Third-Trimester Glucose Homeostasis in Healthy Women Is Differentially Associated with Human Milk Oligosaccharide Composition at 2 Months Postpartum by Secretor Phenotype


Abstract
1. Introduction
2. Materials and Methods
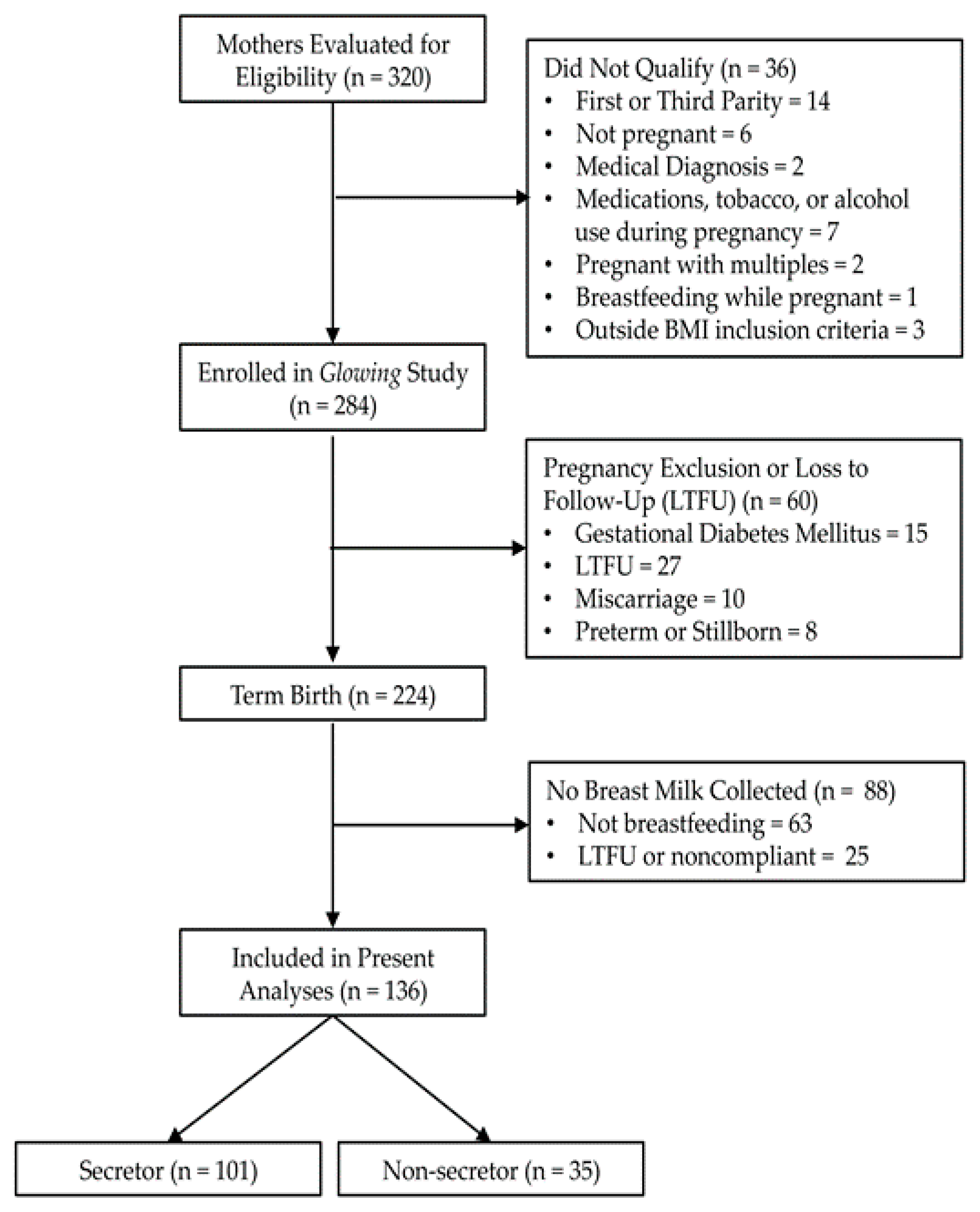
2.1. Participants and Study Design
2.2. Human Milk Collection and HMO Analysis
2.3. Maternal Body Composition and Gestational Weight Gain
2.4. Metabolic Variables
2.5. Self-Reported Outcomes
2.6. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.2. HMO Composition at 2M Postpartum in Secretors and Non-Secretors
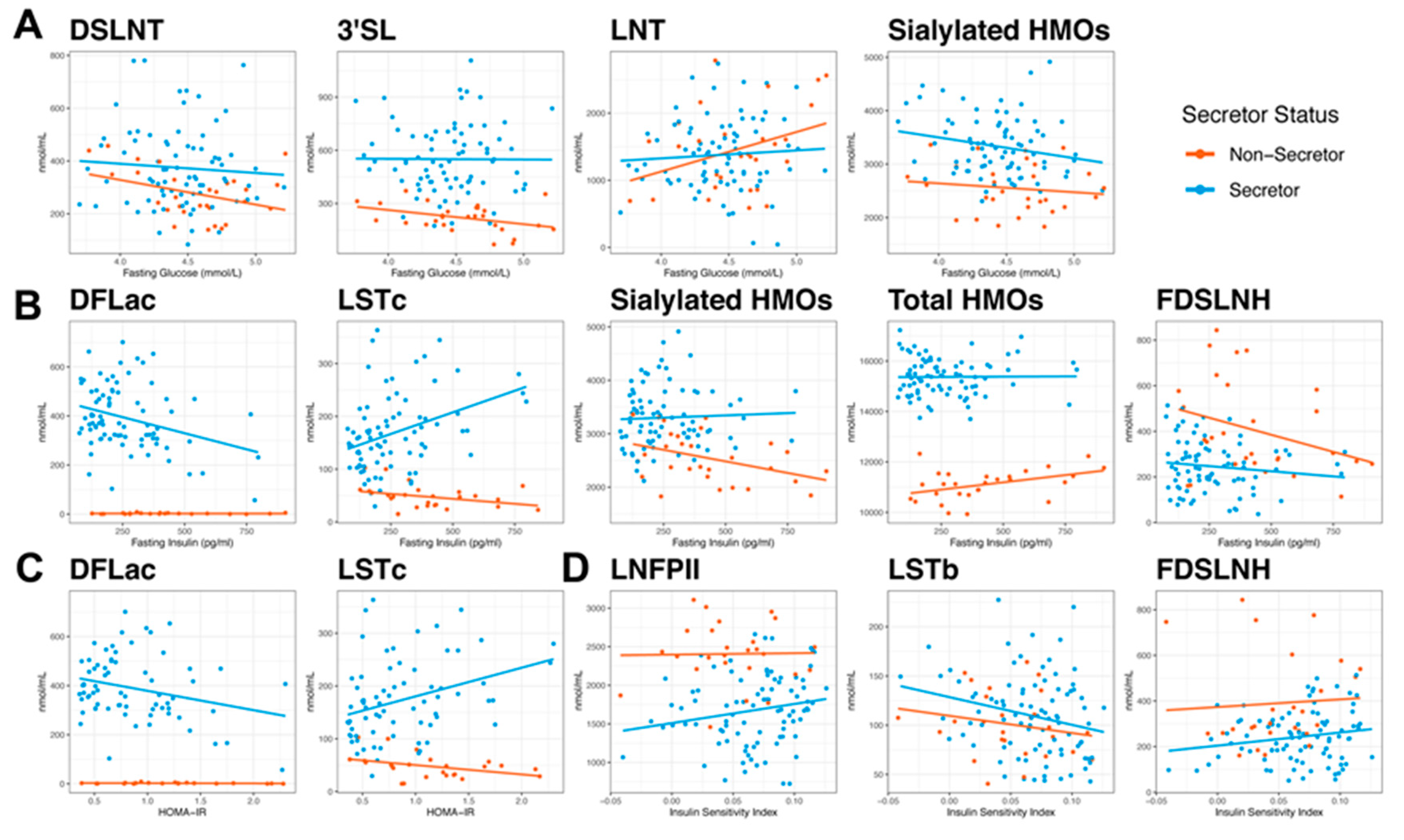
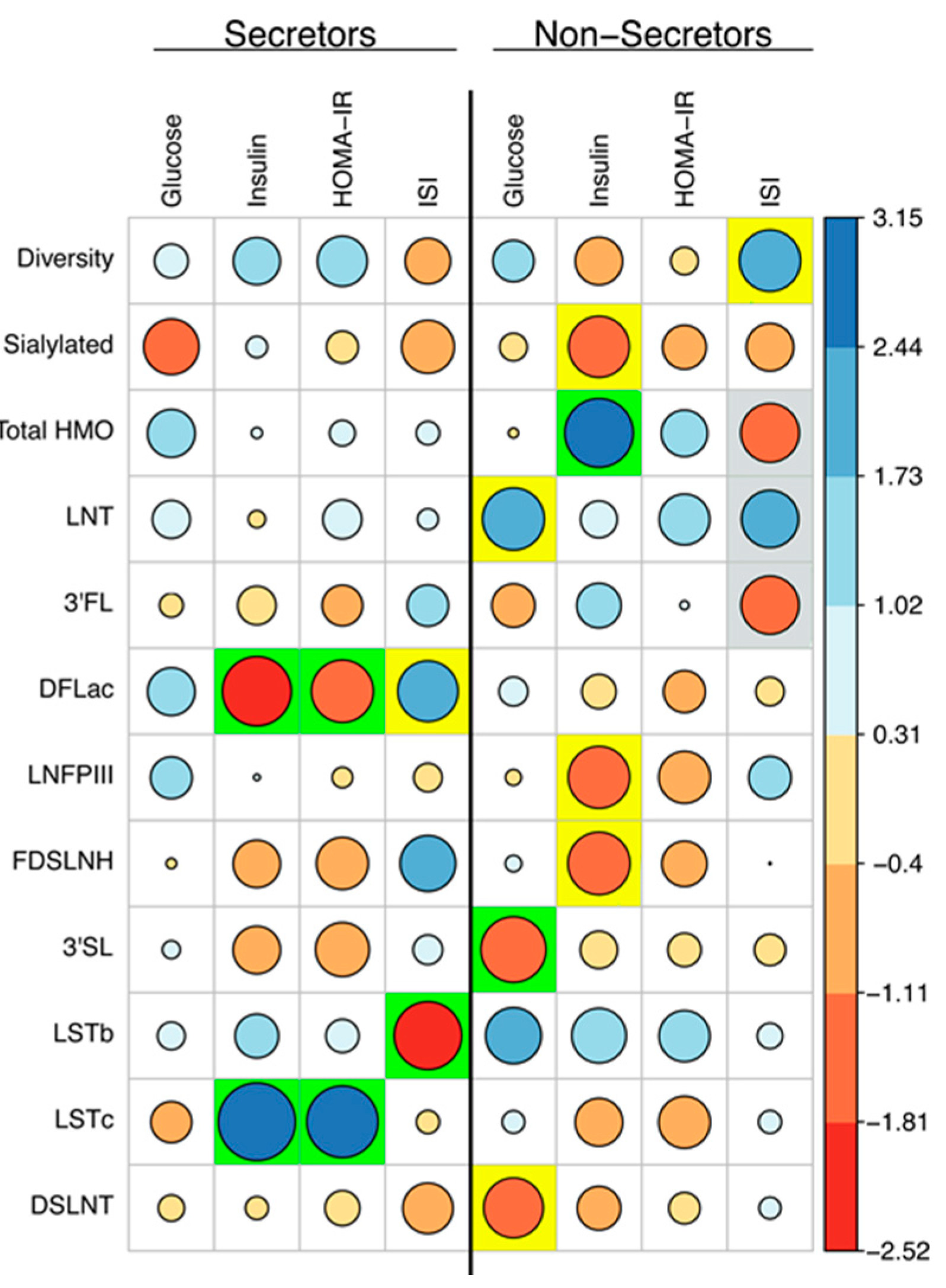
3.3. Measures of Glucose Homeostasis are Correlated with HMO Concentrations at 2M Postpartum
3.4. Maternal Characteristics can Predict HMO Concentrations at 2M Postpartum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bode, L.; Jantscher-Krenn, E. Structure-function relationships of human milk oligosaccharides. Adv. Nutr. 2012, 3, 383S–391S. [Google Scholar] [CrossRef] [PubMed]
- LoCascio, R.G.; Ninonuevo, M.R.; Freeman, S.L.; Sela, D.A.; Grimm, R.; Lebrilla, C.B.; Mills, D.A.; German, J.B. Glycoprofiling of bifidobacterial consumption of human milk oligosaccharides demonstrates strain specific, preferential consumption of small chain glycans secreted in early human lactation. J. Agric. Food Chem. 2007, 55, 8914–8919. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef]
- Sodhi, C.P.; Wipf, P.; Yamaguchi, Y.; Fulton, W.B.; Kovler, M.; Nino, D.F.; Zhou, Q.; Banfield, E.; Werts, A.D.; Ladd, M.R.; et al. The human milk oligosaccharides 2’-fucosyllactose and 6’-sialyllactose protect against the development of necrotizing enterocolitis by inhibiting toll-like receptor 4 signaling. Pediatr. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Donovan, S.M.; Comstock, S.S. Human Milk Oligosaccharides Influence Neonatal Mucosal and Systemic Immunity. Ann. Nutr. Metab. 2016, 69 (Suppl. S2), 42–51. [Google Scholar] [CrossRef] [PubMed]
- Jantscher-Krenn, E.; Aigner, J.; Reiter, B.; Kofeler, H.; Csapo, B.; Desoye, G.; Bode, L.; van Poppel, M.N.M. Evidence of human milk oligosaccharides in maternal circulation already during pregnancy: A pilot study. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E347–E357. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, J.T.; Lebrilla, C.B.; Mills, D.A.; German, J.B.; Freeman, S.L. Breast milk oligosaccharides: Structure-function relationships in the neonate. Annu. Rev. Nutr. 2014, 34, 143–169. [Google Scholar] [CrossRef]
- Bardanzellu, F.; Fanos, V.; Strigini, F.A.L.; Artini, P.G.; Peroni, D.G. Human Breast Milk: Exploring the Linking Ring among Emerging Components. Front. Pediatr. 2018, 6, 215. [Google Scholar] [CrossRef]
- McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Williams, J.E.; Foster, J.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; et al. What’s normal? Oligosaccharide concentrations and profiles in milk produced by healthy women vary geographically. Am. J. Clin. Nutr. 2017, 105, 1086–1100. [Google Scholar] [CrossRef]
- Azad, M.B.; Robertson, B.; Atakora, F.; Becker, A.B.; Subbarao, P.; Moraes, T.J.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; et al. Human Milk Oligosaccharide Concentrations Are Associated with Multiple Fixed and Modifiable Maternal Characteristics, Environmental Factors, and Feeding Practices. J. Nutr. 2018, 148, 1733–1742. [Google Scholar] [CrossRef]
- Samuel, T.M.; Binia, A.; de Castro, C.A.; Thakkar, S.K.; Billeaud, C.; Agosti, M.; Al-Jashi, I.; Costeira, M.J.; Marchini, G.; Martinez-Costa, C.; et al. Impact of maternal characteristics on human milk oligosaccharide composition over the first 4 months of lactation in a cohort of healthy European mothers. Sci. Rep. 2019, 9, 11767. [Google Scholar] [CrossRef] [PubMed]
- Sims, C.R.; Lipsmeyer, M.E.; Turner, D.E.; Andres, A. Human milk composition differs by maternal BMI in the first 9 months postpartum. Am. J. Clin. Nutr. 2020. [CrossRef]
- de la Garza Puentes, A.; Marti Alemany, A.; Chisaguano, A.M.; Montes Goyanes, R.; Castellote, A.I.; Torres-Espinola, F.J.; Garcia-Valdes, L.; Escudero-Marin, M.; Segura, M.T.; Campoy, C.; et al. The Effect of Maternal Obesity on Breast Milk Fatty Acids and Its Association with Infant Growth and Cognition-The PREOBE Follow-Up. Nutrients 2019, 11, 2154. [Google Scholar] [CrossRef]
- Young, B.E.; Levek, C.; Reynolds, R.M.; Rudolph, M.C.; MacLean, P.; Hernandez, T.L.; Friedman, J.E.; Krebs, N.F. Bioactive components in human milk are differentially associated with rates of lean and fat mass deposition in infants of mothers with normal vs. elevated BMI. Pediatr. Obes. 2018, 13, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Sonagra, A.D.; Biradar, S.M.; Dattatreya, K.; Murthy, D.S.J. Normal pregnancy- a state of insulin resistance. J. Clin. Diagn. Res. 2014, 8, CC01–CC03. [Google Scholar] [CrossRef] [PubMed]
- Catalano, P.M.; Shankar, K. Obesity and pregnancy: Mechanisms of short term and long term adverse consequences for mother and child. BMJ 2017, 356, j1. [Google Scholar] [CrossRef] [PubMed]
- Jantscher-Krenn, E.; Treichler, C.; Brandl, W.; Schonbacher, L.; Kofeler, H.; van Poppel, M.N.M. The association of human milk oligosaccharides with glucose metabolism in overweight and obese pregnant women. Am. J. Clin. Nutr. 2019, 110, 1335–1343. [Google Scholar] [CrossRef]
- Sunehag, A.L.; Louie, K.; Bier, J.L.; Tigas, S.; Haymond, M.W. Hexoneogenesis in the human breast during lactation. J. Clin. Endocrinol. Metab. 2002, 87, 297–301. [Google Scholar] [CrossRef]
- Blank, D.; Dotz, V.; Geyer, R.; Kunz, C. Human milk oligosaccharides and Lewis blood group: Individual high-throughput sample profiling to enhance conclusions from functional studies. Adv. Nutr. 2012, 3, 440S–449S. [Google Scholar] [CrossRef]
- Haney, P.M. Glucose transport in lactation. Adv. Exp. Med. Biol. 2004, 554, 253–261. [Google Scholar] [CrossRef]
- Neville, M.C.; Sawicki, V.S.; Hay, W.W., Jr. Effects of fasting, elevated plasma glucose and plasma insulin concentrations on milk secretion in women. J. Endocrinol. 1993, 139, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Neville, M.C.; Webb, P.; Ramanathan, P.; Mannino, M.P.; Pecorini, C.; Monks, J.; Anderson, S.M.; MacLean, P. The insulin receptor plays an important role in secretory differentiation in the mammary gland. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1103–E1114. [Google Scholar] [CrossRef] [PubMed]
- Menzies, K.K.; Lee, H.J.; Lefevre, C.; Ormandy, C.J.; Macmillan, K.L.; Nicholas, K.R. Insulin, a key regulator of hormone responsive milk protein synthesis during lactogenesis in murine mammary explants. Funct. Integr. Genom. 2010, 10, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Nommsen-Rivers, L.A. Does Insulin Explain the Relation between Maternal Obesity and Poor Lactation Outcomes? An Overview of the Literature. Adv. Nutr. 2016, 7, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Krukowski, R.A.; West, D.S.; DiCarlo, M.; Shankar, K.; Cleves, M.A.; Saylors, M.E.; Andres, A. Are early first trimester weights valid proxies for preconception weight? BMC Pregnancy Childbirth 2016, 16, 357. [Google Scholar] [CrossRef] [PubMed]
- Kunz, C.; Rudloff, S.; Hintelmann, A.; Pohlentz, G.; Egge, H. High-pH anion-exchange chromatography with pulsed amperometric detection and molar response factors of human milk oligosaccharides. J. Chromatogr. B Biomed. Appl. 1996, 685, 211–221. [Google Scholar] [CrossRef]
- Institute of Medicine. Institute of Medicine. Institute of Medicine and National Reserach Council Committee to Reexamine IOM Pregnancy Weight Guidelines. In Weight Gain during Pregnancy: Reexamining the Guidelines; Rasmussen, K.M., Yaktine, A.L., Eds.; National Academies Press: Washington, DC, USA, 2009. [Google Scholar] [CrossRef]
- Diaz, E.C.; Borsheim, E.; Shankar, K.; Cleves, M.A.; Andres, A. Prepregnancy Fat Free Mass and Associations to Glucose Metabolism Before and During Pregnancy. J. Clin. Endocrinol. Metab. 2019, 104, 1394–1403. [Google Scholar] [CrossRef]
- Stumvoll, M.; Mitrakou, A.; Pimenta, W.; Jenssen, T.; Yki-Jarvinen, H.; Van Haeften, T.; Renn, W.; Gerich, J. Use of the oral glucose tolerance test to assess insulin release and insulin sensitivity. Diabetes Care 2000, 23, 295–301. [Google Scholar] [CrossRef]
- Ayechu-Muruzabal, V.; van Stigt, A.H.; Mank, M.; Willemsen, L.E.M.; Stahl, B.; Garssen, J.; Van’t Land, B. Diversity of Human Milk Oligosaccharides and Effects on Early Life Immune Development. Front. Pediatr. 2018, 6, 239. [Google Scholar] [CrossRef]
- Berger, P.K.; Plows, J.F.; Jones, R.B.; Alderete, T.L.; Yonemitsu, C.; Poulsen, M.; Ryoo, J.H.; Peterson, B.S.; Bode, L.; Goran, M.I. Human milk oligosaccharide 2’-fucosyllactose links feedings at 1 month to cognitive development at 24 months in infants of normal and overweight mothers. PLoS ONE 2020, 15, e0228323. [Google Scholar] [CrossRef]
- Sprenger, N.; Binia, A.; Austin, S. Human Milk Oligosaccharides: Factors Affecting Their Composition and Their Physiological Significance. Nestle. Nutr. Inst. Workshop Ser. 2019, 90, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Yazici, D.; Sezer, H. Insulin Resistance, Obesity and Lipotoxicity. Adv. Exp. Med. Biol 2017, 960, 277–304. [Google Scholar] [CrossRef]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M.W.; Lind, M.V.; Laursen, R.P.; Yonemitsu, C.; Larnkjaer, A.; Molgaard, C.; Michaelsen, K.F.; Bode, L. Human Milk Oligosaccharide Composition Is Associated with Excessive Weight Gain During Exclusive Breastfeeding-An Explorative Study. Front. Pediatr. 2019, 7, 297. [Google Scholar] [CrossRef] [PubMed]
- Craft, K.M.; Thomas, H.C.; Townsend, S.D. Sialylated variants of lacto-N-tetraose exhibit antimicrobial activity against Group B Streptococcus. Org. Biomol. Chem. 2019, 17, 1893–1900. [Google Scholar] [CrossRef]
- Jantscher-Krenn, E.; Zherebtsov, M.; Nissan, C.; Goth, K.; Guner, Y.S.; Naidu, N.; Choudhury, B.; Grishin, A.V.; Ford, H.R.; Bode, L. The human milk oligosaccharide disialyllacto-N-tetraose prevents necrotising enterocolitis in neonatal rats. Gut 2012, 61, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Autran, C.A.; Kellman, B.P.; Kim, J.H.; Asztalos, E.; Blood, A.B.; Spence, E.C.H.; Patel, A.L.; Hou, J.; Lewis, N.E.; Bode, L. Human milk oligosaccharide composition predicts risk of necrotising enterocolitis in preterm infants. Gut 2018, 67, 1064–1070. [Google Scholar] [CrossRef]
- Charbonneau, M.R.; O’Donnell, D.; Blanton, L.V.; Totten, S.M.; Davis, J.C.; Barratt, M.J.; Cheng, J.; Guruge, J.; Talcott, M.; Bain, J.R.; et al. Sialylated Milk Oligosaccharides Promote Microbiota-Dependent Growth in Models of Infant Undernutrition. Cell 2016, 164, 859–871. [Google Scholar] [CrossRef]
- Cowardin, C.A.; Ahern, P.P.; Kung, V.L.; Hibberd, M.C.; Cheng, J.; Guruge, J.L.; Sundaresan, V.; Head, R.D.; Barile, D.; Mills, D.A.; et al. Mechanisms by which sialylated milk oligosaccharides impact bone biology in a gnotobiotic mouse model of infant undernutrition. Proc. Natl. Acad. Sci. USA 2019, 116, 11988–11996. [Google Scholar] [CrossRef]
- Alderete, T.L.; Autran, C.; Brekke, B.E.; Knight, R.; Bode, L.; Goran, M.I.; Fields, D.A. Associations between human milk oligosaccharides and infant body composition in the first 6 mo of life. Am. J. Clin. Nutr. 2015, 102, 1381–1388. [Google Scholar] [CrossRef]
- D’Adamo, P.J.; Kelly, G.S. Metabolic and immunologic consequences of ABH secretor and Lewis subtype status. Altern. Med. Rev. 2001, 6, 390–405. [Google Scholar] [PubMed]
- Smyth, D.J.; Cooper, J.D.; Howson, J.M.; Clarke, P.; Downes, K.; Mistry, T.; Stevens, H.; Walker, N.M.; Todd, J.A. FUT2 nonsecretor status links type 1 diabetes susceptibility and resistance to infection. Diabetes 2011, 60, 3081–3084. [Google Scholar] [CrossRef] [PubMed]
- Ihara, K.; Fukano, C.; Ayabe, T.; Fukami, M.; Ogata, T.; Kawamura, T.; Urakami, T.; Kikuchi, N.; Yokota, I.; Takemoto, K.; et al. FUT2 non-secretor status is associated with Type 1 diabetes susceptibility in Japanese children. Diabet. Med. 2017, 34, 586–589. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Non-Secretor (n = 35) | Secretor (n = 101) | Total (n = 136) | p-Value | |
|---|---|---|---|---|
| Maternal Age (years) | 0.382 1 | |||
| Mean (SD) | 30.182 (2.884) | 30.743 (3.375) | 30.599 (3.255) | |
| Maternal Race (n (%)) | 0.335 2 | |||
| Caucasian | 29 (82.9%) | 90 (89.1%) | 119 (87.5%) | |
| Non-Caucasian | 6 (17.1%) | 11 (10.9%) | 17 (12.5%) | |
| Maternal BMI (kg/m2) | 0.004 1 | |||
| Mean (SD) | 27.194 (4.048) | 24.890 (3.945) | 25.483 (4.084) | |
| Maternal BMI ≥ 25.0 kg/m2 | 0.011 2 | |||
| n (%) | 24 (68.6%) | 44 (43.6%) | 68 (50.0%) | |
| Gestational Weight Gain (kg) | 0.114 1 | |||
| Mean (SD) | 11.050 (4.127) | 12.248 (3.721) | 11.955 (3.852) | |
| IOM GWG Recommendation (n (%)) | 0.819 2 | |||
| Inadequate | 8 (22.9%) | 25 (25.3%) | 33 (24.6%) | |
| Appropriate | 14 (40.0%) | 43 (43.4%) | 57 (42.5%) | |
| Excessive | 13 (37.1%) | 31 (31.3%) | 44 (32.8%) | |
| Gestational Age (weeks) | 0.996 1 | |||
| Mean (SD) | 39.298 (0.813) | 39.297 (0.904) | 39.297 (0.879) | |
| Delivery Mode | 0.864 2 | |||
| C-Section | 11 (31.4%) | 33 (33.0%) | 44 (32.6%) | |
| Vaginal | 24 (68.6%) | 67 (67.0%) | 91 (67.4%) | |
| Infant Sex | 0.040 2 | |||
| Female | 10 (30.6%) | 49 (48.5%) | 60 (43.4%) | |
| Male | 25 (71.4%) | 52 (51.5%) | 79 (56.6%) |
| Non-Secretor (n = 35) | Secretor (n = 101) | Total (n = 136) | p-Value | |
|---|---|---|---|---|
| Fasting Glucose (mmol/L) | 0.096 1 | |||
| Mean (SD) | 4.553 (0.366) | 4.435 (0.307) | 4.466 (0.326) | |
| Fasting Insulin (pmol/L) | <0.001 1 | |||
| Mean (SD) | 62.500 (27.056) | 42.719 (21.889) | 47.521 (24.628) | |
| HOMA-IR | 0.005 1 | |||
| Mean (SD) | 1.179 (0.533) | 0.871 (0.446) | 0.953 (0.488) | |
| Insulin Sensitivity Index (μmol × kg−1 × min−1 × pM−1) | 0.021 1 | |||
| Mean (SD) | 0.049 (0.036) | 0.067 (0.035) | 0.063 (0.036) |
| Non-Secretor (n = 35) | Secretor (n = 101) | Total (n = 136) | p-Value 1 | |
|---|---|---|---|---|
| Fucosylated, Median (Q1, Q3) | ||||
| 2′FL | 6.40 (2.90, 11.50) | 1580.40 (1143.25, 2156.02) | 1264.10 (19.95, 1914.35) | <0.001 |
| DFLac | 1.90 (0.85, 3.05) | 358.50 (288.85, 469.75) | 320.90 (147.40, 431.45) | <0.001 |
| LNFP I | 142.80 (114.15, 167.10) | 1199.70 (746.20, 1854.05) | 859.70 (197.20, 1513.12) | <0.001 |
| 3FL | 4455.30 (3724.00, 5071.45) | 3320.10 (2459.20, 4384.90) | 3689.15 (2631.80, 4639.85) | <0.001 |
| LNFP II | 2406.40 (2197.00, 2710.20) | 1679.40 (1341.45, 1986.12) | 1814.00 (1458.40, 2249.90) | <0.001 |
| LNFP III | 28.30 (17.95, 44.15) | 49.10 (36.40, 68.85) | 42.90 (30.60, 65.20) | <0.001 |
| DFLNT | 456.20 (290.60, 601.70) | 1971.00 (1778.40, 2238.40) | 1842.80 (816.93, 2119.85) | <0.001 |
| FLNH | 135.55 (86.75, 189.23) | 171.20 (124.60, 248.53) | 167.55 (115.45, 235.67) | 0.0501 |
| DFLNH | 25.25 (19.80, 37.58) | 79.00 (50.55, 125.80) | 56.80 (31.90, 99.40) | <0.001 |
| Sialylated, Median (Q1, Q3) | ||||
| 3′SL | 223.90 (176.55, 275.95) | 531.60 (408.00, 678.90) | 428.35 (257.40, 608.32) | <0.001 |
| 6′SL | 583.65 (436.90, 706.85) | 770.90 (604.70, 997.42) | 712.85 (574.25, 926.15) | <0.001 |
| LSTb | 95.45 (75.72, 127.78) | 109.50 (80.95, 136.35) | 107.50 (80.00, 136.10) | 0.2361 |
| LSTc | 47.60 (31.50, 60.30) | 166.75 (126.03, 215.65) | 131.50 (73.95, 195.90) | <0.001 |
| DSLNT | 245.50 (214.60, 324.15) | 353.70 (273.85, 476.07) | 323.50 (245.50, 431.80) | <0.001 |
| DSLNH | 79.40 (57.65, 108.30) | 136.40 (88.90, 182.30) | 120.85 (78.72, 168.97) | <0.001 |
| Sialylated & Fucosylated, Median (Q1, Q3) | ||||
| FDSLNH | 304.50 (260.25, 558.40) | 232.60 (152.70, 303.10) | 256.75 (172.92, 346.80) | <0.001 |
| Non-fucosylated, neutral, Median (Q1, Q3) | ||||
| LNT | 1389.80 (876.35, 1647.30) | 1396.70 (1061.00, 1776.18) | 1396.30 (1016.25, 1738.75) | 0.8131 |
| LNnT | 92.80 (61.65, 157.10) | 101.90 (73.95, 145.40) | 101.10 (71.55, 146.12) | 0.5361 |
| LNH | 127.05 (75.15, 208.08) | 173.65 (120.05, 258.90) | 169.20 (104.90, 250.60) | 0.0521 |
| Diversity and Evenness 2, Median (Q1, Q3) | ||||
| Diversity (log) | 1.47 (1.33, 1.62) | 2.04 (1.89, 2.15) | 1.97 (1.65, 2.13) | <0.001 |
| Evenness (log) | −1.48 (−1.61, −1.32) | −0.91 (−1.05, −0.79) | −0.98 (−1.30, −0.81) | <0.001 |
| Total Concentrations Median (Q1, Q3) | ||||
| HMO-bound Sia | 2483.40 (2195.40, 2790.30) | 3201.60 (2908.05, 3730.15) | 3036.60 (2658.15, 3458.70) | <0.001 |
| HMO-bound Fuc | 8847.40 (8043.10, 9430.40) | 13718.30 (12827.50, 14602.70) | 13087.55 (10944.05, 14180.55) | <0.001 |
| Overall Total | 11180.15 (10737.27, 11533.83) | 15407.05 (14860.42, 15806.28) | 15008.35 (12668.05, 15657.68) | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saben, J.L.; Abraham, A.; Bode, L.; Sims, C.R.; Andres, A. Third-Trimester Glucose Homeostasis in Healthy Women Is Differentially Associated with Human Milk Oligosaccharide Composition at 2 Months Postpartum by Secretor Phenotype. Nutrients 2020, 12, 2209. https://doi.org/10.3390/nu12082209
Saben JL, Abraham A, Bode L, Sims CR, Andres A. Third-Trimester Glucose Homeostasis in Healthy Women Is Differentially Associated with Human Milk Oligosaccharide Composition at 2 Months Postpartum by Secretor Phenotype. Nutrients. 2020; 12(8):2209. https://doi.org/10.3390/nu12082209
Chicago/Turabian StyleSaben, Jessica L., Ann Abraham, Lars Bode, Clark R. Sims, and Aline Andres. 2020. "Third-Trimester Glucose Homeostasis in Healthy Women Is Differentially Associated with Human Milk Oligosaccharide Composition at 2 Months Postpartum by Secretor Phenotype" Nutrients 12, no. 8: 2209. https://doi.org/10.3390/nu12082209
APA StyleSaben, J. L., Abraham, A., Bode, L., Sims, C. R., & Andres, A. (2020). Third-Trimester Glucose Homeostasis in Healthy Women Is Differentially Associated with Human Milk Oligosaccharide Composition at 2 Months Postpartum by Secretor Phenotype. Nutrients, 12(8), 2209. https://doi.org/10.3390/nu12082209