
|---|---|---|---|---|---|---|---|
| Training | Feed | Interaction | |||||
| MW * | 19,641 ± 1345 bc | 41,378 ± 1269 ad | 11,680 ± 512 ad | 27,385 ± 3282 bd | *** | *** | *** |
| Con | Con + T | LCKD | LCKD + T | Significance | |||
|---|---|---|---|---|---|---|---|
| Training | Feed | Interaction | |||||
| β-hydroxybutyrate, mmol/L | 0.40 ± 0.42 | 0.31 ± 0.35 | 4.48 ± 1.91 | 5.00 ± 3.22 | n. s. | *** | n. s. |
| Albumin, mg/dL | 2.55 ± 0.23 | 2.43 ± 0.19 | 2.33 ± 0.65 | 2.21 ± 0.27 | n. s. | n. s. | n. s. |
| BUN, mg/dL | 23.44 ± 5.02 | 25.08 ± 3.50 | 16.08 ± 4.40 | 15.45 ± 1.81 | n. s. | *** | n. s. |
| Creatinine, mg/dL | 0.10 ± 0.03 c | 0.12 ± 0.02 | 0.07 ± 0.03 ad | 0.12 ± 0.02 c | *** | * | ** |
| Glucose, mg/dL | 201.4 ± 58.67 | 212.3 ± 33.78 | 135.8 ± 31.74 | 123.8 ± 33.17 | n. s. | *** | n. s. |
| T-CHO, mg/dL | 100.9 ± 10.13 | 93.38 ± 8.40 | 135.8 ± 31.74 | 144.0 ± 31.34 | n. s. | *** | n. s. |
| L-CHO, mg/dL | 13.88 ± 3.56 | 15.00 ± 4.81 | 23.63 ± 4.66 | 24.00 ± 7.52 | n. s. | *** | n. s. |
| H-CHO, mg/dL | 76.13 ± 8.17 | 69.75 ± 6.94 | 107.3 ± 28.41 | 111.0 ± 25.25 | n. s. | *** | n. s. |
| NEFA, μEq/L | 2.85 ± 0.31 | 2.64 ± 0.19 | 2.68 ± 0.52 d | 3.11 ± 0.37 c | n. s. | n. s. | * |
| TG, mg/dL | 20.63 ± 7.25 | 16.13 ± 7.85 | 21.75 ± 11.08 | 14.14 ± 4.81 | * | n. s. | n. s. |
| UA, mg/dL | 1.39 ± 0.51 | 1.39 ± 0.51 | 1.35 ± 0.89 | 1.35 ± 0.89 | n. s. | n. s. | n. s. |
| Con | Con + T | LCKD | LCKD + T | Significance | |||
|---|---|---|---|---|---|---|---|
| Training | Feed | Interaction | |||||
| Amylase, IL/L | 1755 ± 252 | 1623 ± 220 | 1525 ± 341 | 1664 ± 155 | n. s. | n. s. | n. s. |
| AST, IU/L | 81 ± 47 | 69 ± 48 | 151 ± 86 | 152 ± 92 | n. s. | ** | n. s. |
| CK, IU/L | 155 ± 130 | 136 ± 205 | 192 ± 121 | 228 ± 130 | n. s. | n. s. | n. s. |
| LDH, IU/L | 421 ± 290 | 881 ± 299 | 536 ± 463 | 548 ± 224 | n. s. | n. s. | n. s. |
| Lipase, IU/L | 40 ± 7 | 46 ± 6 | 66 ± 15 | 74 ± 13 | n. s. | *** | n. s. |
| Con | Con + T | LCKD | LCKD + T | Significance | |||
|---|---|---|---|---|---|---|---|
| Training | Feed | Interaction | |||||
| Muscle glycogen, μg/g protein | 26.3 ± 9 | 35.2 ± 2.5 | 27.3 ± 1.17 | 32.4 ± 0.12 | n. s. | n. s. | n. s. |
| Con | Con + T | LCKD | LCKD + T | Significance | |||
|---|---|---|---|---|---|---|---|
| Training | Feed | Interaction | |||||
| Adiponectin (Anti-inflammation) | 1.00 ± 0.46 | 1.01 ± 0.29 | 0.44 ± 0.24 | 0.52 ± 0.14 | n. s. | *** | n. s. |
| Atgl (Fatty acid mobilization) | 1.00 ± 0.40 | 0.95 ± 0.43 | 0.30 ± 0.12 | 0.30 ± 0.14 | n. s. | * | n. s. |
| Cd36 (Fatty acid transportation) | 1.00 ± 0.36 | 0.90 ± 0.29 | 0.51 ± 0.37 | 0.45 ± 0.32 | n. s. | n. s. | n. s. |
| F4/80 (Inflammation) | 1.00 ± 0.30 | 0.91 ± 0.20 | 0.50 ± 0.24 | 0.55 ± 0.25 | n. s. | *** | n. s. |
| Il-6 (Inflammation) | 1.00 ± 0.51 | 0.53 ± 0.19 | 0.26 ± 0.14 | 0.21 ± 0.61 | ** | *** | n. s. |
| Klotho (FGF21 receptor) | 1.00 ± 0.34 | 1.05 ± 0.50 | 0.95 ± 0.64 | 0.91 ± 0.55 | n. s. | n. s. | n. s. |
| Leptin (Energy metabolism) | 1.00 ± 0.43 bc | 0.49 ± 0.34 a | 0.43 ± 0.39 a | 0.50 ± 0.23 | n. s. | * | * |
| Prdm16 (Browning of WAT) | 1.00 ± 0.29 | 0.88 ± 0.22 | 1.07 ± 1.23 | 0.75 ± 0.61 | n. s. | n. s. | n. s. |
| Pgc1α (Browning of WAT) | 1.00 ± 0.67 | 1.23 ± 0.68 | 0.63 ± 0.32 | 0.63 ± 0.36 | n. s. | * | n. s. |
| Pparγ (Browning of WAT) | 1.00 ± 0.51 | 1.23 ± 0.54 | 0.69 ± 0.44 | 0.90 ± 0.63 | n. s. | n. s. | n. s. |
| Con | Con + T | LCKD | LCKD + T | Significance | |||
|---|---|---|---|---|---|---|---|
| Training | Feed | Interaction | |||||
| Cidea (Thermogenesis) | 1.00 ± 0.15 | 1.20 ± 0.07 | 1.08 ± 0.89 | 0.81 ± 0.12 | n. s. | n. s. | n. s. |
| Il-6 (Inflammation) | 1.00 ± 0.51 | 1.45 ± 0.55 | 1.89 ± 2.65 | 1.32 ± 0.42 | n. s. | n. s. | n. s. |
| Prdm16 (Thermogenesis) | 1.00 ± 0.28 | 1.15 ± 0.34 | 0.83 ± 0.61 | 0.59 ± 0.08 | n. s. | n. s. | n. s. |
| Ucp1 (Thermogenesis) | 1.00 ± 0.24 | 1.60 ± 0.54 | 2.42 ± 1.82 | 1.85 ± 0.43 | n. s. | * | n. s. |
| Con | Con + T | LCKD | LCKD + T | Significance | |||
|---|---|---|---|---|---|---|---|
| Training | Feed | Interaction | |||||
| Acat1 (Ketone body metabolism) | 1.00 ± 0.46 | 1.05 ± 0.44 | 1.30 ± 1.45 | 0.75 ± 0.23 | n. s. | n. s. | n. s. |
| Cd36 (Fatty acid transportation) | 1.00 ± 0.28 | 0.99 ± 0.29 | 0.70 ± 0.32 | 0.75 ± 0.26 | n. s. | n. s. | n. s. |
| Cox4 (Mitochondrial respiration) | 1.00 ± 0.30 | 0.97 ± 0.28 | 0.65 ± 0.29 | 0.79 ± 0.14 | n. s. | n. s. | n. s. |
| Cs (Citrate synthase) (Mitochondrial respiration) | 1.00 ± 0.23 | 1.19 ± 0.43 | 0.73 ± 0.32 | 1.02 ± 0.11 | n. s. | n. s. | n. s. |
| Cytochrome c (Mitochondrial respiration) | 1.00 ± 0.28 | 1.12 ± 0.44 | 0.90 ± 0.55 | 1.08 ± 0.28 | n. s. | n. s. | n. s. |
| Gp (Glycogen phosphatase) | 1.00 ± 0.25 | 1.21 ± 0.68 | 0.71 ± 0.35 | 0.52 ± 0.19 | n. s. | * | n. s. |
| Gs (Glycogen synthase) | 1.00 ± 0.40 | 0.83 ± 0.40 | 0.66 ± 0.53 | 0.65 ± 0.49 | n. s. | * | n. s. |
| Hadh (Fatty acid oxidation) | 1.00 ± 0.27 | 0.96 ± 0.37 | 1.27 ± 0.57 | 1.22 ± 0.36 | n. s. | n. s. | n. s. |
| Hbdh (Ketone body metabolism) | 1.00 ± 0.45 | 0.95 ± 0.44 | 0.55 ± 0.35 | 0.31 ± 0.18 | n. s. | n. s. | n. s. |
| Hk2 (Glycolysis) | 1.00 ± 0.11 | 1.16 ± 0.37 | 0.61 ± 0.28 | 0.60 ± 0.12 | n. s. | * | n. s. |
| Mct1 (Ketone body transportation) | 1.00 ± 0.22 | 0.93 ± 0.43 | 1.78 ± 0.84 | 1.34 ± 0.39 | n. s. | * | n. s. |
| Oxct1 (Ketone body metabolism) | 1.00 ± 1.10 | 0.53 ± 0.30 | 0.35 ± 0.22 | 0.34 ± 0.08 | n. s. | n. s. | n. s. |
| Sirt1 (Mitochondrial respiration) | 1.00 ± 0.39 | 0.79 ± 0.18 | 0.65 ± 0.15 | 0.62 ± 0.14 | n. s. | n. s. | n. s. |
| Tfam (Mitochondrial respiration) | 1.00 ± 0.27 | 1.09 ± 0.42 | 0.88 ± 0.30 | 0.97 ± 0.24 | n. s. | n. s. | n. s. |
| Con | Con + T | LCKD | LCKD + T | Significance | |||
|---|---|---|---|---|---|---|---|
| Training | Feed | Interaction | |||||
| Acat1 (Ketone body metabolism) | 1.00 ± 0.35 | 0.51 ± 0.16 | 0.70 ± 0.19 | 0.85 ± 0.28 | n. s. | n. s. | n. s. |
| Cox4 (Mitochondrial respiration) | 1.00 ± 0.33 | 0.62 ± 0.17 | 2.00 ± 0.33 | 2.51 ± 0.70 | n. s. | * | n. s. |
| Cpt1α (Fatty acid oxidation) | 1.00 ± 0.23 | 0.61 ± 0.21 | 1.35 ± 0.34 | 1.29 ± 0.37 b | n. s. | *** | n. s. |
| Hadh (Fatty acid oxidation) | 1.00 ± 0.59 | 0.67 ± 0.30 | 4.19 ± 0.90 | 4.28 ± 1.70 | n. s. | *** | n. s. |
| Hbdh (Ketone body metabolism) | 1.00 ± 0.51 | 0.70 ± 0.25 | 0.64 ± 0.06 | 0.60 ± 0.19 | n. s. | n. s. | n. s. |
| Hk2 (Glycolysis) | 1.00 ± 0.42 | 1.09 ± 0.13 | 0.93 ± 0.30 | 0.87 ± 0.27 | n. s. | n. s. | n. s. |
| Il-6 (Fatty acid mobilization) | 1.00 ± 0.57 | 1.05 ± 0.57 | 1.64 ± 1.04 | 1.35 ± 0.48 | n. s. | n. s. | n. s. |
| Oxct1 (Ketone body metabolism) | 1.00 ± 1.53 | 0.13 ± 0.06 | 0.34 ± 0.11 | 0.23 ± 0.06 | n. s. | n. s. | n. s. |
| Mcad (Fatty acid oxidation) | 1.00 ± 0.64 | 0.55 ± 0.31 | 3.75 ± 1.10 | 5.53 ± 2.75 | n. s. | *** | n. s. |
| Mct1 (Ketone body transportation) | 1.00 ± 0.85 | 1.32 ± 0.52 | 3.75 ± 1.30 | 2.67 ± 0.64 | n. s. | *** | n. s. |
| Tfam (Mitochondrial respiration) | 1.00 ± 0.53 | 0.82 ± 0.26 | 1.63 ± 0.50 | 1.38 ± 0.63 | n. s. | * | n. s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, S.; Yang, J.; Tominaga, T.; Liu, C.; Suzuki, K. A Low-Carbohydrate Ketogenic Diet and Treadmill Training Enhanced Fatty Acid Oxidation Capacity but Did Not Enhance Maximal Exercise Capacity in Mice. Nutrients 2021, 13, 611. https://doi.org/10.3390/nu13020611
Ma S, Yang J, Tominaga T, Liu C, Suzuki K. A Low-Carbohydrate Ketogenic Diet and Treadmill Training Enhanced Fatty Acid Oxidation Capacity but Did Not Enhance Maximal Exercise Capacity in Mice. Nutrients. 2021; 13(2):611. https://doi.org/10.3390/nu13020611
Chicago/Turabian StyleMa, Sihui, Jiao Yang, Takaki Tominaga, Chunhong Liu, and Katsuhiko Suzuki. 2021. "A Low-Carbohydrate Ketogenic Diet and Treadmill Training Enhanced Fatty Acid Oxidation Capacity but Did Not Enhance Maximal Exercise Capacity in Mice" Nutrients 13, no. 2: 611. https://doi.org/10.3390/nu13020611
APA StyleMa, S., Yang, J., Tominaga, T., Liu, C., & Suzuki, K. (2021). A Low-Carbohydrate Ketogenic Diet and Treadmill Training Enhanced Fatty Acid Oxidation Capacity but Did Not Enhance Maximal Exercise Capacity in Mice. Nutrients, 13(2), 611. https://doi.org/10.3390/nu13020611