
A Role for Peripheral Anandamide and 2-Arachidonoylglycerol in Short-Term Food Intake and Orexigenic Hypothalamic Responses in a Species with Continuous Nutrient Delivery


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Analyses of Plasma Endocannabinoids and Metabolites
2.3. Brain Sampling, mRNA Isolation and RT-qPCR
2.4. Immunohistochemistry (IHC)
2.5. Statistical Analyses
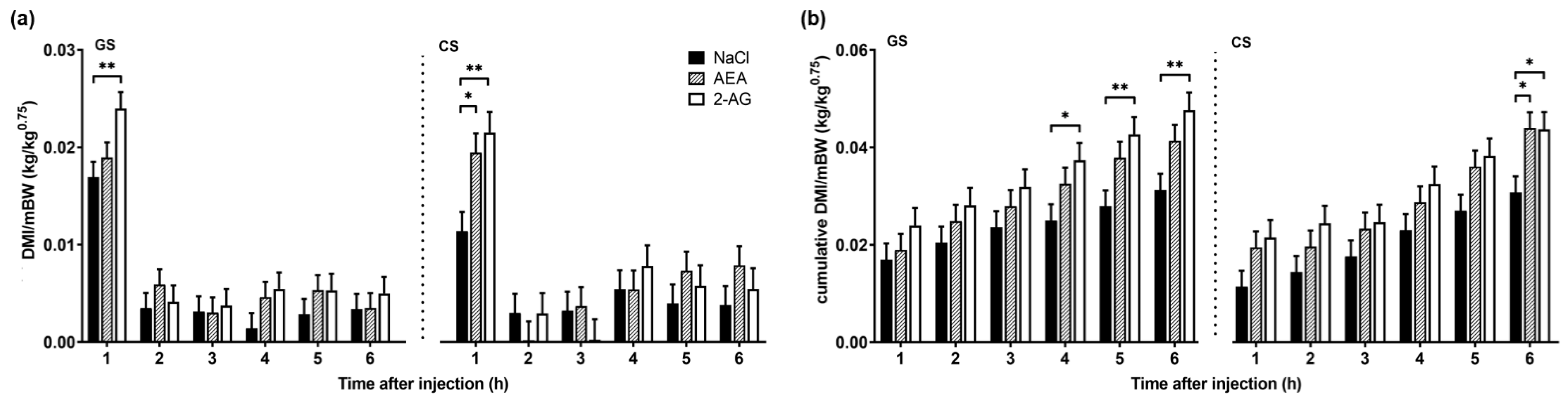
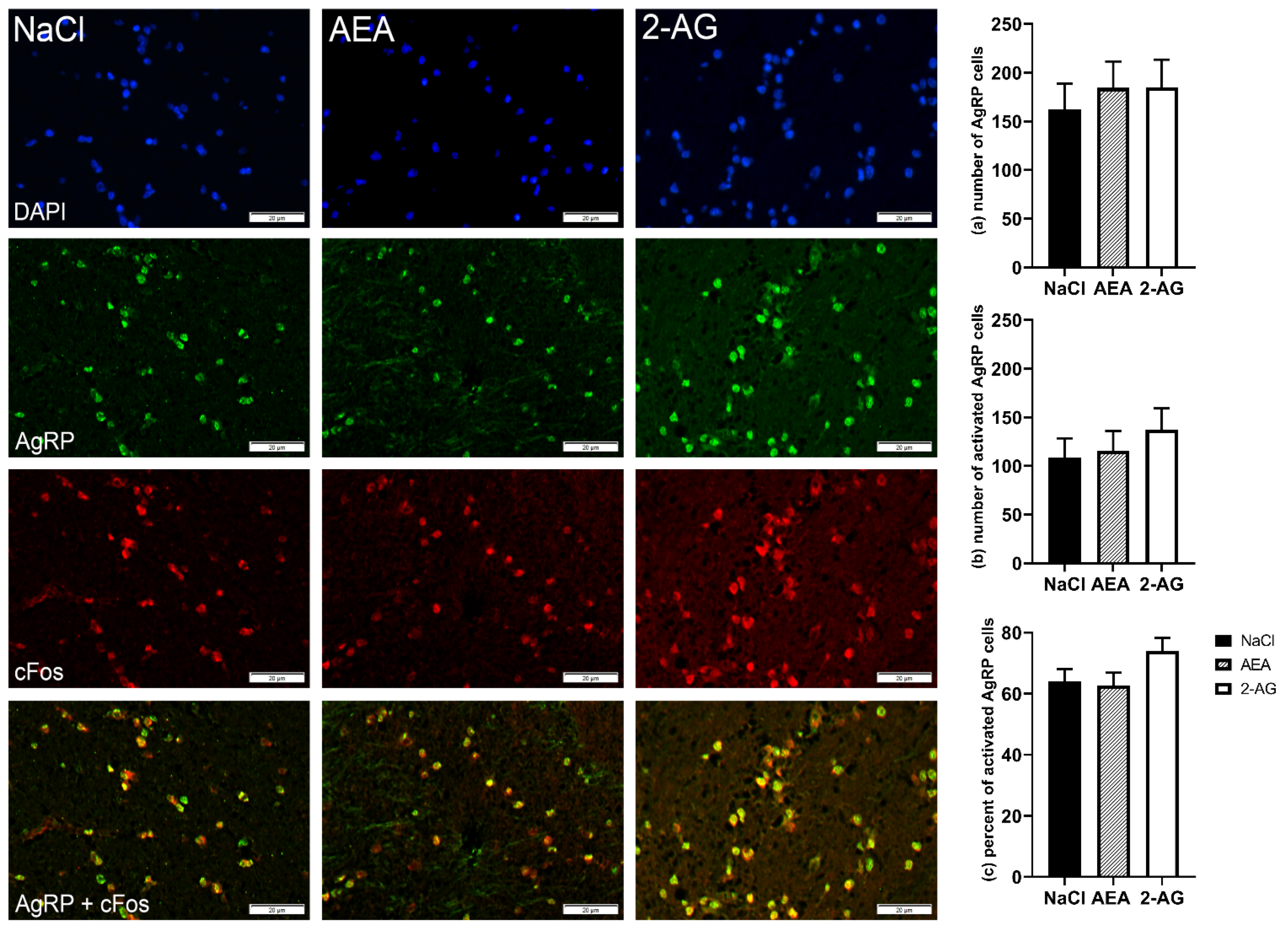
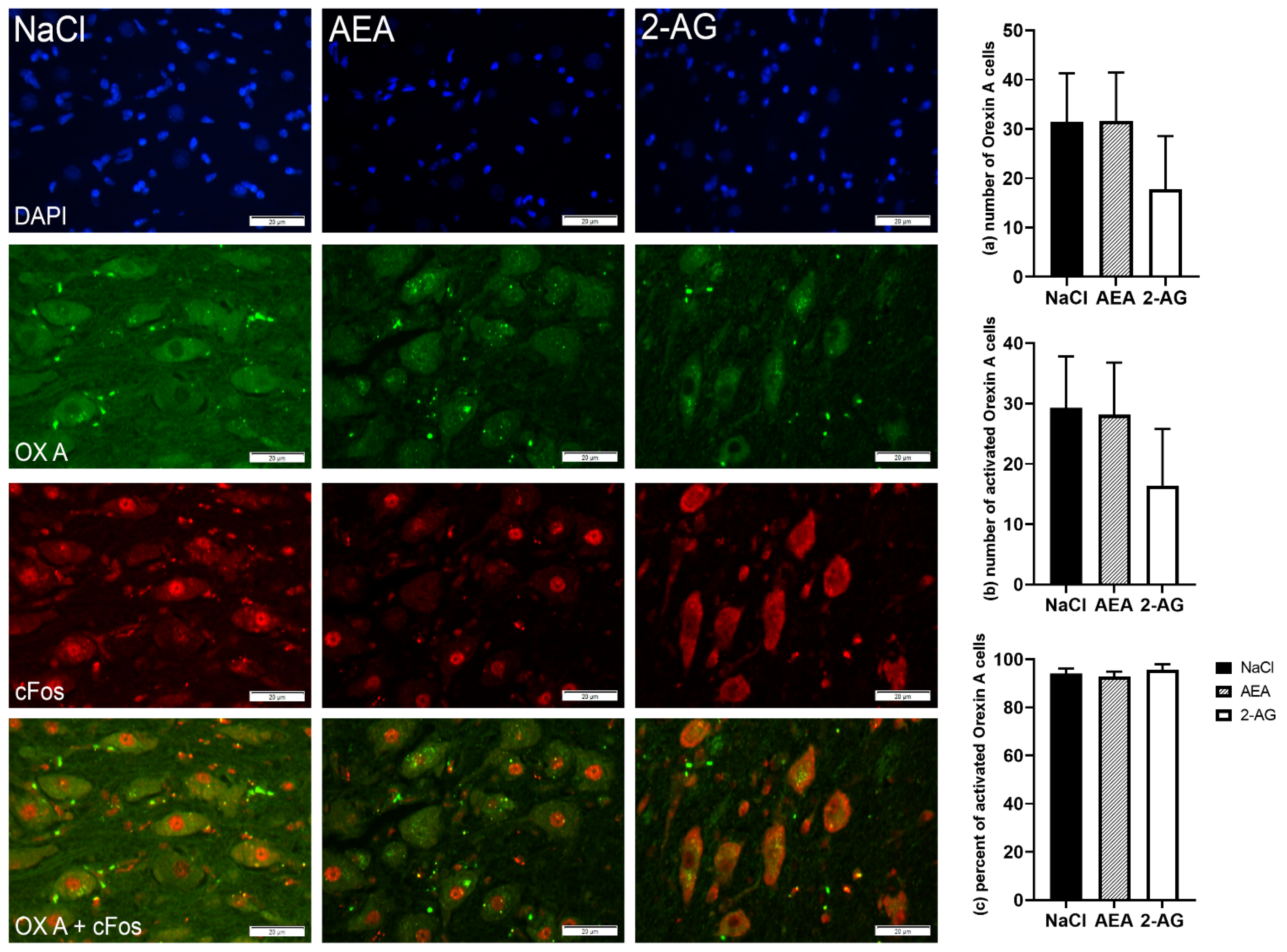
3. Results
3.1. Experiment 1
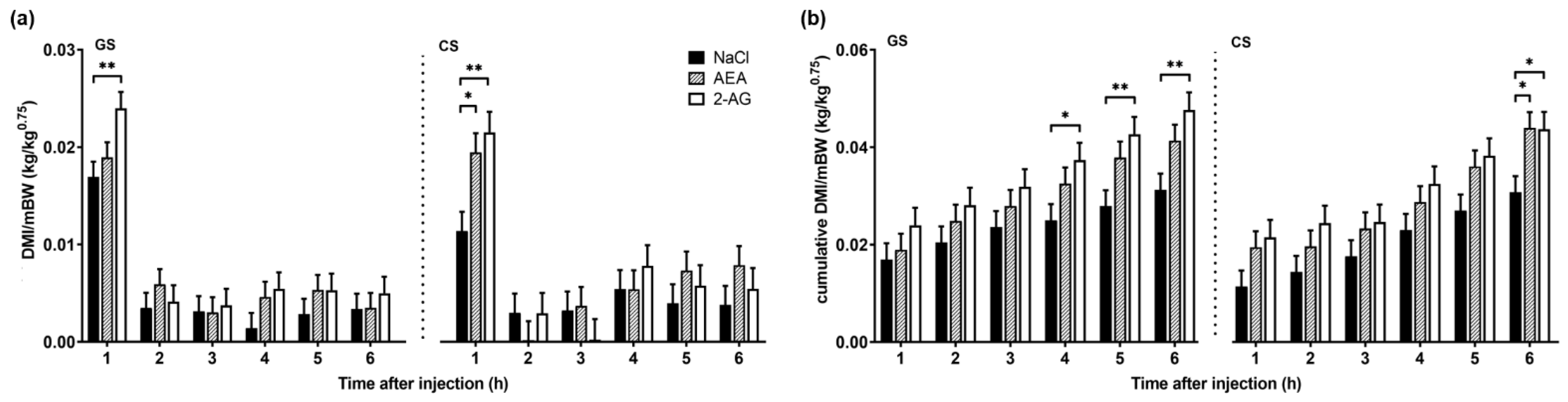
3.1.1. Body Weight, Milk Yield and Intake
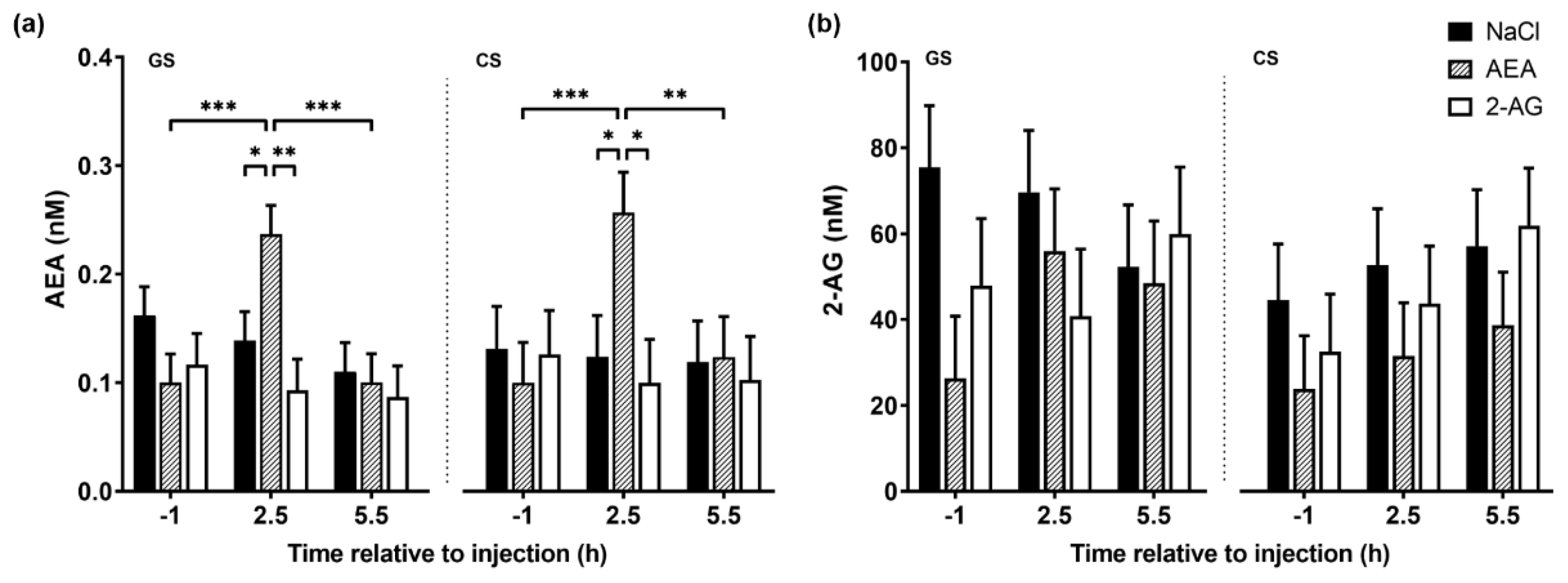
3.1.2. Plasma Endocannabinoid Concentrations
3.1.3. Transcription of Hypothalamic Genes
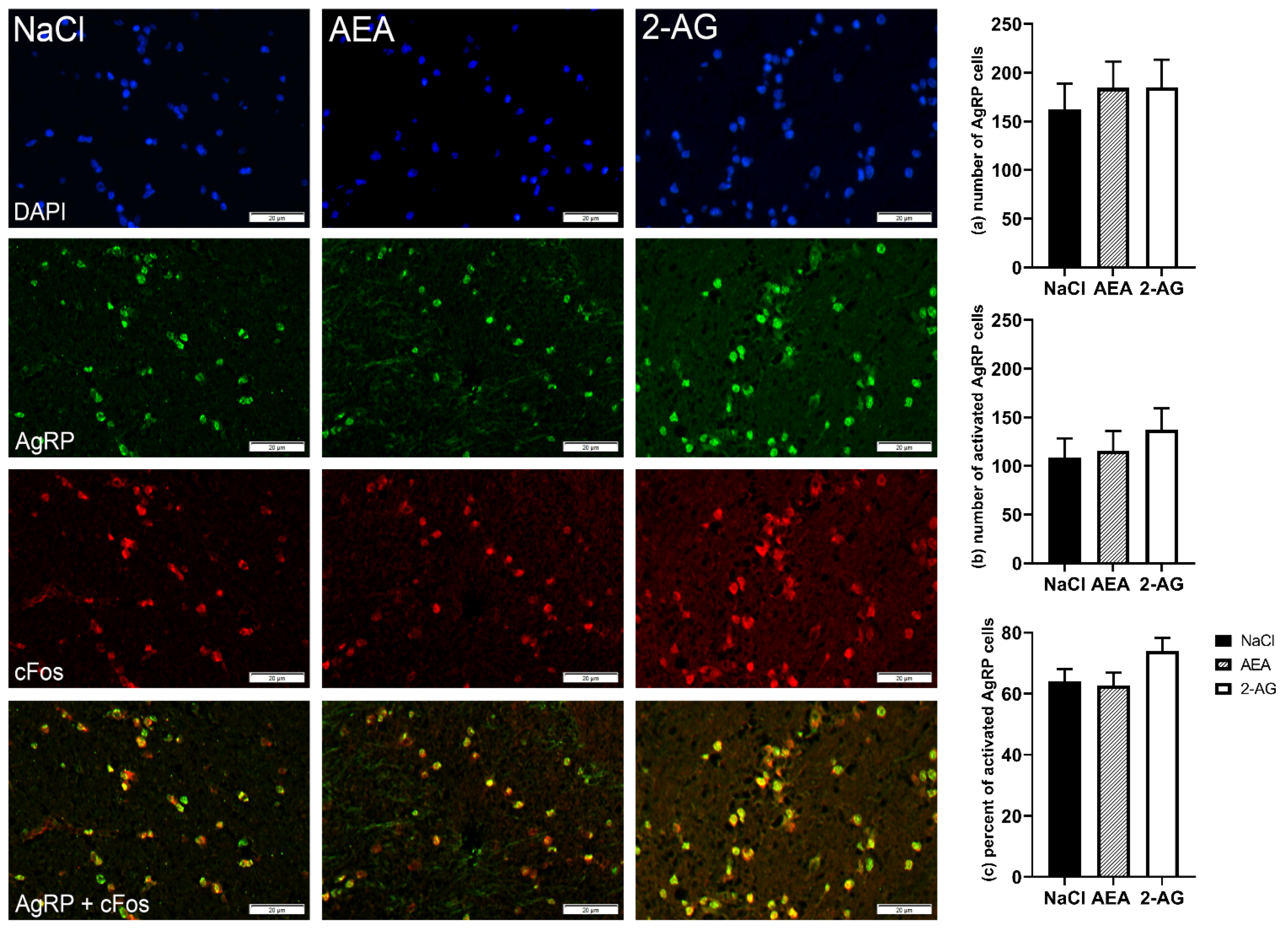
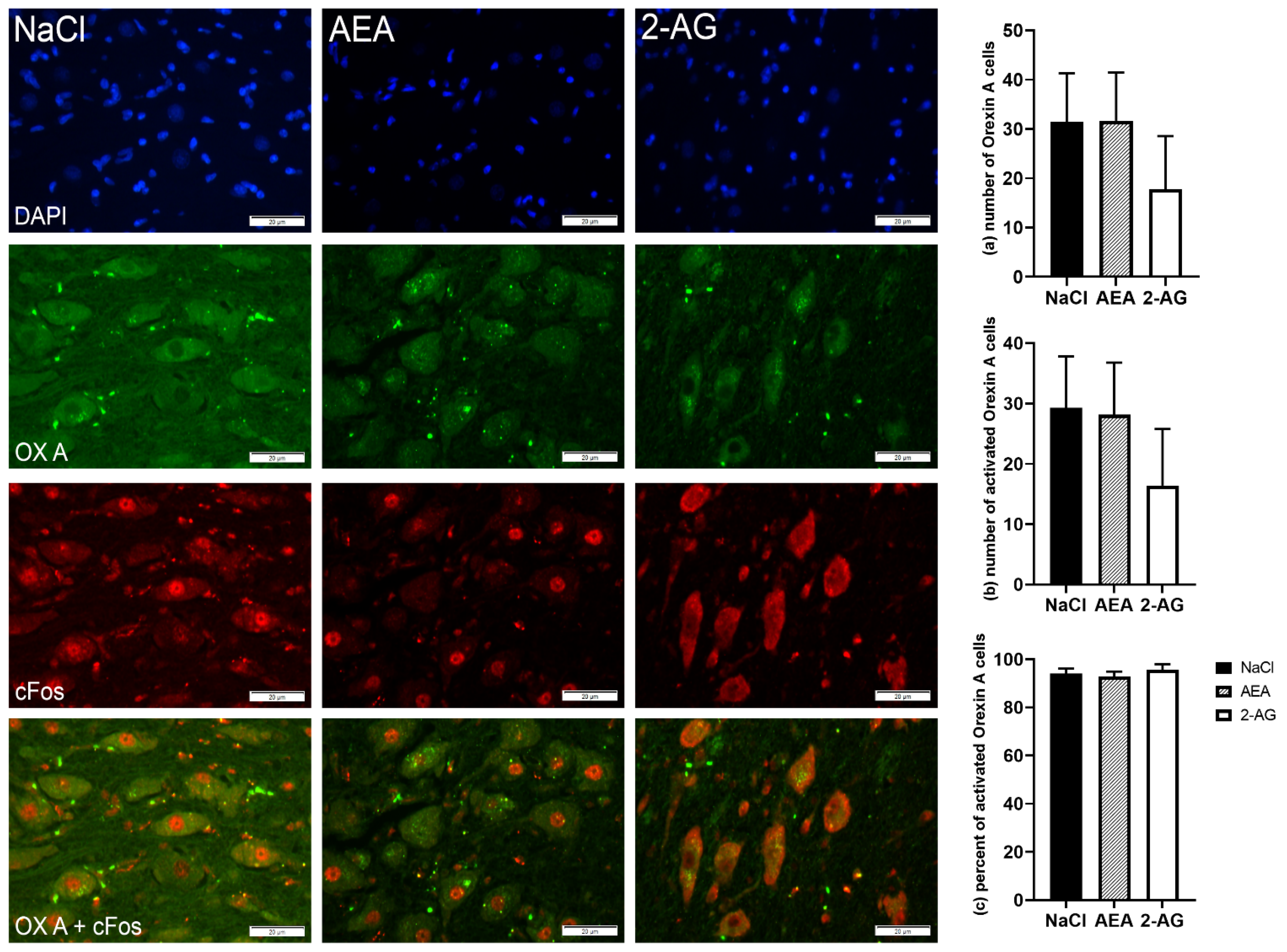
3.1.4. Immunohistochemistry
3.2. Experiment 2
Food Deprivation
4. Discussion
4.1. Effect of Endocannabinoids on Food Intake
4.2. Effect of Endocannabinoids on Plasma AEA and 2-AG Concentration
4.3. Effect of Endocannabinoids on Hypothalamic Orexigenic Circuits
4.4. Effect of Food Deprivation on Plasma Endocannabinoid Concentrations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwartz, M.W.; Woods, S.C.; Porte, D., Jr.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef]
- Wynne, K.; Stanley, S.; McGowan, B.; Bloom, S. Appetite control. J. Endocrinol. 2005, 184, 291–318. [Google Scholar] [CrossRef] [Green Version]
- Matias, I.; Bisogno, T.; Di Marzo, V. Endogenous cannabinoids in the brain and peripheral tissues: Regulation of their levels and control of food intake. Int. J. Obes. 2006, 30 (Suppl. S1), S7–S12. [Google Scholar] [CrossRef] [Green Version]
- Lau, B.K.; Cota, D.; Cristino, L.; Borgland, S.L. Endocannabinoid modulation of homeostatic and non-homeostatic feeding circuits. Neuropharmacology 2017, 124, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacol. Ther. 1997, 74, 129–180. [Google Scholar] [CrossRef]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-Arachidonoylglycerol: A possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar] [CrossRef]
- Di Marzo, V. Endocannabinoids: Synthesis and degradation. Rev. Physiol. Biochem. Pharmacol. 2008, 160, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Richard, D.; Guesdon, B.; Timofeeva, E. The brain endocannabinoid system in the regulation of energy balance. Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 17–32. [Google Scholar] [CrossRef]
- Burdyga, G.; Lal, S.; Varro, A.; Dimaline, R.; Thompson, D.G.; Dockray, G.J. Expression of cannabinoid CB1 receptors by vagal afferent neurons is inhibited by cholecystokinin. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 2708–2715. [Google Scholar] [CrossRef] [Green Version]
- DiPatrizio, N.V.; Astarita, G.; Schwartz, G.; Li, X.; Piomelli, D. Endocannabinoid signal in the gut controls dietary fat intake. Proc. Natl. Acad. Sci. USA 2011, 108, 12904–12908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdyga, G.; Varro, A.; Dimaline, R.; Thompson, D.G.; Dockray, G.J. Expression of cannabinoid CB1 receptors by vagal afferent neurons: Kinetics and role in influencing neurochemical phenotype. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G63–G69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiPatrizio, N.V. Endocannabinoids and the Gut-Brain Control of Food Intake and Obesity. Nutrients 2021, 13, 1214. [Google Scholar] [CrossRef] [PubMed]
- Bojesen, I.N.; Hansen, H.S. Binding of anandamide to bovine serum albumin. J. Lipid Res. 2003, 44, 1790–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oddi, S.; Fezza, F.; Pasquariello, N.; De Simone, C.; Rapino, C.; Dainese, E.; Finazzi-Agrò, A.; Maccarrone, M. Evidence for the intracellular accumulation of anandamide in adiposomes. Cell. Mol. Life Sci. CMLS 2008, 65, 840–850. [Google Scholar] [CrossRef]
- Maccarrone, M.; Dainese, E.; Oddi, S. Intracellular trafficking of anandamide: New concepts for signaling. Trends Biochem. Sci. 2010, 35, 601–608. [Google Scholar] [CrossRef]
- Willoughby, K.A.; Moore, S.F.; Martin, B.R.; Ellis, E.F. The biodisposition and metabolism of anandamide in mice. J. Pharmacol. Exp. Ther. 1997, 282, 243–247. [Google Scholar]
- Járai, Z.; Wagner, J.A.; Goparaju, S.K.; Wang, L.; Razdan, R.K.; Sugiura, T.; Zimmer, A.M.; Bonner, T.I.; Zimmer, A.; Kunos, G. Cardiovascular effects of 2-arachidonoyl glycerol in anesthetized mice. Hypertension 2000, 35, 679–684. [Google Scholar] [CrossRef] [Green Version]
- Di Marzo, V.; Melck, D.; Bisogno, T.; De Petrocellis, L. Endocannabinoids: Endogenous cannabinoid receptor ligands with neuromodulatory action. Trends Neurosci. 1998, 21, 521–528. [Google Scholar] [CrossRef]
- Hao, S.; Avraham, Y.; Mechoulam, R.; Berry, E.M. Low dose anandamide affects food intake, cognitive function, neurotransmitter and corticosterone levels in diet-restricted mice. Eur. J. Pharmacol. 2000, 392, 147–156. [Google Scholar] [CrossRef]
- Gomez, R.; Navarro, M.; Ferrer, B.; Trigo, J.M.; Bilbao, A.; Del Arco, I.; Cippitelli, A.; Nava, F.; Piomelli, D.; Rodriguez de Fonseca, F. A peripheral mechanism for CB1 cannabinoid receptor-dependent modulation of feeding. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 9612–9617. [Google Scholar] [CrossRef] [Green Version]
- Avraham, Y.; Paturski, I.; Magen, I.; Vorobiev, L.; Berry, E.M. 2-Arachidonoylglycerol as a possible treatment for anorexia nervosa in animal model in mice. Brain Res. 2017, 1670, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Wenger, T.; Jamali, K.A.; Juanéda, C.; Léonardelli, J.; Tramu, G. Arachidonyl ethanolamide (anandamide) activates the parvocellular part of hypothalamic paraventricular nucleus. Biochem. Biophys. Res. Commun. 1997, 237, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Ligresti, A.; Cristino, L. The endocannabinoid system as a link between homoeostatic and hedonic pathways involved in energy balance regulation. Int. J. Obes. 2009, 33 (Suppl. S2), S18–S24. [Google Scholar] [CrossRef] [Green Version]
- Gamber, K.M.; Macarthur, H.; Westfall, T.C. Cannabinoids augment the release of neuropeptide Y in the rat hypothalamus. Neuropharmacology 2005, 49, 646–652. [Google Scholar] [CrossRef]
- Morozov, Y.M.; Koch, M.; Rakic, P.; Horvath, T.L. Cannabinoid type 1 receptor-containing axons innervate NPY/AgRP neurons in the mouse arcuate nucleus. Mol. Metab. 2017, 6, 374–381. [Google Scholar] [CrossRef]
- Cristino, L.; Busetto, G.; Imperatore, R.; Ferrandino, I.; Palomba, L.; Silvestri, C.; Petrosino, S.; Orlando, P.; Bentivoglio, M.; Mackie, K.; et al. Obesity-driven synaptic remodeling affects endocannabinoid control of orexinergic neurons. Proc. Natl. Acad. Sci. USA 2013, 110, E2229–E2238. [Google Scholar] [CrossRef] [Green Version]
- Hilairet, S.; Bouaboula, M.; Carriere, D.; Le Fur, G.; Casellas, P. Hypersensitization of the Orexin 1 receptor by the CB1 receptor: Evidence for cross-talk blocked by the specific CB1 antagonist, SR141716. J. Biol. Chem. 2003, 278, 23731–23737. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.; Pediani, J.D.; Canals, M.; Milasta, S.; Milligan, G. Orexin-1 receptor-cannabinoid CB1 receptor heterodimerization results in both ligand-dependent and -independent coordinated alterations of receptor localization and function. J. Biol. Chem. 2006, 281, 38812–38824. [Google Scholar] [CrossRef] [Green Version]
- Imperatore, R.; D’Angelo, L.; Safari, O.; Motlagh, H.A.; Piscitelli, F.; de Girolamo, P.; Cristino, L.; Varricchio, E.; di Marzo, V.; Paolucci, M. Overlapping Distribution of Orexin and Endocannabinoid Receptors and Their Functional Interaction in the Brain of Adult Zebrafish. Front. Neuroanat. 2018, 12, 62. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.S.; Bradford, B.J.; Harvatine, K.J. The cow as a model to study food intake regulation. Annu. Rev. Nutr. 2005, 25, 523–547. [Google Scholar] [CrossRef] [PubMed]
- Kuhla, B.; Kuhla, S.; Rudolph, P.E.; Albrecht, D.; Metges, C.C. Proteomics analysis of hypothalamic response to energy restriction in dairy cows. Proteomics 2007, 7, 3602–3617. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.J.; Graugnard, D.E.; Loor, J.J. Endocannabinoid system and proopiomelanocortin gene expression in peripartal bovine liver in response to prepartal plane of nutrition. J. Anim. Physiol. Anim. Nutr. 2012, 96, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Zachut, M.; Kra, G.; Moallem, U.; Livshitz, L.; Levin, Y.; Udi, S.; Nemirovski, A.; Tam, J. Characterization of the endocannabinoid system in subcutaneous adipose tissue in periparturient dairy cows and its association to metabolic profiles. PLoS ONE 2018, 13, e0205996. [Google Scholar] [CrossRef]
- Kuhla, B.; Kaever, V.; Tuchscherer, A.; Kuhla, A. Involvement of plasma endocannabinoids and the hypothalamic endocannabinoid system in increasing feed intake after parturition of dairy cows. Neuroendocrinology 2019, 110, 246–257. [Google Scholar] [CrossRef]
- Sulcova, E.; Mechoulam, R.; Fride, E. Biphasic effects of anandamide. Pharmacol. Biochem. Behav. 1998, 59, 347–352. [Google Scholar] [CrossRef]
- Long, J.Z.; Li, W.; Booker, L.; Burston, J.J.; Kinsey, S.G.; Schlosburg, J.E.; Pavón, F.J.; Serrano, A.M.; Selley, D.E.; Parsons, L.H.; et al. Selective blockade of 2-arachidonoylglycerol hydrolysis produces cannabinoid behavioral effects. Nat. Chem. Biol. 2009, 5, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoerner, A.A.; Batkai, S.; Suchy, M.T.; Gutzki, F.M.; Engeli, S.; Jordan, J.; Tsikas, D. Simultaneous UPLC-MS/MS quantification of the endocannabinoids 2-arachidonoyl glycerol (2AG), 1-arachidonoyl glycerol (1AG), and anandamide in human plasma: Minimization of matrix-effects, 2AG/1AG isomerization and degradation by toluene solvent extraction. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 883–884, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef] [Green Version]
- Van Ackern, I.; Wulf, R.; Dannenberger, D.; Tuchscherer, A.; Kuhla, B. Effects of endocannabinoids on feed intake, stress response and whole-body energy metabolism in dairy cows. 2021. to be submitted. [Google Scholar]
- Kirkham, T.C.; Williams, C.M.; Fezza, F.; Di Marzo, V. Endocannabinoid levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding and satiation: Stimulation of eating by 2-arachidonoyl glycerol. Br. J. Pharmacol. 2002, 136, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Koch, M. Cannabinoid Receptor Signaling in Central Regulation of Feeding Behavior: A Mini-Review. Front. Neurosci. 2017, 11, 293. [Google Scholar] [CrossRef]
- Higgs, S.; Williams, C.M.; Kirkham, T.C. Cannabinoid influences on palatability: Microstructural analysis of sucrose drinking after delta(9)-tetrahydrocannabinol, anandamide, 2-arachidonoyl glycerol and SR141716. Psychopharmacology 2003, 165, 370–377. [Google Scholar] [CrossRef]
- DiPatrizio, N.V.; Simansky, K.J. Activating parabrachial cannabinoid CB1 receptors selectively stimulates feeding of palatable foods in rats. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 9702–9709. [Google Scholar] [CrossRef]
- Soria-Gómez, E.; Bellocchio, L.; Reguero, L.; Lepousez, G.; Martin, C.; Bendahmane, M.; Ruehle, S.; Remmers, F.; Desprez, T.; Matias, I.; et al. The endocannabinoid system controls food intake via olfactory processes. Nat. Neurosci. 2014, 17, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Al Shoyaib, A.; Archie, S.R.; Karamyan, V.T. Intraperitoneal Route of Drug Administration: Should it Be Used in Experimental Animal Studies? Pharm. Res. 2019, 37, 12. [Google Scholar] [CrossRef]
- Kozak, K.R.; Crews, B.C.; Ray, J.L.; Tai, H.H.; Morrow, J.D.; Marnett, L.J. Metabolism of prostaglandin glycerol esters and prostaglandin ethanolamides in vitro and in vivo. J. Biol. Chem. 2001, 276, 36993–36998. [Google Scholar] [CrossRef] [Green Version]
- Storr, M.A.; Sharkey, K.A. The endocannabinoid system and gut-brain signalling. Curr. Opin. Pharmacol. 2007, 7, 575–582. [Google Scholar] [CrossRef]
- Fride, E.; Mechoulam, R. Pharmacological activity of the cannabinoid receptor agonist, anandamide, a brain constituent. Eur. J. Pharmacol. 1993, 231, 313–314. [Google Scholar] [CrossRef]
- Soria-Gómez, E.; Matias, I.; Rueda-Orozco, P.E.; Cisneros, M.; Petrosino, S.; Navarro, L.; Di Marzo, V.; Prospéro-García, O. Pharmacological enhancement of the endocannabinoid system in the nucleus accumbens shell stimulates food intake and increases c-Fos expression in the hypothalamus. Br. J. Pharmacol. 2007, 151, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.H.; Chen, Y.J.; Chua, S.C., Jr.; Talmage, D.A.; Role, L.W. Integration of endocannabinoid and leptin signaling in an appetite-related neural circuit. Neuron 2005, 48, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Imperatore, R.; Palomba, L.; Morello, G.; Spiezio, A.D.; Piscitelli, F.; Marzo, V.D.; Cristino, L. Formation of OX-1R/CB1R heteromeric complexes in embryonic mouse hypothalamic cells: Effect on intracellular calcium, 2-arachidonoyl-glycerol biosynthesis and ERK phosphorylation. Pharmacol. Res. 2016, 111, 600–609. [Google Scholar] [CrossRef]
- Merroun, I.; El Mlili, N.; Martinez, R.; Porres, J.M.; Llopis, J.; Ahabrach, H.; Aranda, P.; Sanchez Gonzalez, C.; Errami, M.; Lopez-Jurado, M. Interaction between orexin A and cannabinoid system in the lateral hypothalamus of rats and effects of subchronic intraperitoneal administration of cannabinoid receptor inverse agonist on food intake and the nutritive utilization of protein. J. Physiol. Pharmacol. 2015, 66, 181–190. [Google Scholar] [PubMed]
- Morello, G.; Imperatore, R.; Palomba, L.; Finelli, C.; Labruna, G.; Pasanisi, F.; Sacchetti, L.; Buono, L.; Piscitelli, F.; Orlando, P.; et al. Orexin-A represses satiety-inducing POMC neurons and contributes to obesity via stimulation of endocannabinoid signaling. Proc. Natl. Acad. Sci. USA 2016, 113, 4759–4764. [Google Scholar] [CrossRef] [Green Version]
- DiPatrizio, N.V.; Igarashi, M.; Narayanaswami, V.; Murray, C.; Gancayco, J.; Russell, A.; Jung, K.M.; Piomelli, D. Fasting stimulates 2-AG biosynthesis in the small intestine: Role of cholinergic pathways. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R805–R813. [Google Scholar] [CrossRef] [Green Version]
- Matias, I.; Gonthier, M.P.; Orlando, P.; Martiadis, V.; De Petrocellis, L.; Cervino, C.; Petrosino, S.; Hoareau, L.; Festy, F.; Pasquali, R.; et al. Regulation, function, and dysregulation of endocannabinoids in models of adipose and beta-pancreatic cells and in obesity and hyperglycemia. J. Clin. Endocrinol. Metab. 2006, 91, 3171–3180. [Google Scholar] [CrossRef]
- Martín-Pérez, C.; Contreras-Rodríguez, O.; Pastor, A.; Christensen, E.; Andrews, Z.B.; de la Torre, R.; Verdejo-García, A. Endocannabinoid signaling of homeostatic status modulates functional connectivity in reward and salience networks. Psychopharmacology 2021. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M.; Engeli, S.; Klöting, N.; Berndt, J.; Fasshauer, M.; Bátkai, S.; Pacher, P.; Schön, M.R.; Jordan, J.; Stumvoll, M. Dysregulation of the peripheral and adipose tissue endocannabinoid system in human abdominal obesity. Diabetes 2006, 55, 3053–3060. [Google Scholar] [CrossRef] [Green Version]
- Gatta-Cherifi, B.; Matias, I.; Vallee, M.; Tabarin, A.; Marsicano, G.; Piazza, P.V.; Cota, D. Simultaneous postprandial deregulation of the orexigenic endocannabinoid anandamide and the anorexigenic peptide YY in obesity. Int. J. Obes. 2012, 36, 880–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanus, L.; Avraham, Y.; Ben-Shushan, D.; Zolotarev, O.; Berry, E.M.; Mechoulam, R. Short-term fasting and prolonged semistarvation have opposite effects on 2-AG levels in mouse brain. Brain Res. 2003, 983, 144–151. [Google Scholar] [CrossRef]
- Bobe, G.; Young, J.W.; Beitz, D.C. Invited review: Pathology, etiology, prevention, and treatment of fatty liver in dairy cows. J. Dairy Sci. 2004, 87, 3105–3124. [Google Scholar] [CrossRef] [Green Version]
- Mallipedhi, A.; Prior, S.L.; Dunseath, G.; Bracken, R.M.; Barry, J.; Caplin, S.; Eyre, N.; Morgan, J.; Baxter, J.N.; O’Sullivan, S.E.; et al. Changes in plasma levels of N-arachidonoyl ethanolamine and N-palmitoylethanolamine following bariatric surgery in morbidly obese females with impaired glucose homeostasis. J. Diabetes Res. 2015, 2015, 680867. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | NaCl | AEA | 2-AG | ANOVA |
|---|---|---|---|---|
| ARC | ||||
| AGRP | 1.72 ± 0.46 | 0.55 ± 0.04 | 2.03 ± 0.87 | F(2,17) = 3.13, p = 0.07 |
| CNR1 | 1.03 ± 0.16 | 1.06 ± 0.17 | 1.11 ± 0.14 | F(2,17) = 0.06, p = 0.95 |
| DAGLA | 1.00 ± 0.09 | 0.98 ± 0.09 | 1.07 ± 0.07 | F(2,17) = 0.29, p = 0.75 |
| FAAH | 1.07 ± 0.18 | 0.98 ± 0.10 | 1.12 ± 0.07 | F(2,17) = 0.31, p = 0.73 |
| NAPELD | 1.05 ± 0.03 | 0.95 ± 0.11 | 1.07 ± 0.11 | F(2,17) = 0.53, p = 0.60 |
| NPY * | 1.84 ± 0.70 | 0.42 ± 0.11 | 1.91 ± 0.48 | F(2,15) = 2.81, p = 0.09 |
| PLAAT5 * | 1.16 ± 0.45 | 1.43 ± 0.42 | 1.05 ± 0.23 | F(2,14) = 0.39, p = 0.68 |
| PVN | ||||
| AGRP | 1.17 ± 0.17 | 1.15 ± 0.38 | 1.48 ± 0.47 | F(2,17) = 0.24, p = 0.79 |
| CNR1 | 0.91 ± 0.07 | 1.07 ± 0.24 | 1.37 ± 0.24 | F(2,17) = 1.12, p = 0.35 |
| DAGLA | 0.99 ± 0.06 | 1.06 ± 0.10 | 1.00 ± 0.14 | F(2,17) = 0.16, p = 0.85 |
| FAAH | 0.99 ± 0.10 | 1.00 ± 0.07 | 1.08 ± 0.09 | F(2,17) = 0.29, p = 0.75 |
| NAPELD | 1.01 ± 0.09 | 1.06 ± 0.09 | 1.00 ± 0.08 | F(2,17) = 0.12, p = 0.89 |
| PLAAT5 | 1.38 ± 0.24 | 1.26 ± 0.46 | 1.65 ± 0.66 | F(2,17) = 0.18, p = 0.84 |
| Ad Libitum | Food Deprivation | Refeeding | ANOVA | |
|---|---|---|---|---|
| AEA (nM) | 0.26 a ± 0.02 | 0.29 a ± 0.02 | 0.27 a ± 0.02 | F(2,12) = 0.69, p = 0.52 |
| 2-AG (nM) | 3.60 a ± 0.49 | 4.73 b ± 0.49 | 3.63 a ± 0.49 | F(2,12) = 6.17, p < 0.05 |
| NEFA (µmol/L) | 162.71 a ± 40.28 | 237.14 b ± 40.28 | 189.29 ab ± 40.28 | F(2,12) = 7.46, p < 0.01 |
| Glucose (mmol/L) | 4.06 a ± 0.13 | 3.65 b ± 0.13 | 3.73 b ± 0.13 | F(2,12) = 8.38, p < 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Ackern, I.; Kuhla, A.; Kuhla, B. A Role for Peripheral Anandamide and 2-Arachidonoylglycerol in Short-Term Food Intake and Orexigenic Hypothalamic Responses in a Species with Continuous Nutrient Delivery. Nutrients 2021, 13, 3587. https://doi.org/10.3390/nu13103587
van Ackern I, Kuhla A, Kuhla B. A Role for Peripheral Anandamide and 2-Arachidonoylglycerol in Short-Term Food Intake and Orexigenic Hypothalamic Responses in a Species with Continuous Nutrient Delivery. Nutrients. 2021; 13(10):3587. https://doi.org/10.3390/nu13103587
Chicago/Turabian Stylevan Ackern, Isabel, Angela Kuhla, and Björn Kuhla. 2021. "A Role for Peripheral Anandamide and 2-Arachidonoylglycerol in Short-Term Food Intake and Orexigenic Hypothalamic Responses in a Species with Continuous Nutrient Delivery" Nutrients 13, no. 10: 3587. https://doi.org/10.3390/nu13103587
APA Stylevan Ackern, I., Kuhla, A., & Kuhla, B. (2021). A Role for Peripheral Anandamide and 2-Arachidonoylglycerol in Short-Term Food Intake and Orexigenic Hypothalamic Responses in a Species with Continuous Nutrient Delivery. Nutrients, 13(10), 3587. https://doi.org/10.3390/nu13103587