
Eggshell Membrane Ameliorates Hyperuricemia by Increasing Urate Excretion in Potassium Oxonate-Injected Rats



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Hyperuricemia Induction and EM Administration
2.3. Collection of Blood, Urine, and Tissue Samples
2.4. Analysis of Uric Acid Concentrations in Serum and Urine
2.5. Measurement of Proinflammatory Cytokines Levels in Serum
2.6. In Vitro Xanthine Oxidase Activity from Liver and Serum
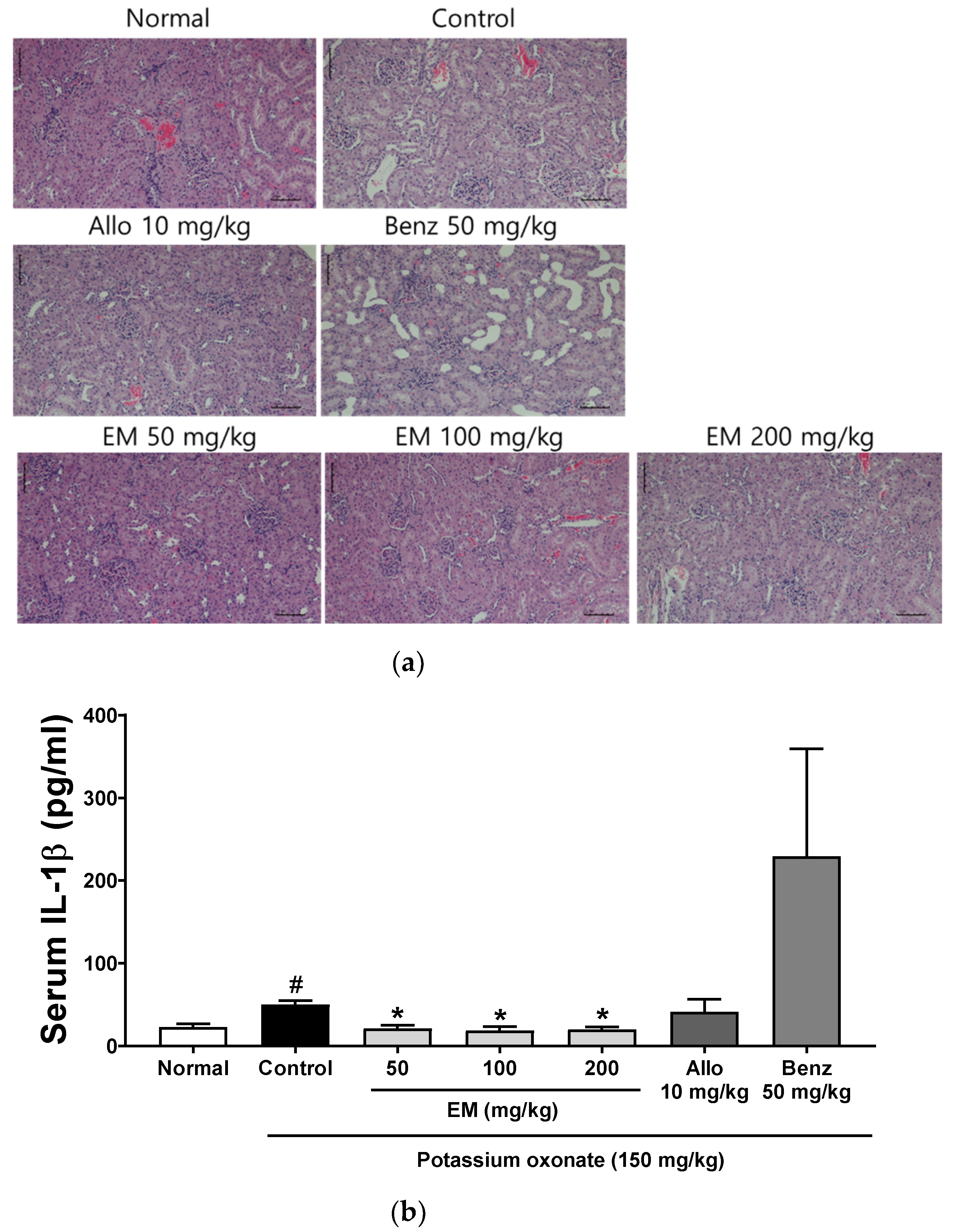
2.7. Kidney Histopathological Examination
2.8. Western Blot Analysis
2.9. Measurement of Urate Uptake Using UART1-Expressing Oocytes
2.10. Statistics
3. Results
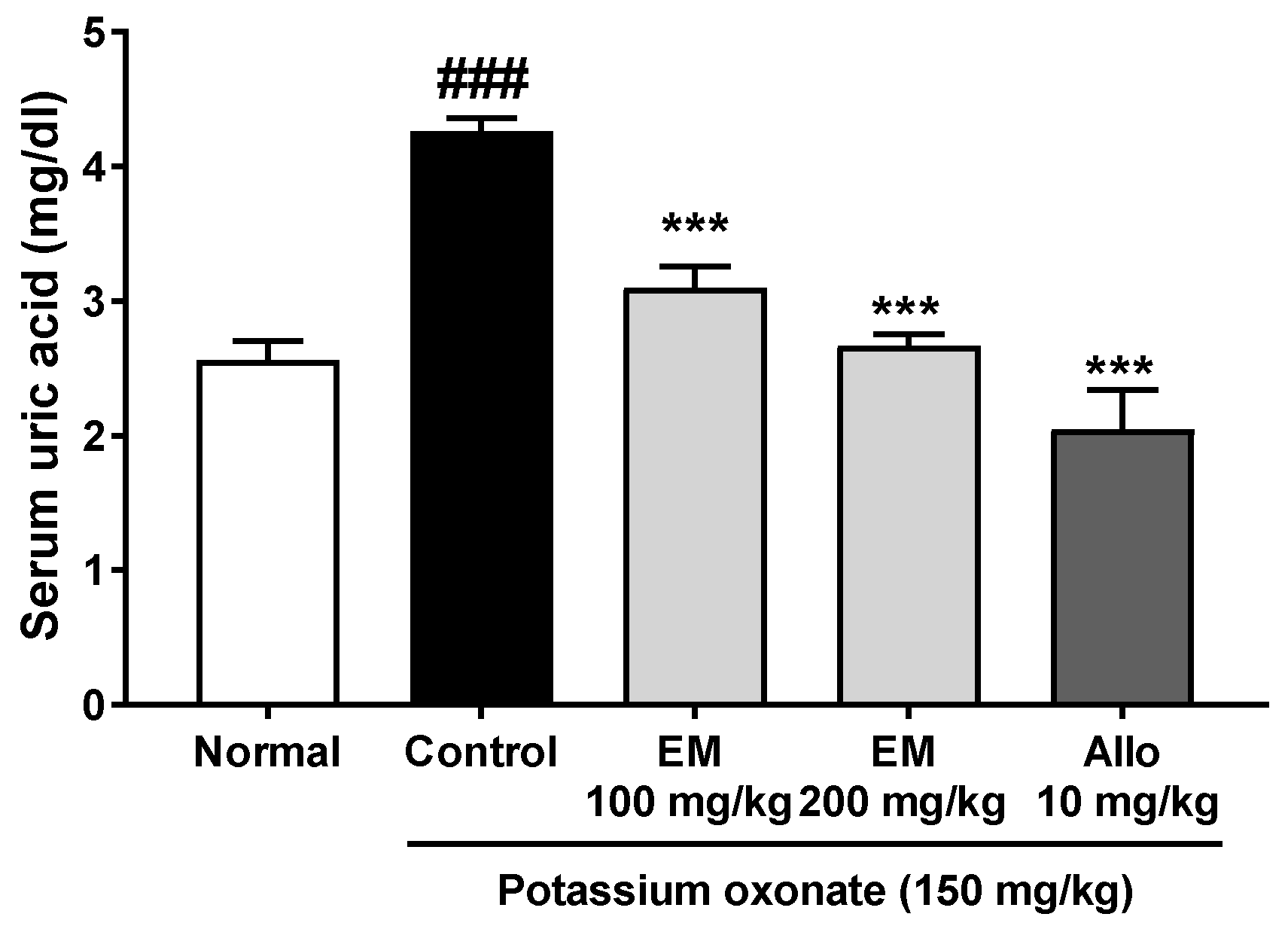
3.1. Effects of Eggshell Membrane on Serum Uric Acid Secretion
3.2. Effects of Eggshell Membrane on Serum and Urine Uric Acid Levels
3.3. Effects of Eggshell Membrane on Renal Inflammation
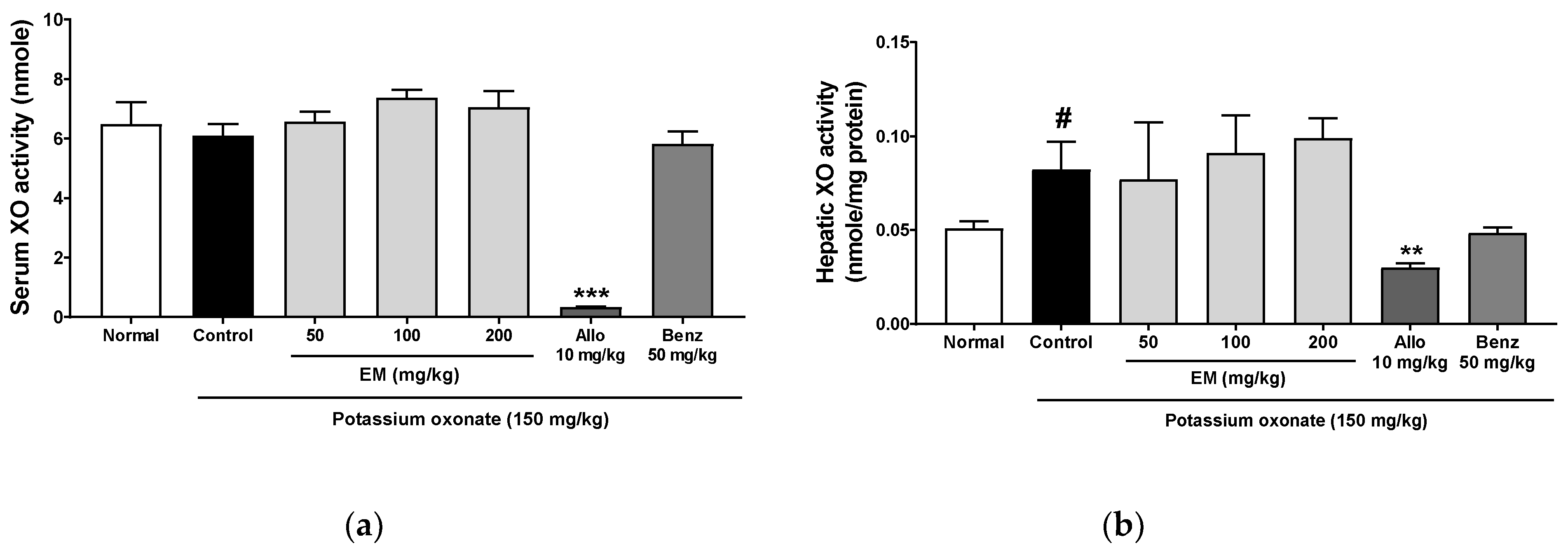
3.4. Effects of Eggshell Membrane on XO Activity
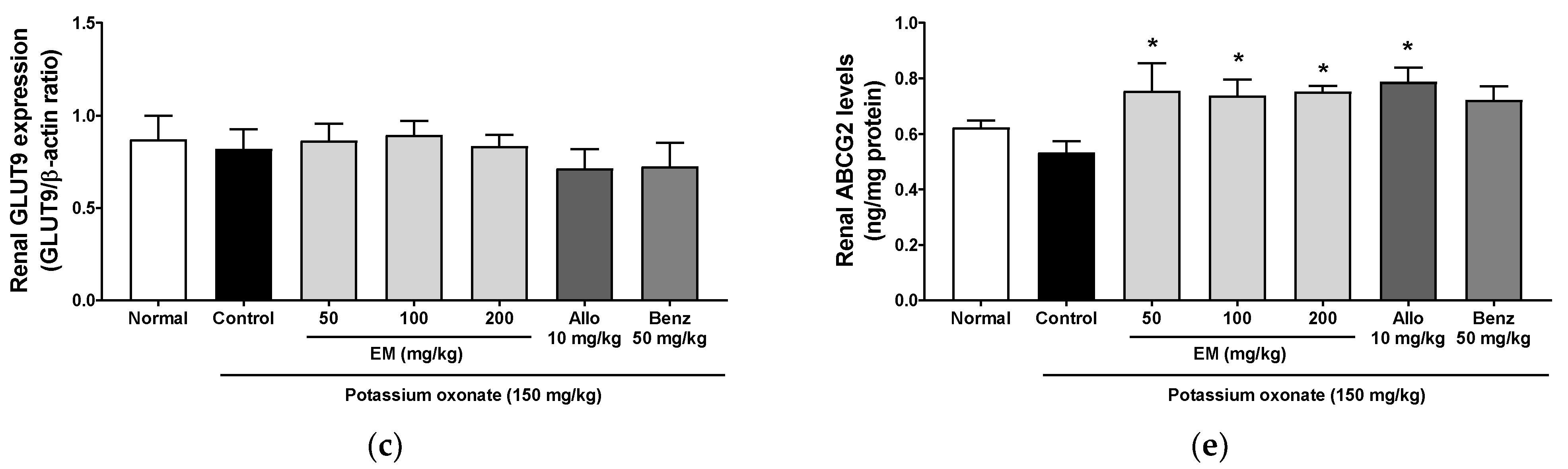
3.5. Effects of Eggshell Membrane on Urate Excretion
3.6. Effect of EM on In Vitro URAT1 Uptake in Oocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leach, R.M., Jr.; Rucker, R.B.; Van Dyke, G.P. Egg shell membrane protein: A nonelastin desmosine/isodesmosine-containing protein. Arch Biochem. Biophys. 1981, 207, 353–359. [Google Scholar] [CrossRef]
- Okubo, T.; Akachi, S.; Hatta, H. Structure of hen eggs and physiology of egg laying. In Hen Eggs: Their Basic and Applied Science; Yamamoto, T., Juneja, L.R., Hatta, H., Kim, M., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 1–12. [Google Scholar]
- Jeon, T.W.; Park, K.M. Functional properties of egg shell membrane hydrolysate as a food material. J. Korean Soc. Food Sci. Anim. Resour. 2002, 22, 267–273. [Google Scholar]
- Ruff, K.J.; DeVore, D.P. Reduction of pro-inflammatory cytokines in rats following 7-day oral supplementation with a proprietary eggshell membrane-derived product. Mod. Res. Inflamm. 2014, 3, 19–25. [Google Scholar] [CrossRef]
- Ruff, K.J.; Winkler, A.; Jackson, R.W.; DeVore, D.P.; Ritz, B.W. Eggshell membrane in the treatment of pain and stiffness from osteoarthritis of the knee: A randomized, multicenter, double-blind, placebo-controlled clinical study. Clin. Rheumatol. 2009, 28, 907–914. [Google Scholar] [CrossRef] [Green Version]
- Wedekind, K.J.; Ruff, K.J.; Atwell, C.A.; Evans, J.L.; Bendele, A.M. Beneficial effects of natural eggshell membrane (NEM) on multiple indices of arthritis in collagen-induced arthritic rats. Mod. Rheumatol. 2017, 27, 838–848. [Google Scholar] [CrossRef] [Green Version]
- Sim, B.Y.; Bak, J.W.; Lee, H.J.; Jun, J.A.; Choi, H.J.; Kwon, C.J.; Kim, H.Y.; Kevin, J.R.; Karsten, B.; Kim, D.H. Effects of natural eggshell membrane (NEM) on monosodium iodoacetate-induced arthritis in rats. J. Nutr. Health 2015, 48, 310–318. [Google Scholar] [CrossRef]
- Danesch, U.; Seybold, M.; Rittinghausen, R.; Treibel, W.; Bitterlich, N. NEM brand eggshell membrane effective in the treatment of pain associated with knee and hip osteoarthritis: Results from a six center, open label German clinical study. J. Arthritis 2014, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.A.; Reddy, S.G.; Kundukulam, J. Risk factors for gout and prevention: A systematic review of the literature. Curr. Opin. Rheumatol. 2011, 23, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Ma, C.H.; Zhou, F.; Kong, L.D. Siwu decoction attenuates oxonate-induced hyperuricemia and kidney inflammation in mice. Chin. J. Nat. Med. 2016, 14, 499–507. [Google Scholar] [CrossRef]
- Wu, A.H.; Gladden, J.D.; Ahmed, M.; Ahmed, A.; Filippatos, G. Relation of serum uric acid to cardiovascular disease. Int. J. Cardiol. 2016, 213, 4–7. [Google Scholar] [CrossRef]
- Su, J.; Wei, Y.; Liu, M.; Liu, T.; Li, J.; Ji, Y.; Liang, J. Anti-hyperuricemic and nephroprotective effects of Rhizoma Dioscoreae septemlobae extracts and its main component dioscin via regulation of mOAT1, mURAT1 and mOCT2 in hypertensive mice. Arch. Pharm. Res. 2014, 37, 1336–1344. [Google Scholar] [CrossRef]
- Benn, C.L.; Dua, P.; Gurrell, R.; Loudon, P.; Pike, A.; Storer, R.I.; Vangjeli, C. Physiology of hyperuricemia and urate-lowering treatments. Front. Med. 2018, 5, 160. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Lu, J.M.; Yao, Q. Hyperuricemia-Related Diseases and Xanthine Oxidoreductase (XOR) Inhibitors: An Overview. Med. Sci. Monit. 2016, 22, 2501–2512. [Google Scholar] [CrossRef] [Green Version]
- Chohan, S. Safety and efficacy of febuxostat treatment in subjects with gout and severe allopurinol adverse reactions. J. Rheumatol. 2011, 38, 1957–1959. [Google Scholar] [CrossRef]
- Strilchuk, L.; Fogacci, F.; Cicero, A.F. Safety and tolerability of available urate-lowering drugs: A critical review. Expert Opin. Drug Saf. 2019, 18, 261–271. [Google Scholar] [CrossRef]
- Azevedo, V.F.; Kos, I.A.; Vargas-Santos, A.B.; da Rocha Castelar Pinheiro, G.; Dos Santos Paiva, E. Benzbromarone in the treatment of gout. Adv. Rheumatol. 2019, 59, 37. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.H.; Ye, Y.S.; Wang, C.Y.; Li, Z.L.; Zheng, H.; Ma, K.L. Potassium oxonate induces acute hyperuricemia in the tree shrew (tupaia belangeri chinensis). Exp. Anim. 2017, 66, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Hall, H.; Scoville, J.P.; Reynolds, D.J.; Simlot, R.; Duncan, P. Substituted cyclic imides as potential anti-gout agents. Life Sci. 1990, 46, 1923–1927. [Google Scholar]
- Lee, Y.S.; Kim, S.H.; Yuk, H.J.; Kim, D.S. DKB114, A Mixture of Chrysanthemum Indicum Linne Flower and Cinnamomum Cassia (L.) J. Presl bark extracts, improves hyperuricemia through inhibition of xanthine oxidase activity and increasing urine excretion. Nutrients 2018, 10, 1381. [Google Scholar] [CrossRef] [PubMed]
- Yuk, H.J.; Lee, Y.S.; Ryu, H.W.; Kim, S.H.; Kim, D.S. Effects of Toona sinensis leaf extract and its chemical constituents on xanthine oxidase activity and serum uric acid levels in potassium oxonate-induced hyperuricemic rats. Molecules 2018, 23, 3254. [Google Scholar] [CrossRef] [Green Version]
- Lipkowitz, M.S. Regulation of uric acid excretion by the kidney. Curr. Rheumatol. Rep. 2012, 14, 179–188. [Google Scholar] [CrossRef]
- Ndrepepa, G. Uric acid and cardiovascular disease. Clin. Chim. Acta 2018, 484, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Chiang, Y.F.; Chen, H.Y.; Huang, Y.J.; Wang, K.L.; Hong, Y.H.; Ali, M.; Shieh, T.M.; Hsia, S.M. Anti-inflammatory and anti-hyperuricemic effects of chrysin on a high fructose corn syrup-induced hyperuricemia rat model via the amelioration of urate transporters and inhibition of NLRP3 inflammasome signaling pathway. Oxidant 2021, 10, 564. [Google Scholar]
- Maiuolo, J.; Oppedisano, F.; Gratteri, S.; Muscoli, C.; Mollace, V. Regulation of uric acid metabolism and excretion. Int. J. Cardiol. 2016, 213, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, H.; Nakayama, A.; Sakiyama, M.; Chiba, T.; Shimizu, S.; Kawamura, Y.; Nakashima, H.; Nakamura, T.; Takada, Y.; Oikawa, Y.; et al. ABCG2 dysfunction causes hyperuricemia due to both renal urate underexcretion and renal urate overload. Sci. Rep. 2014, 4, 3755. [Google Scholar] [CrossRef] [PubMed]
- Ichida, K.; Matsuo, H.; Takada, T.; Nakayama, A.; Murakami, K.; Shimizu, T.; Yamanashi, Y.; Kasuga, H.; Nakashima, H.; Nakamura, T. Decreased extra-renal urate excretion is a common cause of hyperuricemia. Nat. Commun. 2012, 3, 764. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Wang, S.; Lin, Y.; Zhao, Y.; Yang, S.; Song, J.; Xie, T.; Tian, J.; Wu, S.; Du, G. antihyperuricemic effect of mangiferin aglycon derivative J99745 by inhibiting xanthine oxidase activity and urate transporter 1 expression in mice. Acta Pharm. Sin. B 2018, 8, 306–315. [Google Scholar] [CrossRef]
- Bove, M.; Cicero, A.F.; Veronesi, M.; Borghi, C. An evidence-based review on urate-lowering treatments: Implications for optimal treatment of chronic hyperuricemia. Vasc. Health Risk Manag. 2017, 13, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, V.F.; Buiar, P.G.; Giovanella, L.H.; Severo, C.R.; Carvalho, M. Allopurinol, benzbromarone, or a combination in treating patients with gout: Analysis of a series of outpatients. Int. J. Rheumatol. 2014, 2014, 263720. [Google Scholar] [CrossRef] [Green Version]
- Hermann, W.; Lambova, S.; Muller-Ladner, U. Current Treatment Options for Osteoarthritis. Curr. Rheumatol. Rev. 2018, 14, 108–116. [Google Scholar] [CrossRef]
- Jerosch, J. Effects of glucosamine and chondroitin sulfate on cartilage metabolism in OA: Outlook on other nutrient partners especially omega-3 fatty acids. Int. J. Rheumatol. 2011, 2011, 969012. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, D.; Wang, X.; Bai, R.; Wang, C.; Gao, Y.; Anastassiades, T. N-Butyrylated hyaluronic acid ameliorates gout and hyperuricemia in animal models. Pharm. Biol. 2019, 57, 717–728. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sung, Y.-Y.; Kim, D.-S. Eggshell Membrane Ameliorates Hyperuricemia by Increasing Urate Excretion in Potassium Oxonate-Injected Rats. Nutrients 2021, 13, 3323. https://doi.org/10.3390/nu13103323
Sung Y-Y, Kim D-S. Eggshell Membrane Ameliorates Hyperuricemia by Increasing Urate Excretion in Potassium Oxonate-Injected Rats. Nutrients. 2021; 13(10):3323. https://doi.org/10.3390/nu13103323
Chicago/Turabian StyleSung, Yoon-Young, and Dong-Seon Kim. 2021. "Eggshell Membrane Ameliorates Hyperuricemia by Increasing Urate Excretion in Potassium Oxonate-Injected Rats" Nutrients 13, no. 10: 3323. https://doi.org/10.3390/nu13103323
APA StyleSung, Y.-Y., & Kim, D.-S. (2021). Eggshell Membrane Ameliorates Hyperuricemia by Increasing Urate Excretion in Potassium Oxonate-Injected Rats. Nutrients, 13(10), 3323. https://doi.org/10.3390/nu13103323