Zinc Oxide Exerts Anti-Inflammatory Properties on Human Placental Cells

 , , , , ,
, , , , ,  and
and 
Abstract
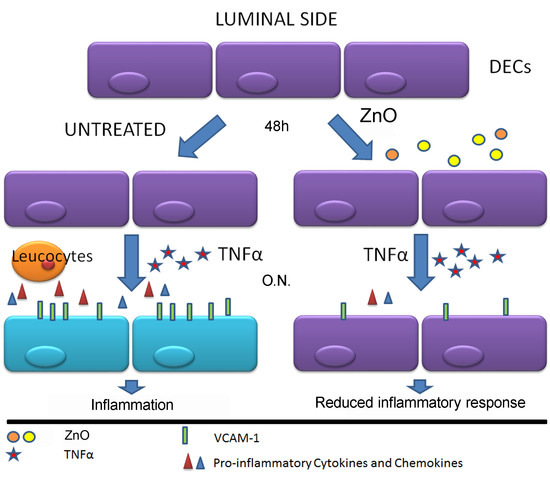
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Isolation and Culture
2.3. Immunofluorescence
2.4. Enzyme-Linked Immunosorbent Assay (ELISA) for VCAM-1 or ICAM-1 on Whole Cells
2.5. Evaluation of mRNA Expression of Pro-Inflammatory Cytokines
2.6. Evaluation of IL-8 Secretion by ELISA
2.7. Statistical Analysis
3. Results
3.1. Definition of the Optimal Concentration of ZnO for In Vitro Analysis
3.2. ZnO Exerted Anti-Inflammatory Effect Reducing the Expression of VCAM-1 by TNF-α Stimulated DECs
3.3. Analysis of the ICAM-1 Expression by First Trimester Trophoblast Cells Stimulated with TNF-α
3.4. ZnO Modulated the Pro-Inflammatory Response of DECs, Reducing Their Expression of Inflammatory Cytokines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Redline, R.W. Placental inflammation. Semin Neonatol. 2004, 9, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.C.V. Bounds KR, Mitchell BM. Regulation of the Anti-Inflammatory Cytokines Interleukin-4 and Interleukin-10 during Pregnancy. Front. Immunol. 2014, 27, 253. [Google Scholar]
- Zev Williams. Inducing Tolerance to Pregnancy. N. Eng. J. Med. 2012, 387, 1159–1181. [Google Scholar]
- Xu, Y.; Plazyo, O.; Romero, R.; Hassan, S.S.; Gomez-Lopez, N. Isolation of Leukocytes from the Human Maternal-fetal Interface. J. Vis. Exp. 2015. [Google Scholar] [CrossRef]
- Masat, E.; Gasparini, C.; Agostinis, C.; Bossi, F.; Radillo, O.; De Seta, F.; Tamassia, N.; Cassatella, M.A.; Bulla, R. RelB activation in anti-inflammatory decidual endothelial cells: A master plan to avoid pregnancy failure? Sci. Rep. 2015, 5, 14847. [Google Scholar] [CrossRef]
- Bulla, R.; Bossi, F.; Agostinis, C.; Radillo, O.; Colombo, F.; De Seta, F.; Tedesco, F. Complement production by trophoblast cells at the feto-maternal interface. J. Reprod. Immunol. 2009, 82, 119–125. [Google Scholar] [CrossRef]
- Gaffuri, B.; Vigano, P.; Nozza, A.; Gornati, G.; Di Blasio, A.M.; Vignali, M. Expression of intercellular adhesion molecule-1 messenger ribonucleic acid and protein in human term placental cells and its modulation by pro-inflammatory cytokines (interleukin-1beta and tumor necrosis factor alpha). Biol. Reprod. 1998, 58, 1003–1008. [Google Scholar] [CrossRef]
- Lundqvist. Reported dietary intake in early pregnant compared to non-pregnant women–A cross-sectional study. BMC Pregnancy Childbirth 2014, 14, 373. [Google Scholar]
- Marangoni, F.; Cetin, I.; Verduci, E.; Canzone, G.; Giovannini, M.; Scollo, P.; Corsello, G.; Poli, A. Maternal Diet and Nutrient Requirements in Pregnancy and Breastfeeding. An Italian Consensus Document. Nutrients 2016, 8, 629. [Google Scholar] [CrossRef]
- Nordic Nutrition Recommendations. Available online: https://norden.diva-portal.org/smash/get/diva2:704251/FULLTEXT01.pdf (accessed on 10 March 2014).
- Terrin, G.; Berni Canani, R.; Di Chiara, M.; Pietravalle, A.; Aleandri, V.; Conte, F.; De Curtis, M. Zinc in Early Life: A Key Element in the Fetus and Preterm Neonate. Nutrients 2015, 7, 10427–10446. [Google Scholar] [CrossRef] [PubMed]
- Chaffee, B.W.; King, J.C. Effect of zinc supplementation on pregnancy and infant outcomes: A systematic review. Paediatr. Perinat Epidemiol. 2012, 26 (Suppl. 1), 118–137. [Google Scholar] [CrossRef]
- Gernand, A.D.; Schulze, K.J.; Stewart, C.P.; West, K.P., Jr.; Christian, P. Micronutrient deficiencies in pregnancy worldwide: Health effects and prevention. Nat. Rev. Endocrinol. 2016, 12, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Adamo, A.M.; Oteiza, P.I. Zinc deficiency and neurodevelopment: The case of neurons. Biofactors 2010, 36, 117–124. [Google Scholar] [CrossRef]
- Ha, H.T.T.; Leal-Ortiz, S.; Lalwani, K.; Kiyonaka, S.; Hamachi, I.; Mysore, S.P.; Montgomery, J.M.; Garner, C.C.; Huguenard, J.R.; Kim, S.A. Shank and Zinc Mediate an AMPA Receptor Subunit Switch in Developing Neurons. Front. Mol. Neurosci. 2018, 11, 405. [Google Scholar] [CrossRef]
- Huang, L.; Drake, V.J.; Ho, E. Zinc. Adv. Nutr. 2015, 6, 224–226. [Google Scholar] [CrossRef]
- Osendarp, S.J.; West, C.E.; Black, R.E.; Maternal Zinc Supplementation Study Group. The need for maternal zinc supplementation in developing countries: An unresolved issue. J. Nutr. 2003, 133, 817S–827S. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc in human health: Effect of zinc on immune cells. Mol. Med. 2008, 14, 353–357. [Google Scholar] [CrossRef]
- Swanson, C.A.; King, J.C. Zinc and pregnancy outcome. Am. J. Clin. Nutr. 1987, 46, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Cousins, R.J. Absorption, transport, and hepatic metabolism of copper and zinc: Special reference to metallothionein and ceruloplasmin. Physiol. Rev. 1985, 65, 238–309. [Google Scholar] [CrossRef]
- Turnlund, J.R.; King, J.C.; Keyes, W.R.; Gong, B.; Michel, M.C. A stable isotope study of zinc absorption in young men: Effects of phytate and alpha-cellulose. Am. J. Clin. Nutr. 1984, 40, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Bulla, R.; Agostinis, C.; Bossi, F.; Rizzi, L.; Debeus, A.; Tripodo, C.; Radillo, O.; De Seta, F.; Ghebrehiwet, B.; Tedesco, F. Decidual endothelial cells express surface-bound C1q as a molecular bridge between endovascular trophoblast and decidual endothelium. Mol. Immunol. 2008, 45, 2629–2640. [Google Scholar] [CrossRef] [PubMed]
- Edwards, H.M., 3rd; Baker, D.H. Bioavailability of zinc in several sources of zinc oxide, zinc sulfate, and zinc metal. J. Anim. Sci. 1999, 77, 2730–2735. [Google Scholar] [CrossRef]
- Maares, M.; Haase, H. A Guide to Human Zinc Absorption: General Overview and Recent Advances of In Vitro Intestinal Models. Nutrients 2020, 12, 762. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Bussolino, F.; Dejana, E. Cytokine regulation of endothelial cell function. FASEB J. 1992, 6, 2591–2599. [Google Scholar] [CrossRef]
- Osborn, L.; Hession, C.; Tizard, R.; Vassallo, C.; Luhowskyj, S.; Chi-Rosso, G.; Lobb, R. Direct expression cloning of vascular cell adhesion molecule 1, a cytokine-induced endothelial protein that binds to lymphocytes. Cell 1989, 59, 1203–1211. [Google Scholar] [CrossRef]
- Redman, C.W.; Sacks, G.P.; Sargent, I.L. Preeclampsia: An excessive maternal inflammatory response to pregnancy. Am. J. Obstet. Gynecol. 1999, 180, 499–506. [Google Scholar] [CrossRef]
- Romero, R.; Dey, S.K.; Fisher, S.J. Preterm labor: One syndrome, many causes. Science 2014, 345, 760–765. [Google Scholar] [CrossRef]
- Gathiram, P.; Moodley, J. Pre-eclampsia: Its pathogenesis and pathophysiolgy. Cardiovasc J. Afr. 2016, 27, 71–78. [Google Scholar] [CrossRef]
- Takacs, P.; Damjanovich, P.; Sipos, A.G.; Kozma, B. The effect of oral zinc supplementation on cervicovaginal lavage fluid zinc level. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 248, 106–109. [Google Scholar] [CrossRef]
- Wessels, I.; Maywald, M.; Rink, L. Zinc as a Gatekeeper of Immune Function. Nutrients 2017, 9, 1286. [Google Scholar] [CrossRef]
- Gammoh, N.Z.; Rink, L. Zinc in Infection and Inflammation. Nutrients 2017, 9, 624. [Google Scholar] [CrossRef]
- Neish, A.S.; Williams, A.J.; Palmer, H.J.; Whitley, M.Z.; Collins, T. Functional analysis of the human vascular cell adhesion molecule 1 promoter. J. Exp. Med. 1992, 176, 1583–1593. [Google Scholar] [CrossRef] [PubMed]
- Agostinis, C.; Masat, E.; Bossi, F.; Ricci, G.; Menegazzi, R.; Lombardelli, L.; Zito, G.; Mangogna, A.; Degan, M.; Gattei, V.; et al. Transcriptomics and Immunological Analyses Reveal a Pro-Angiogenic and Anti-Inflammatory Phenotype for Decidual Endothelial Cells. Int. J. Mol. Sci. 2019, 20, 1604. [Google Scholar] [CrossRef]
- Bonaventura, P.; Benedetti, G.; Albarede, F.; Miossec, P. Zinc and its role in immunity and inflammation. Autoimmun. Rev. 2015, 14, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Mariani, E.; Cattini, L.; Neri, S.; Malavolta, M.; Mocchegiani, E.; Ravaglia, G.; Facchini, A. Simultaneous evaluation of circulating chemokine and cytokine profiles in elderly subjects by multiplex technology: Relationship with zinc status. Biogerontology 2006, 7, 449–459. [Google Scholar] [CrossRef]
- Foster, M.; Samman, S. Zinc and regulation of inflammatory cytokines: Implications for cardiometabolic disease. Nutrients 2012, 4, 676–694. [Google Scholar] [CrossRef]
- Vasto, S.; Mocchegiani, E.; Candore, G.; Listi, F.; Colonna-Romano, G.; Lio, D.; Malavolta, M.; Giacconi, R.; Cipriano, C.; Caruso, C. Inflammation, genes and zinc in ageing and age-related diseases. Biogerontology 2006, 7, 315–327. [Google Scholar] [CrossRef]
- Kinlaw, W.B.; Levine, A.S.; Morley, J.E.; Silvis, S.E.; McClain, C.J. Abnormal zinc metabolism in type II diabetes mellitus. Am. J. Med. 1983, 75, 273–277. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc: An antioxidant and anti-inflammatory agent: Role of zinc in degenerative disorders of aging. J. Trace Elem. Med. Biol. 2014. [Google Scholar] [CrossRef]
- Ota, E.; Mori, R.; Middleton, P.; Tobe-Gai, R.; Mahomed, K.; Miyazaki, C.; Bhutta, Z.A. Zinc supplementation for improving pregnancy and infant outcome. Cochrane Database Syst. Rev. 2015. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Tm | Sense | Sequence | Accession Number |
|---|---|---|---|---|
| 18S | 60 | Forward | ATCCCTGAAAAGTTCCAGCA | NM_022551.2 |
| Reverse | CCCTCTTGGTGAGGTCAATG | |||
| IL-8 | 60 | Forward | AGGTGCAGTAGTTTTGCCAAGGA | NM_000584.3 |
| Reverse | TTTCTGTGTTGGCGCAGTGT | |||
| TNF-α | 66 | Forward | GGCCCAGGCAGTCAGATCAT | NM_000594.3 |
| Reverse | GGGGCTCTTGATGGCAGAGA | |||
| IL-6 | 60 | Forward | GTACATCCTCGACGGCATC | NM_000600.3 |
| Reverse | CCAGGCAAGTCTCCTCATTG | |||
| MCP-1 | 60 | Forward | ATCAATGCCCCAGTACC | NM_002982 |
| Reverse | AGTCTTCGGTAGTTTGGG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balduit, A.; Mangogna, A.; Agostinis, C.; Zito, G.; Romano, F.; Ricci, G.; Bulla, R. Zinc Oxide Exerts Anti-Inflammatory Properties on Human Placental Cells. Nutrients 2020, 12, 1822. https://doi.org/10.3390/nu12061822
Balduit A, Mangogna A, Agostinis C, Zito G, Romano F, Ricci G, Bulla R. Zinc Oxide Exerts Anti-Inflammatory Properties on Human Placental Cells. Nutrients. 2020; 12(6):1822. https://doi.org/10.3390/nu12061822
Chicago/Turabian StyleBalduit, Andrea, Alessandro Mangogna, Chiara Agostinis, Gabriella Zito, Federico Romano, Giuseppe Ricci, and Roberta Bulla. 2020. "Zinc Oxide Exerts Anti-Inflammatory Properties on Human Placental Cells" Nutrients 12, no. 6: 1822. https://doi.org/10.3390/nu12061822
APA StyleBalduit, A., Mangogna, A., Agostinis, C., Zito, G., Romano, F., Ricci, G., & Bulla, R. (2020). Zinc Oxide Exerts Anti-Inflammatory Properties on Human Placental Cells. Nutrients, 12(6), 1822. https://doi.org/10.3390/nu12061822