The Powdered Root of Eurycoma longifolia Jack Improves Beta-Cell Number and Pancreatic Islet Performance through PDX1 Induction and Shows Antihyperglycemic Activity in db/db Mice

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and E. longifolia (EL) Powdered Root Preparation
2.2. Animal Husbandry
2.3. Animal Experiments
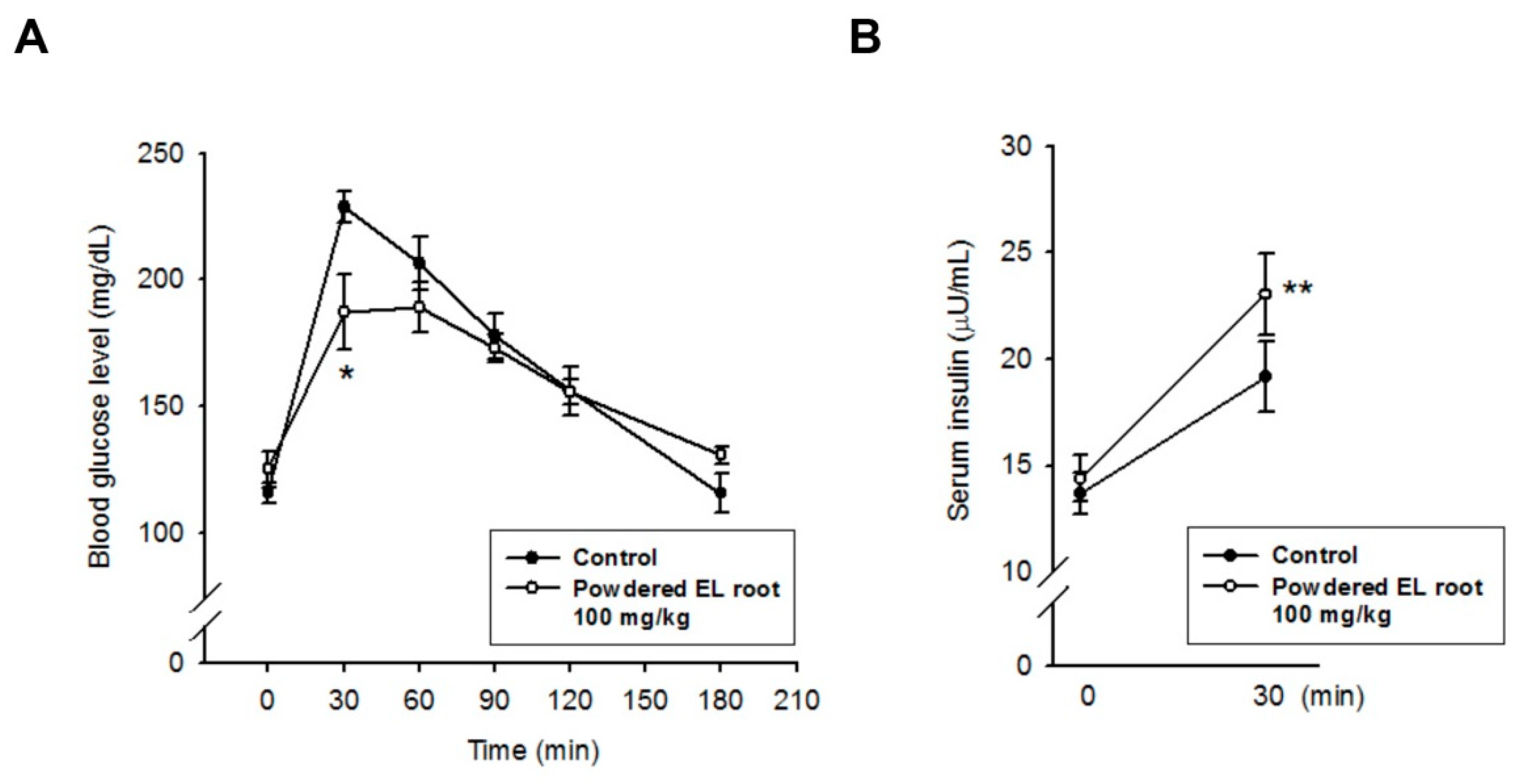
2.4. Oral Glucose Tolerance Test (OGTT)
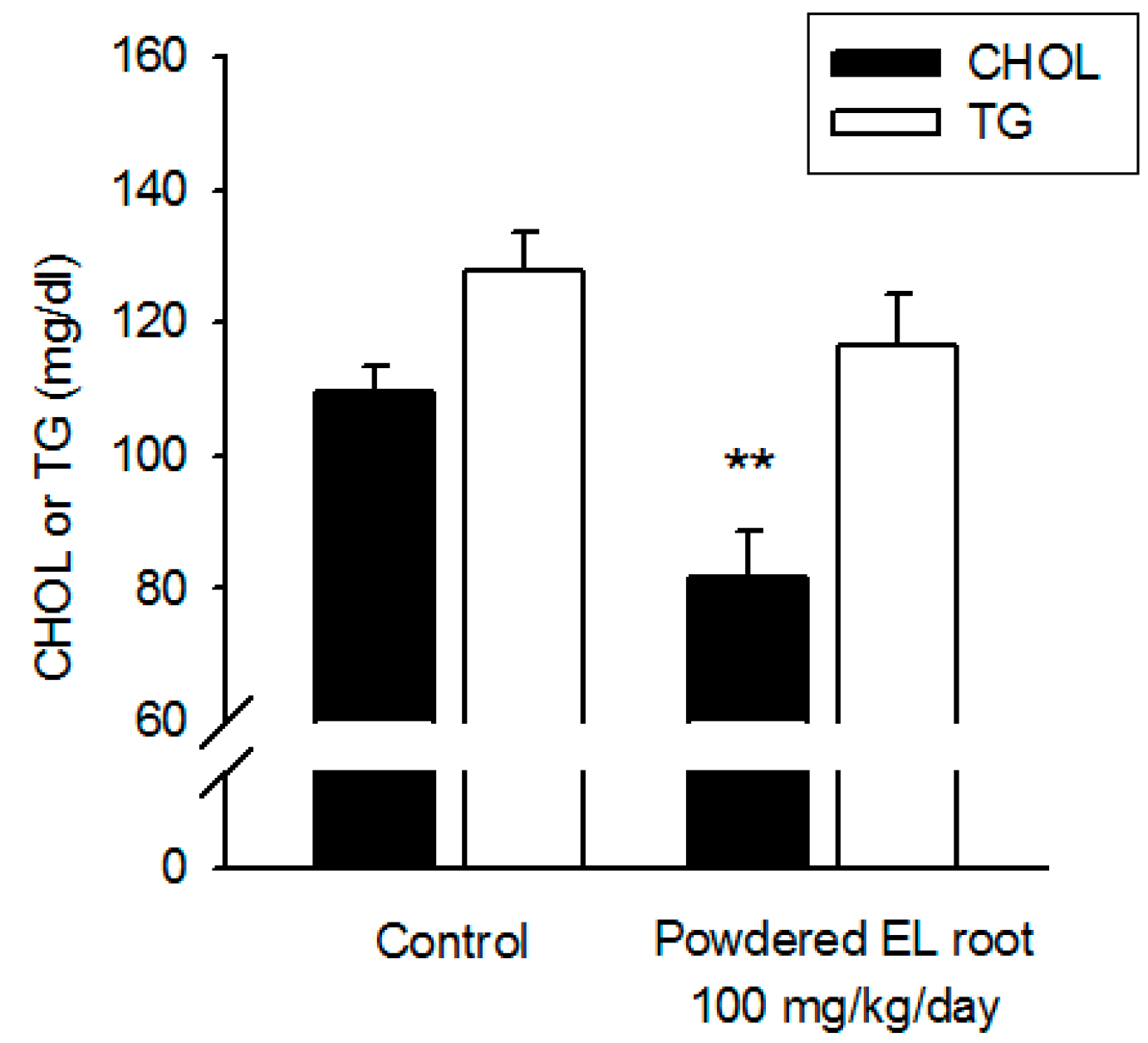
2.5. Serum Biochemistry
2.6. Insulin Quantification and Beta-Cell Function Evaluation
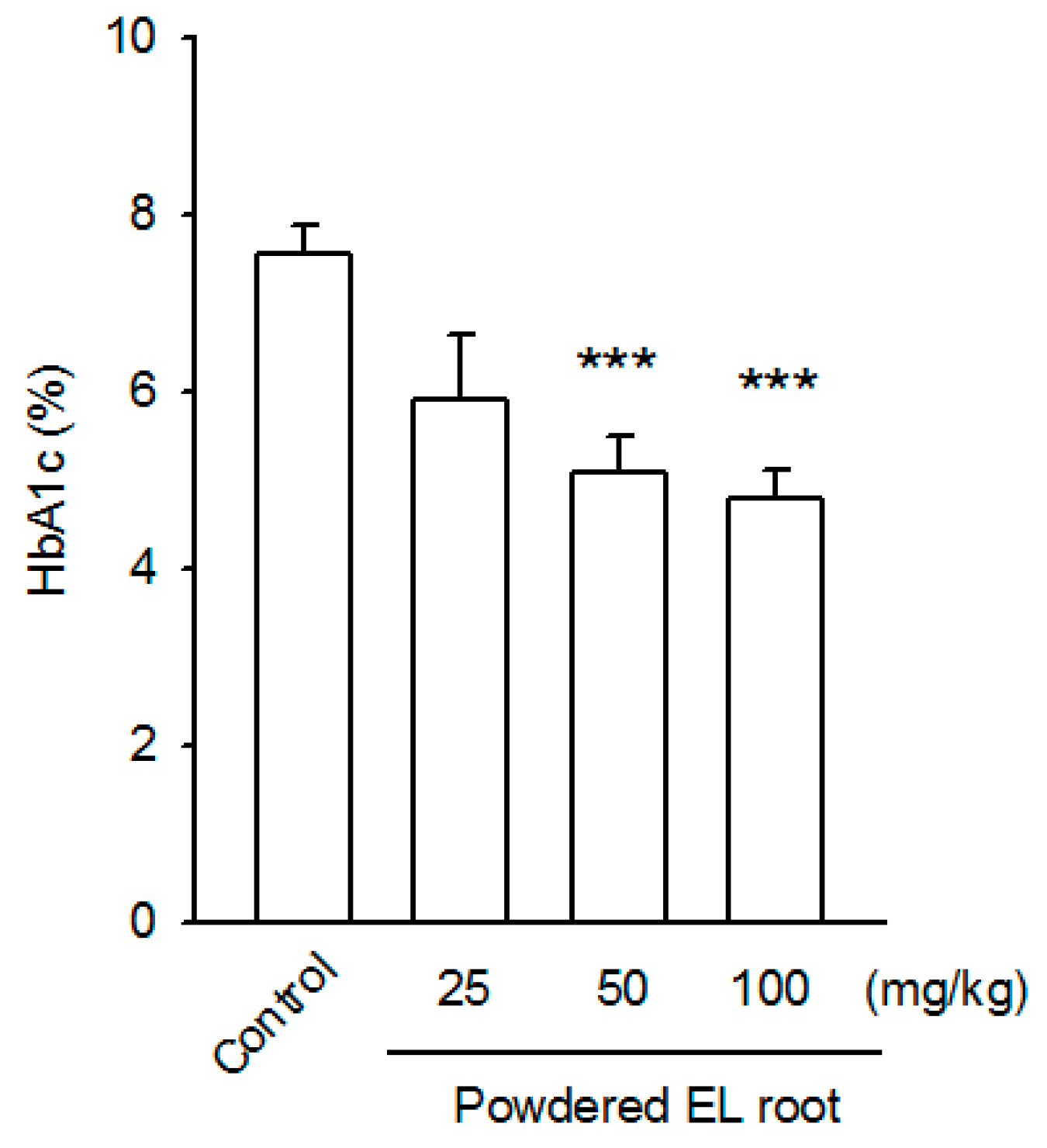
2.7. Determination of Glycated Hemoglobin (HbA1c)
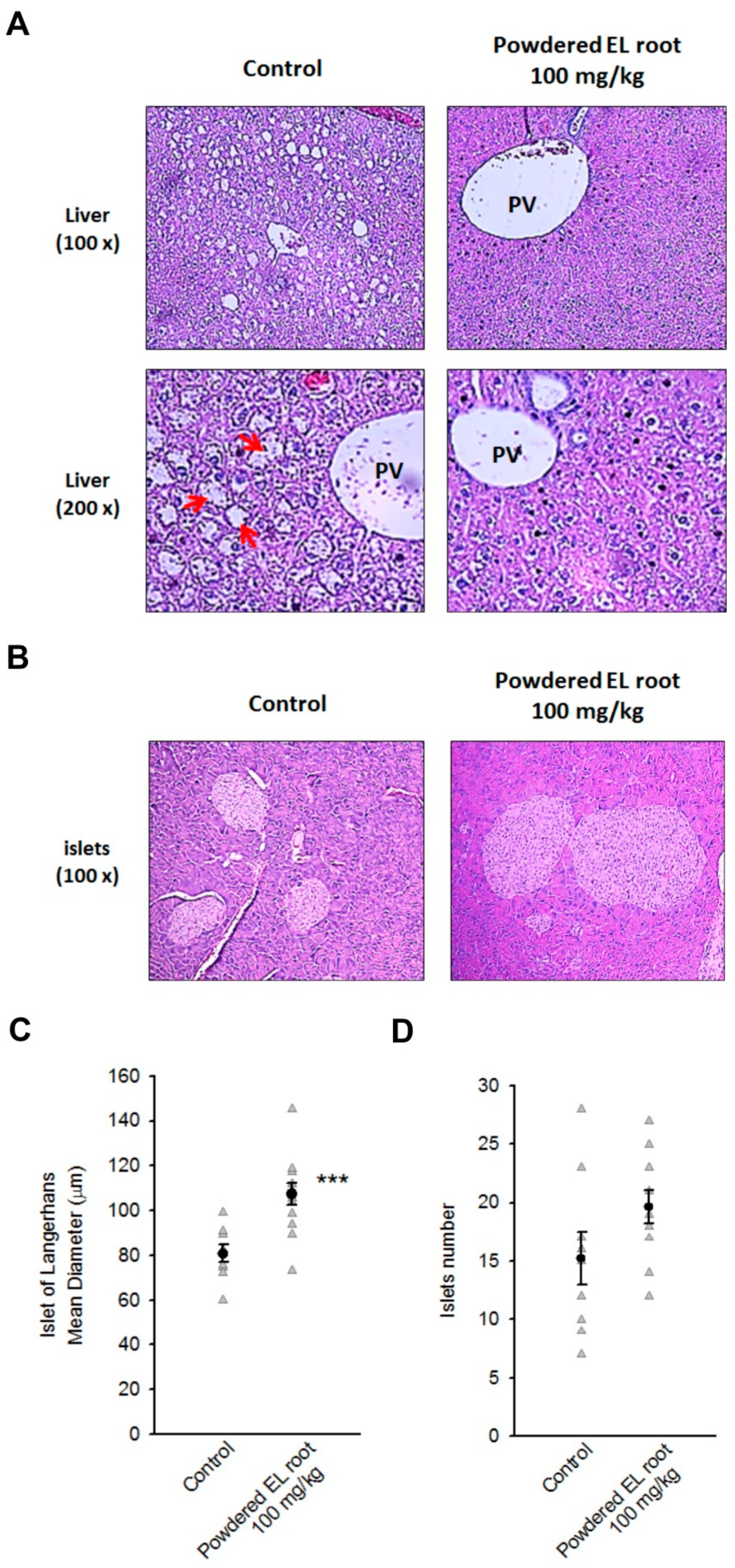
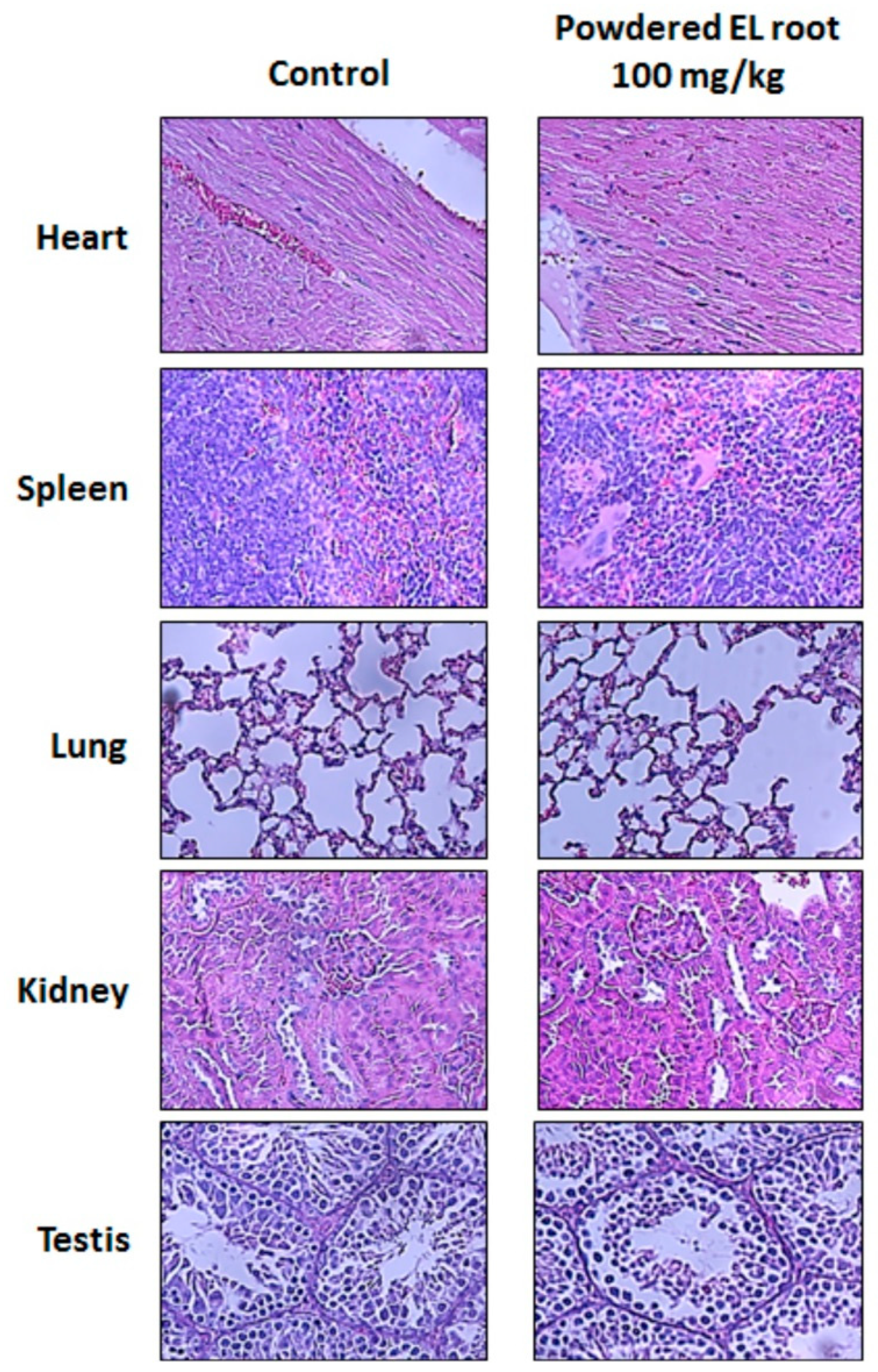
2.8. Histopathological Examination and Immunohistochemical (IHC) Staining
2.9. Statistical Analyses
3. Results
3.1. E. longifolia (EL) Reduced Fasting Blood Glucose Levels in Diabetic db/db Mice
3.2. Effects of Powdered EL Root on Oral Glucose-Loaded Diabetic db/db Mice
3.3. Powdered EL Root Had Lipid-Lowering Effects in Diabetic db/db Mice
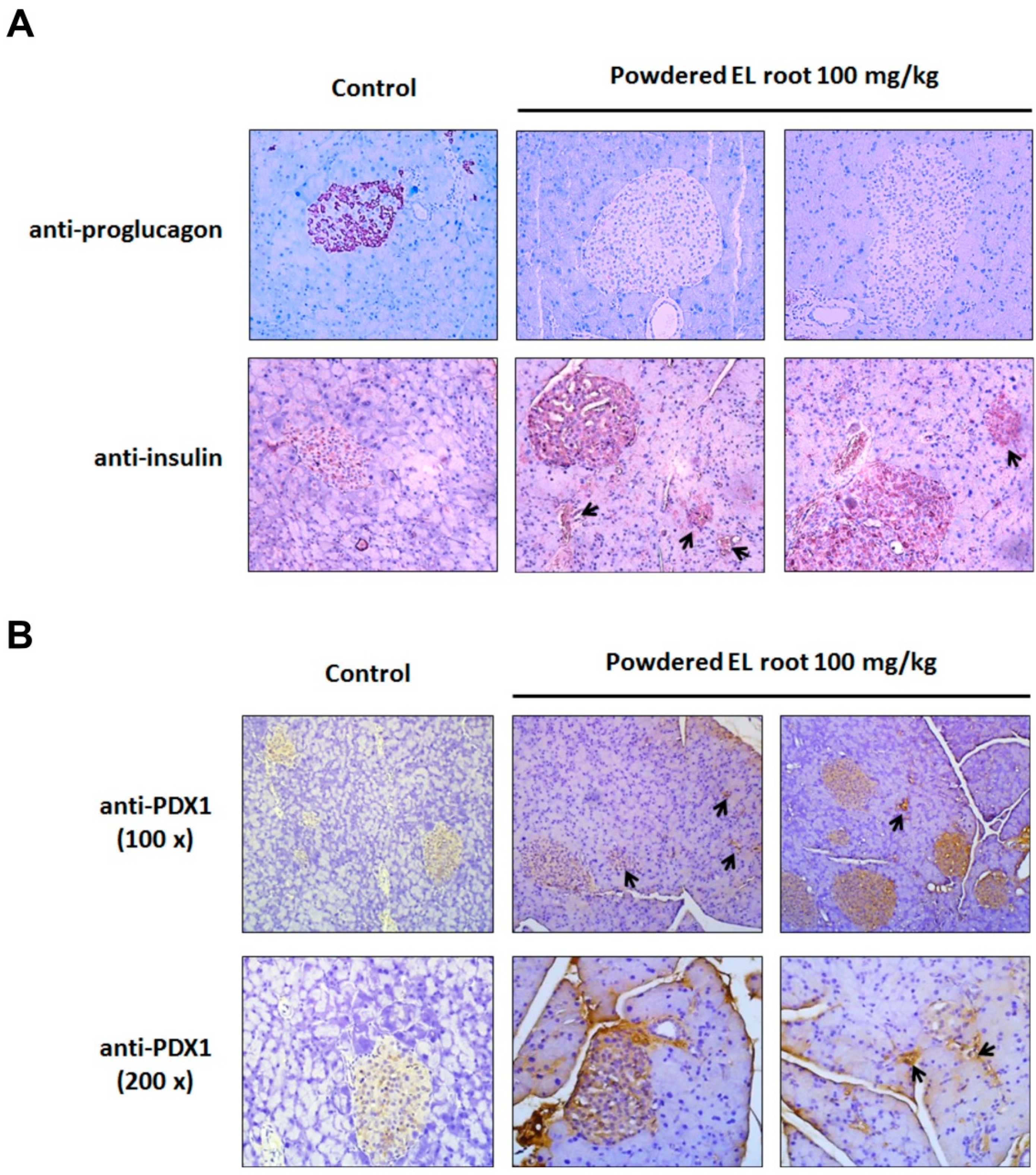
3.4. Powdered EL Root Reduced Glycogen Accumulation in Hepatocytes but Improved Pancreatic Islet Volume in db/db Mice
3.5. Powdered EL Root Increased PDX1 Expression and Beta-Cell Number Resulting in an Improvement in Insulin Secretion
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EL | Eurycoma longifolia |
| DM | diabetes mellitus |
| IDDM | insulin-dependent diabetes mellitus |
| NIDDM | non-insulin-dependent diabetes mellitus |
| PDX1 | pancreatic and duodenal homeobox 1 |
| HbA1c | hemoglobin A1c |
| OGTT | Oral glucose tolerance test |
| NOAEL | no observed adverse effect level |
| ADI | acceptable daily intake |
| HOMA-IR | homeostasis model assessment-insulin resistance |
| HOMA-β | homeostasis model assessment-beta-cell function |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Powdered EL Root (mg/kg) | Blood Glucose Level (mg/dL) | |||||
|---|---|---|---|---|---|---|
| Diabetic db/db Mice | Normoglycemic C57BL/6J Mice | |||||
| Week 0 | Week 1 | Week 2 | Week 0 | Week 1 | Week 2 | |
| 0 | 361.0 ± 49.0 | 366.0 ± 46.5 | 336.3 ± 19.9 | 128.6 ± 7.1 | 153.4 ± 6.7 | 118.2 ± 7.2 |
| 200 | 369.3 ± 20.0 | 184.8 ± 20.4 * | 150.0 ± 22.8 * | 156.6 ± 2.2 | 137.4 ± 5.4 | 115.8 ± 5.7 |

References
- Kaiser, A.B.; Zhang, N.; Pluijm, W.V.D. Global Prevalence of Type 2 Diabetes over the Next Ten Years (2018–2028). Diabetes 2018, (Suppl. 1). [Google Scholar] [CrossRef]
- Tao, Z.; Shi, A.; Zhao, J. Epidemiological perspectives of diabetes. Cell. Biochem. Biophys. 2015, 73, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Ata, A.; V Anil Kumar, N.; Sharopov, F.; Ramírez-Alarcón, K.; Ruiz-Ortega, A.; Abdulmajid Ayatollahi, S.; Tsouh Fokou, P.V.; Kobarfard, F.; Amiruddin Zakaria, Z.; et al. Antidiabetic potential of medicinal plants and their active components. Biomolecules 2019, 9, 551. [Google Scholar] [CrossRef]
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14. [Google Scholar] [CrossRef]
- Tillin, T.; Hughes, A.D.; Godsland, I.F.; Whincup, P.; Forouhi, N.G.; Welsh, P.; Sattar, N.; McKeigue, P.M.; Chaturvedi, N. Insulin resistance and truncal obesity as important determinants of the greater incidence of diabetes in Indian Asians and African Caribbeans compared with Europeans: The Southall And Brent REvisited (SABRE) cohort. Diabetes Care. 2013, 36, 383–393. [Google Scholar] [CrossRef]
- World Health Organization. Available online: http://www.who.int/mediacentre/factsheets/fs312/en/ (accessed on 31 May 2016).
- Schwingshackl, L.; Hoffmann, G.; Lampousi, A.M.; Knüppel, S.; Iqbal, K.; Schwedhelm, C.; Bechthold, A.; Schlesinger, S.; Boeing, H. Food groups and risk of type 2 diabetes mellitus: A systematic review and meta-analysis of prospective studies. Eur. J. Epidemiol. 2017, 32, 363–375. [Google Scholar] [CrossRef]
- Neeland, I.J.; Turer, A.T.; Ayers, C.R.; Powell-Wiley, T.M.; Vega, G.L.; Farzaneh-Far, R.; Grundy, S.M.; Khera, A.; McGuire, D.K.; de Lemos, J.A. Dysfunctional adiposity and the risk of prediabetes and type 2 diabetes in obese adults. JAMA 2012, 308, 1150–1159. [Google Scholar] [CrossRef]
- Hamer, M.; Bostock, S.; Hackett, R.; Steptoe, A. Objectively assessed sedentary time and type 2 diabetes mellitus: A case–control study. Diabetologia 2013, 56, 2761–2762. [Google Scholar] [CrossRef]
- Reis, J.P.; Loria, C.M.; Sorlie, P.D.; Park, Y.; Hollenbeck, A.; Schatzkin, A. Lifestyle factors and risk for new-onset diabetes: A population-based cohort study. Ann. Intern. Med. 2011, 155, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.U.; Choe, K.; Yoo, H.H. Review on a traditional herbal medicine, Eurycoma longifolia Jack (Tongkat Ali): Its traditional uses, chemistry, evidence-based pharmacology and toxicology. Molecules 2016, 21, 331. [Google Scholar] [CrossRef]
- Bhat, R.; Karim, A. Tongkat Ali (Eurycoma longifolia Jack): A review on its ethnobotany and pharmacological importance. Fitoterapia 2010, 81, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Low, B.S.; Choi, S.B.; Abdul Wahab, H.; Das, P.K.; Chan, K.L. Eurycomanone, the major quassinoid in Eurycoma longifolia root extract increases spermatogenesis by inhibiting the activity of phosphodiesterase and aromatase in steroidogenesis. J. Ethnopharmacol. 2013, 149, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Henkel, R.R.; Wang, R.; Bassett, S.H.; Chen, T.; Liu, N.; Zhu, Y.; Tambi, M.I. Tongkat Ali as a potential herbal supplement for physically active male and female seniors–A pilot study. Phytother. Res. 2014, 28, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Ezzat, S.M.; Ezzat, M.I.; Okba, M.M.; Hassan, S.M.; Alkorashy, A.I.; Karar, M.M.; Ahmed, S.H.; Mohamed, S.O. Brain cortical and hippocampal dopamine: A new mechanistic approach for Eurycoma longifolia well-known aphrodisiac activity and its chemical characterization. Evid. Based Complement. Alternat. Med. 2019, 2019, 7543460. [Google Scholar] [CrossRef]
- Tee, B.H.; Hoe, S.Z.; Cheah, S.H.; Lam, S.K. First report of Eurycoma longifolia Jack root extract causing relaxation of aortic rings in rats. Biomed. Res. Int. 2016, 2016, 1361508. [Google Scholar] [CrossRef]
- He, P.; Dong, Z.; Wang, Q.; Zhan, Q.P.; Zhang, M.M.; Wu, H. Structural characterization and immunomodulatory activity of a polysaccharide from Eurycoma longifolia. J. Nat. Prod. 2019, 82, 169–176. [Google Scholar] [CrossRef]
- Thu, H.E.; Hussain, Z.; Mohamed, I.N.; Shuid, A.N. Eurycoma longifolia, a promising suppressor of RANKL-induced differentiation and activation of osteoclasts: An in vitro mechanistic evaluation. J. Ayurveda Integr. Med. 2019, 10, 102–110. [Google Scholar] [CrossRef]
- Thu, H.E.; Hussain, Z.; Mohamed, I.N.; Shuid, A.N. Eurycoma longifolia, A potential phytomedicine for the treatment of cancer: Evidence of p53-mediated apoptosis in cancerous cells. Curr. Drug Targets. 2018, 19, 1109–1126. [Google Scholar] [CrossRef]
- Talbott, S.M.; Talbott, J.A.; George, A.; Pugh, M. Effect of Tongkat Ali on stress hormones and psychological mood state in moderately stressed subjects. J. Int. Soc. Sports Nutr. 2013, 10, 28. [Google Scholar] [CrossRef]
- Hajjouli, S.; Chateauvieux, S.; Teiten, M.H.; Orlikova, B.; Schumacher, M.; Dicato, M.; Choo, C.Y.; Diederich, M. Eurycomanone and eurycomanol from Eurycoma longifolia Jack as regulators of signaling pathways involved in proliferation, cell death and inflammation. Molecules 2014, 19, 14649–14666. [Google Scholar] [CrossRef]
- Wong, P.F.; Cheong, W.F.; Shu, M.H.; The, C.H.; Chan, K.L.; AbuBakar, S. Eurycomanone suppresses expression of lung cancer cell tumor markers, prohibitin, annexin 1 and endoplasmic reticulum protein 28. Phytomedicine 2012, 19, 138–144. [Google Scholar] [CrossRef]
- Tung, N.H.; Uto, T.; Hai, N.T.; Li, G.; Shoyama, Y. Quassinoids from the root of Eurycoma longifolia and their antiproliferative activity on human cancer cell lines. Pharmacogn. Mag. 2017, 13, 459–462. [Google Scholar] [CrossRef]
- Li, C.H.; Liao, J.W.; Liao, P.L.; Huang, W.K.; Tse, L.S.; Lin, C.H.; Kang, J.J.; Cheng, Y.W. Evaluation of acute 13-week subchronic toxicity and genotoxicity of the powdered root of Tongkat Ali (Eurycoma longifolia Jack). Evid. Based Complement. Alternat. Med. 2013, 2013, 102987. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia. 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Kobayashi, K.; Forte, T.M.; Taniguchi, S.; Ishida, B.Y.; Oka, K.; Chan, L. The db/db mouse, a model for diabetic dyslipidemia: Molecular characterization and effects of Western diet feeding. Metabolism 2000, 49, 22–31. [Google Scholar] [CrossRef]
- Sears, B.; Perry, M. The role of fatty acids in insulin resistance. Lipids Health Dis. 2015, 14, 121. [Google Scholar] [CrossRef]
- Fayyad, A.M.; Khan, A.A.; Abdallah, S.H.; Alomran, S.S.; Bajou, K.; Khattak, M.N.K. Rosiglitazone enhances browning adipocytes in association with MAPK and PI3-K pathways during the differentiation of telomerase-transformed mesenchymal stromal cells into adipocytes. Int. J. Mol. Sci. 2019, 20, 1618. [Google Scholar] [CrossRef]
- Biserni, M.; Mesnage, R.; Ferro, R.; Wozniak, E.; Xenakis, T.; Mein, C.A.; Antoniou, M.N. Quizalofop-p-ethyl induces adipogenesis in 3T3-L1 adipocytes. Toxicol. Sci. 2019, 170, 452–461. [Google Scholar] [CrossRef]
- Al-Hussaini, A.A.; Sulaiman, N.M.; Alzahrani, M.D.; Alenizi, A.S.; Khan, M. Prevalence of hepatopathy in type 1 diabetic children. BMC Pediatr. 2012, 12, 160. [Google Scholar] [CrossRef]
- Sherigar, J.M.; Castro, J.; Yin, Y.M.; Guss, D.; Mohanty, S.R. Glycogenic hepatopathy: A narrative review. World J. Hepatol. 2018, 10, 172–185. [Google Scholar] [CrossRef]
- Lahrita, L.; Kato, E.; Kawabata, J. Uncovering potential of Indonesian medicinal plants on glucose uptake enhancement and lipid suppression in 3T3-L1 adipocytes. J. Ethnopharmacol. 2015, 168, 229–236. [Google Scholar] [CrossRef]
- Butler, A.E.; Janson, J.; Bonner-Weir, S.; Ritzel, R.; Rizza, R.A.; Butler, P.C. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes 2003, 52, 102–110. [Google Scholar] [CrossRef]
- Rahier, J.; Guiot, Y.; Goebbels, R.M.; Sempoux, C.; Henquin, J.C. Pancreatic beta-cell mass in European subjects with type 2 diabetes. Diabetes Obes. Metab. 2008, 10 (Suppl. 4), 32–42. [Google Scholar] [CrossRef]
- Tran, T.V.; Malainer, C.; Schwaiger, S.; Atanasov, A.G.; Heiss, E.H.; Dirsch, V.M.; Stuppner, H. NF-κB inhibitors from Eurycoma longifolia. J. Nat. Prod. 2014, 77, 483–488. [Google Scholar] [CrossRef]
- Han, Y.M.; Woo, S.U.; Choi, M.S.; Park, Y.N.; Kim, S.H.; Yim, H.; Yoo, H.H. Antiinflammatory and analgesic effects of Eurycoma longifolia extracts. Arch. Pharm. Res. 2016, 39, 421–428. [Google Scholar] [CrossRef]
- Ruan, J.; Li, Z.; Zhang, Y.; Chen, Y.; Liu, M.; Han, L.; Zhang, Y.; Wang, T. Bioactive constituents from the roots of Eurycoma longifolia. Molecules 2019, 24, 3157. [Google Scholar] [CrossRef]
- Burke, S.J.; Batdorf, H.M.; Burk, D.H.; Noland, R.C.; Eder, A.E.; Boulos, M.S.; Karlstad, M.D.; Collier, J.J. db/db mice exhibit features of human type 2 diabetes that are not present in weight-matched C57BL/6J mice fed a western diet. J. Diabetes Res. 2017, 2017, 8503754. [Google Scholar] [CrossRef]
- Weir, G.C.; Bonner-Weir, S. Islet β cell mass in diabetes and how it relates to function, birth, and death. Ann. N. Y. Acad. Sci. 2013, 1281, 92–105. [Google Scholar] [CrossRef]
- Brüning, J.C.; Winnay, J.; Bonner-Weir, S.; Taylor, S.I.; Accili, D.; Kahn, C.R. Development of a novel polygenic model of NIDDM in mice heterozygous for IR and IRS-1 null alleles. Cell 1997, 88, 561–572. [Google Scholar] [CrossRef]
- Gapp, D.A.; Leiter, E.H.; Coleman, D.L.; Schwizer, R.W. Temporal changes in pancreatic islet composition in C57BL/6J-db/db (diabetes) mice. Diabetologia 1983, 25, 439–443. [Google Scholar] [CrossRef]
- Zhou, Q.; Brown, J.; Kanarek, A.; Rajagopal, J.; Melton, D.A. In vivo reprogramming of adult pancreatic exocrine cells to beta-cells. Nature 2008, 455, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liu, Q.; Zhou, Z.; Ikeda, Y. PDX1, Neurogenin-3, and MAFA: Critical transcription regulators for beta cell development and regeneration. Stem Cell Res. Ther. 2017, 8, 240. [Google Scholar] [CrossRef] [PubMed]
- Holland, A.M.; Hale, M.A.; Kagami, H.; Hammer, R.E.; MacDonald, R.J. Experimental control of pancreatic development and maintenance. Proc. Natl. Acad. Sci. USA 2002, 99, 12236–12241. [Google Scholar] [CrossRef]
- Holland, A.M.; Góñez, L.J.; Naselli, G.; Macdonald, R.J.; Harrison, L.C. Conditional expression demonstrates the role of the homeodomain transcription factor Pdx1 in maintenance and regeneration of beta-cells in the adult pancreas. Diabetes 2005, 54, 2586–2595. [Google Scholar] [CrossRef] [PubMed]
- Bru-Tari, E.; Cobo-Vuilleumier, N.; Alonso-Magdalena, P.; Dos Santos, R.S.; Marroqui, L.; Nadal, A.; Gauthier, B.R.; Quesada, I. Pancreatic alpha-cell mass in the early-onset and advanced stage of a mouse model of experimental autoimmune diabetes. Sci. Rep. 2019, 9, 9515. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, Y.; Xu, N.; Zhou, W.; Yang, L.; Chen, R.; Yang, R.; Sun, J.; Chen, H. A new way for beta cell neogenesis: Transdifferentiation from alpha cells induced by glucagon-like peptide 1. J. Diabetes Res. 2019, 2019, 2583047. [Google Scholar] [CrossRef]
- Xiao, X.; Guo, P.; Shiota, C.; Zhang, T.; Coudriet, G.M.; Fischbach, S.; Prasadan, K.; Fusco, J.; Ramachandran, S.; Witkowski, P.; et al. Endogenous reprogramming of alpha cells into beta cells, induced by viral gene therapy, reverses autoimmune diabetes. Cell Stem Cell. 2018, 22, 78–90.e4. [Google Scholar] [CrossRef]
- Husen, R.; Pihie, A.H.; Nallappan, M. Screening for antihyperglycaemic activity in several local herbs of Malaysia. J. Ethnopharmacol. 2004, 95, 205–208. [Google Scholar] [CrossRef]
- Shuid, A.; Siang, L.; Chin, T.; Muhammad, N.; Mohamed, N.; Soelaiman, I. Acute and Subacute Toxicity Studies of Eurycoma longifolia in Male Rats. Int. J. Pharm. 2011, 7, 641–646. [Google Scholar] [CrossRef]
- Choudhary, Y.K.; Bommu, P.; Ming, Y.K.; Zulkawi, N.B. Acute, sub-acute, and subchronic 90-days toxicity of Eurycoma longifolia aqueous extract (Physta) in wistar rats. Int. J. Pharm. Pharm. Sci. 2012, 4, 232–238. [Google Scholar]







| Powdered EL Root (mg/kg) | Mean Body Weight (g) | ||||
|---|---|---|---|---|---|
| Week 0 | Week 1 | Week 2 | Week 4 | Week 8 | |
| 0 | 50.65 ± 1.82 | 52.06 ± 1.57 | 52.80 ± 1.40 | 54.4 ± 1.43 | 55.04 ± 1.61 |
| 25 | 54.89 ± 1.27 | 55.07 ± 1.24 | 55.13 ± 1.23 | 54.49 ± 1.38 | 56.16 ± 1.21 |
| 50 | 52.91 ± 2.53 | 53.75 ± 2.17 | 53.71 ± 1.76 | 54.15 ± 1.80 | 53.16 ± 1.87 |
| 100 | 51.54 ± 1.86 | 51.84 ± 1.80 | 51.15 ± 2.05 | 50.58 ± 2.47 | 50.17 ± 3.05 |
| Powdered EL Root (mg/kg) | Blood Glucose Level (mg/dL) | ||||
|---|---|---|---|---|---|
| Week 0 | Week 1 | Week 2 | Week 4 | Week 8 | |
| 0 | 346.2 ± 11.9 | 367.0 ± 33.2 | 344.67 ± 26.4 | 398.1 ± 17.8 | 373.6 ± 32.0 |
| 25 | 306.0 ± 33.2 | 305.2 ± 26.3 | 285.6 ± 35.5 | 282.3 ± 35.7 | |
| 50 | 300.3 ± 38.7 | 226.6 ± 36.0 * | 205.0 ± 26.3 * | 193.6 ± 29.7 * | |
| 100 | 228.8 ± 27.7 * | 150.8 ± 18.5 * | 143.0 ± 16.4 * | 134.6 ± 18.7 * | |
| Powdered EL Root (mg/kg) | Blood Glucose Level (mg/dL) | |||||
|---|---|---|---|---|---|---|
| 0 min | 30 min | 60 min | 90 min | 120 min | 180 min | |
| 0 | 373.9 ± 31.0 | 750.4 ± 43.6 | 680.5 ± 33.8 | 555.7 ± 32.9 | 503.6 ± 33.6 | 398.2 ± 26.5 |
| 25 | 337.9 ± 46.8 | 582.5 ± 37.8 | 584.2 ± 49.2 | 542.8 ± 46.1 | 484.5 ± 36.9 | 395.4 ± 42.9 |
| 50 | 269.5 ± 37.2 | 523.0 ± 65.7 | 496.2 ± 71.6 | 446.8 ± 78.8 | 427.8 ± 86.6 | 362.3 ± 57.6 |
| 100 | 174.9 ± 26.5 | 396.4 ± 23.7 | 322.3 ± 30.2 | 279.8 ± 24.7 | 252.9 ± 25.5 | 181.4 ± 20.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, C.-H.; Fang, T.-C.; Liao, P.-L.; Liao, J.-W.; Chan, Y.-J.; Cheng, Y.-W.; Li, C.-H. The Powdered Root of Eurycoma longifolia Jack Improves Beta-Cell Number and Pancreatic Islet Performance through PDX1 Induction and Shows Antihyperglycemic Activity in db/db Mice. Nutrients 2020, 12, 2111. https://doi.org/10.3390/nu12072111
Tsai C-H, Fang T-C, Liao P-L, Liao J-W, Chan Y-J, Cheng Y-W, Li C-H. The Powdered Root of Eurycoma longifolia Jack Improves Beta-Cell Number and Pancreatic Islet Performance through PDX1 Induction and Shows Antihyperglycemic Activity in db/db Mice. Nutrients. 2020; 12(7):2111. https://doi.org/10.3390/nu12072111
Chicago/Turabian StyleTsai, Chi-Hao, Te-Chao Fang, Po-Lin Liao, Jiunn-Wang Liao, Yen-Ju Chan, Yu-Wen Cheng, and Ching-Hao Li. 2020. "The Powdered Root of Eurycoma longifolia Jack Improves Beta-Cell Number and Pancreatic Islet Performance through PDX1 Induction and Shows Antihyperglycemic Activity in db/db Mice" Nutrients 12, no. 7: 2111. https://doi.org/10.3390/nu12072111
APA StyleTsai, C.-H., Fang, T.-C., Liao, P.-L., Liao, J.-W., Chan, Y.-J., Cheng, Y.-W., & Li, C.-H. (2020). The Powdered Root of Eurycoma longifolia Jack Improves Beta-Cell Number and Pancreatic Islet Performance through PDX1 Induction and Shows Antihyperglycemic Activity in db/db Mice. Nutrients, 12(7), 2111. https://doi.org/10.3390/nu12072111