Calcium Oxalate Nephrolithiasis and Gut Microbiota: Not just a Gut-Kidney Axis. A Nutritional Perspective

 ,
,
Abstract
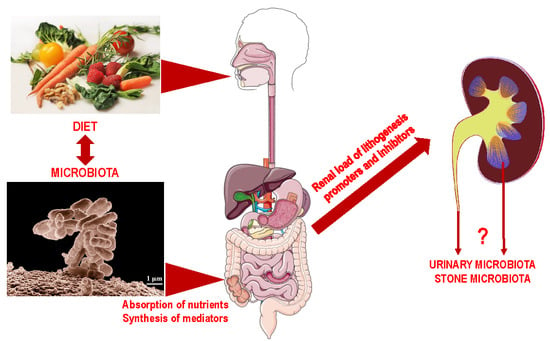
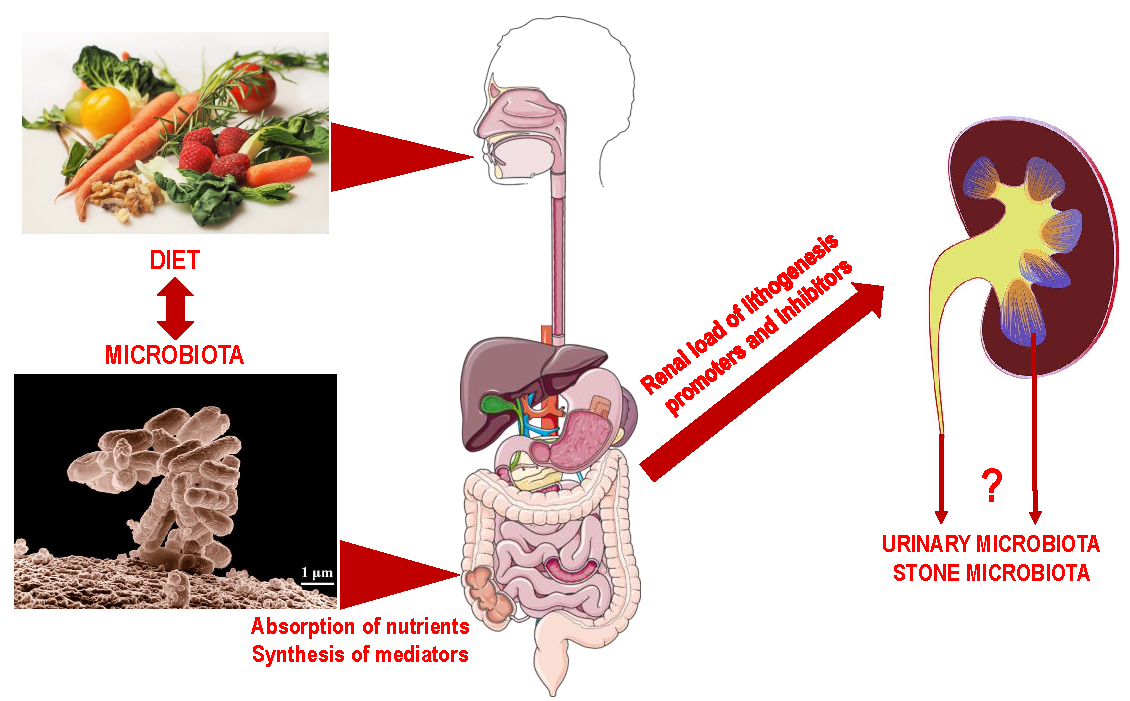
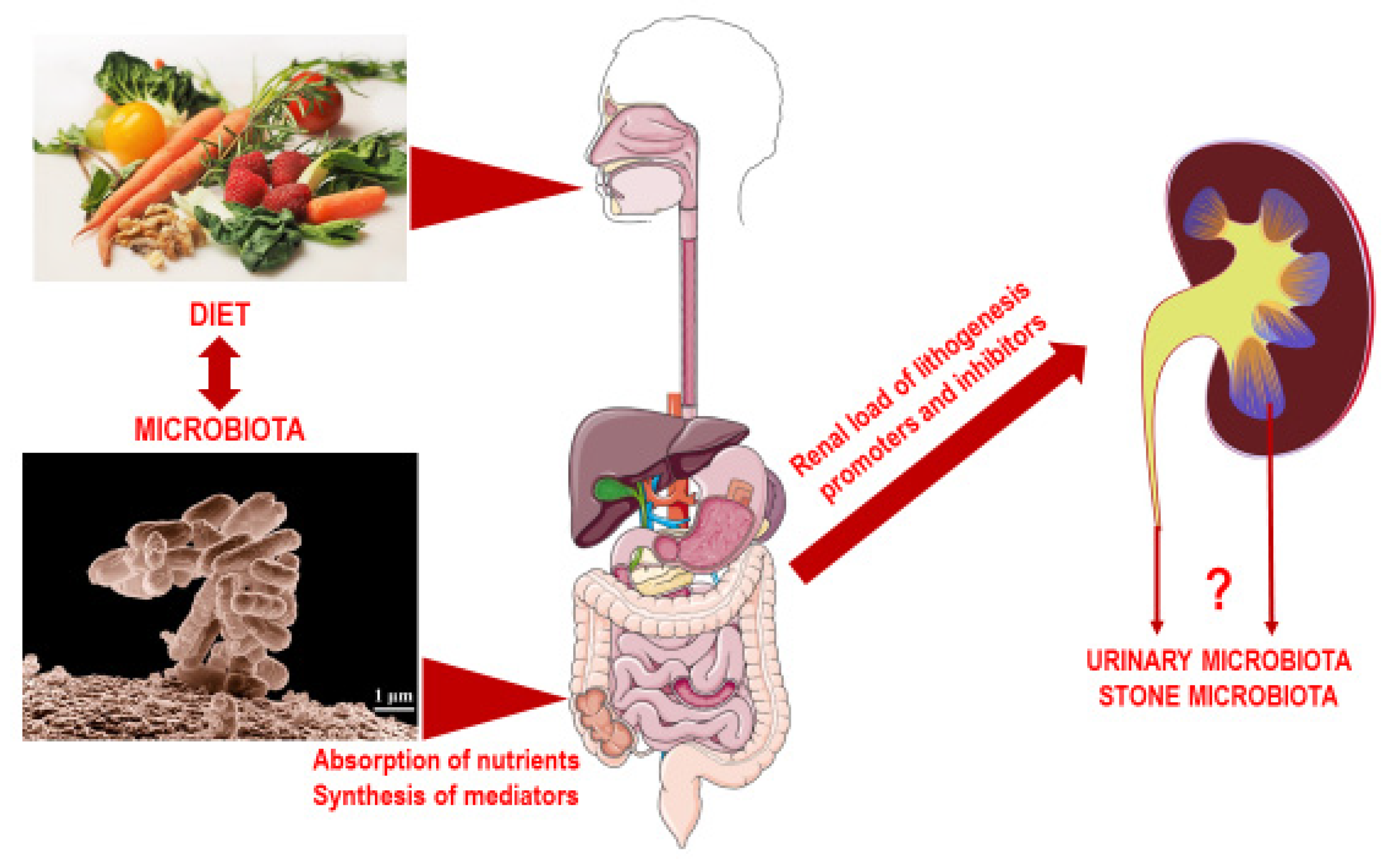
1. Introduction
2. Gut Microbiota and Calcium Oxalate Stone Disease: An Overview
2.1. Before the Microbiota Revolution: Focus on Oxalobacter
2.2. Beyond the Microbiota Revolution: Oxalobacter as Part of a Network
3. The Role of Diet in the Gut-Kidney Axis
3.1. The Determinants of Nephrolithiasis-Associated Gut Microbiota Dysbiosis
3.2. Salt and Microbiota
3.3. Animal Proteins and Microbiota
3.4. Oxalate Intake and Microbiota
3.5. Calcium Intake and Microbiota
3.6. FAV Intake, legume Intake, and Microbiota
3.7. Water Intake and Gut Microbiota
4. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coe, F.L.; Parks, J.H.; Asplin, J.R. The pathogenesis and treatment of kidney stones. N. Engl. J. Med. 1992, 327, 1141–1152. [Google Scholar] [CrossRef]
- Sakhaee, K. Recent advances in the pathophysiology of nephrolithiasis. Kidney Int. 2009, 75, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Coe, F.L.; Worcester, E.M. Clinical practice. Calcium kidney stones. N. Engl. J. Med. 2010, 363, 954–963. [Google Scholar]
- Pak, C.Y.; Sakhaee, K.; Moe, O.W.; Poindexter, J.; Adams-Huet, B.; Pearle, M.S.; Zerwekh, J.E.; Premiger, G.M.; Wills, M.R.; Breslau, N.A.; et al. Defining hypercalciuria in nephrolithiasis. Kidney Int. 2011, 80, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Robijn, S.; Hoppe, B.; Vervaet, B.A.; D’Haese, P.C.; Verhulst, A. Hyperoxaluria: A gut-kidney axis? Kidney Int. 2011, 80, 1146–1158. [Google Scholar] [CrossRef]
- Whittamore, J.M.; Hatch, M. The role of intestinal oxalate transport in hyperoxaluria and the formation of kidney stones in animals and man. Urolithiasis 2017, 45, 89–108. [Google Scholar] [CrossRef]
- Ticinesi, A.; Nouvenne, A.; Meschi, T. Gut microbiome and kidney stone disease: Not just an Oxalobacter story. Kidney Int. 2019, 96, 25–27. [Google Scholar] [CrossRef]
- Siva, S.; Barrack, E.R.; Reddy, G.P.; Thamilselvan, V.; Thamilselvan, S.; Menon, M.; Bhandari, M. A critical analysis of the role of Oxalobacter formigenes in oxalate stone disease. BJU Int. 2009, 103, 18–21. [Google Scholar] [CrossRef]
- Hoesl, C.E.; Altwein, J.E. The probiotic approach: An alternative treatment option in urology. Eur. Urol. 2005, 47, 288–296. [Google Scholar] [CrossRef]
- Sadaf, H.; Raza, S.I.; Hassan, S.W. Role of gut microbiota against calcium oxalate. Microb. Pathog. 2017, 109, 287–291. [Google Scholar] [CrossRef]
- Rescigno, M. The microbiota revolution: Excitement and caution. Eur. J. Immunol. 2017, 47, 1406–1413. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Chen, D.Q.; Chen, L.; Liu, J.R.; Vaziri, N.D.; Guo, Y.; Zhao, Y.Y. Microbiome-metabolome reveals the contribution of gut-kidney axis on kidney disease. J. Transl. Med. 2019, 17, 5. [Google Scholar] [CrossRef] [PubMed]
- Mehta, M.; Goldfarb, D.S.; Nazzal, L. The role of the microbiome in kidney stone formation. Int. J. Surg. 2016, 36, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Curhan, G.C.; Willett, W.C.; Rimm, E.B.; Stampfer, M.J. A prospective study of dietary calcium and other nutrients and the risk of symptomatic kidney stones. N. Engl. J. Med. 1993, 328, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Curhan, G.C.; Willett, W.C.; Speizer, F.E.; Spiegelman, D.; Stampfer, M.J. Comparison of dietary calcium with supplemental calcium and other nutrients as factors affecting the risk for kidney stones in women. Ann. Intern. Med. 1997, 126, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Prezioso, D.; Strazzullo, P.; Lotti, T.; Bianchi, G.; Borghi, L.; Caione, P.; Carini, M.; Caudarella, R.; Ferraro, M.; Gambaro, G.; et al. Dietary treatment of urinary risk factors for renal stone formation. A review of CLU Working Group. Arch. Ital. Urol. Androl. 2015, 57, 105–120. [Google Scholar] [CrossRef]
- Gambaro, G.; Croppi, E.; Coe, F.; Lingeman, J.; Moe, O.; Worcester, E.; Buchholz, N.; Bushinsky, D.; Curhan, G.C.; Ferraro, P.M.; et al. Metabolic diagnosis and medical prevention of calcium nephrolithiasis and its systemic manifestations: A consensus statement. J. Nephrol. 2016, 29, 715–734. [Google Scholar] [CrossRef]
- Dawson, K.A.; Allison, M.J.; Hartman, P.A. Isolation and some characteristics of anaerobic oxalate-degrading bacteria from the rumen. Appl. Environ. Microbiol. 1980, 40, 833–839. [Google Scholar] [CrossRef]
- Allison, M.J.; Dawson, K.A.; Mayberry, W.R.; Foss, J.G. Oxalobacter formigenes gen. nov., sp. nov.: Oxalate-degrading anaerobes that inhabit the gastrointestinal tract. Arch. Microbiol. 1985, 141, 1–7. [Google Scholar] [CrossRef]
- Miller, A.W.; Dearing, D. The metabolic and ecological interactions of oxalate-degrading bacteria in the mammalian gut. Pathogens 2013, 2, 636–652. [Google Scholar] [CrossRef]
- Allison, M.J.; Cook, H.M.; Milne, D.B.; Gallagher, S.; Clayman, R.V. Oxalate degradation by gastrointestinal bacteria from humans. J. Nutr. 1986, 116, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Argenzio, R.A.; Liacos, J.A.; Allison, M.J. Intestinal oxalate-degrading bacteria reduce oxalate absorption and toxicity in guinea pigs. J. Nutr. 1988, 118, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, H.; Holmes, R.P.; Allison, M.J.; Peck, A.B. Direct quantification of the enteric bacterium Oxalobacter formigenes in human fecal samples by quantitative competitive-template PCR. J. Clin. Microbiol. 1999, 37, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.S.; Duncan, S.H.; Cave, D.R. Oxalobacter formigenes and its role in oxalate metabolism in the human gut. FEMS Microbiol. Lett. 2004, 230, 1–7. [Google Scholar] [CrossRef]
- Arvans, D.; Jung, Y.C.; Antonopoulos, D.; Koval, J.; Granja, I.; Bashir, M.; Karrar, E.; Roy-Chowdhury, J.; Musch, M.; Asplin, J.; et al. Oxalobacter formigenes-derived bioactive factors stimulate oxalate transport by intestinal epithelial cells. J. Am. Soc. Nephrol. 2017, 28, 876–887. [Google Scholar] [CrossRef]
- Sidhu, H.; Hoppe, B.; Hesse, A.; Tenbrock, K.; Brömme, S.; Rietschel, E.; Peck, A.B. Absence of Oxalobacter formigenes in cystic fibrosis patients: A risk factor for hyperoxaluria. Lancet 1998, 352, 1026–1029. [Google Scholar] [CrossRef]
- Sidhu, H.; Schmidt, M.E.; Cornelius, J.G.; Thamilselvan, S.; Khan, S.R.; Hesse, A.; Peck, A.B. Direct correlation between hyperoxaluria/oxalate stone disease and the absence of the gastrointestinal tract-dwelling bacterium Oxalobacter formigenes: Possible prevention by gut recolonization or enzyme replacement therapy. J. Am. Soc. Nephrol. 1999, 10, S334–S340. [Google Scholar]
- Kumar, R.; Ghoshal, U.C.; Singh, G.; Mittal, R.D. Infrequency of colonization with Oxalobacter formigenes in inflammatory bowel disease: Possible role in renal stone formation. J. Gastroenterol. Hepatol. 2004, 19, 1403–1409. [Google Scholar] [CrossRef]
- Kaufman, D.W.; Kelly, J.P.; Curhan, G.C.; Anderson, T.E.; Dretler, S.P.; Preminger, G.M.; Cave, D.R. Oxalobacter formigenes may reduce the risk of calcium oxalate kidney stones. J. Am. Soc. Nephrol. 2008, 19, 1197–1203. [Google Scholar] [CrossRef]
- Tavasoli, S.; Alebouyeh, M.; Naji, M.; Shakiba Majd, G.; Shabani Nashtaei, M.; Broumandnia, N.; Basiri, A. Association of intestinal oxalate-degrading bacteria with recurrent calcium kidney stone formation and hyperoxaluria: A case-control study. BJU Int. 2020, 125, 133–143. [Google Scholar] [CrossRef]
- Duffey, B.G.; Miyaoka, R.; Holmes, R.; Assimos, D.; Hinck, B.; Korman, E.; Kieley, F.; Ikramuddin, S.; Kellogg, T.; Moeding, A.; et al. Oxalobacter colonization in the morbidly obese and correlation with urinary stone risk. Urology 2011, 78, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Siener, R.; Bangen, U.; Sidhu, H.; Hönow, R.; von Unruh, G.; Hesse, A. The role of Oxalobacter formigenes colonization in calcium oxalate stone disease. Kidney Int. 2013, 83, 1144–1149. [Google Scholar] [CrossRef] [PubMed]
- Barnett, C.; Nazzal, L.; Goldfarb, D.S.; Blaser, M.J. The presence of Oxalobacter formigenes in the microbiome of healthy young adults. J. Urol. 2016, 195, 499–506. [Google Scholar] [CrossRef] [PubMed]
- PeBenito, A.; Nazzal, L.; Wang, C.; Li, H.; Jay, M.; Noya-Alarcon, O.; Contreras, M.; Lander, O.; Leach, J.; Dominguez-Bello, M.G.; et al. Comparative prevalence of Oxalobacter formigenes in three human populations. Sci. Rep. 2019, 9, 574. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Koh, H.; Kurtz, Z.D.; Battaglia, T.; PeBenito, A.; Li, H.; Nazzal, L.; Blaser, M.J. Oxalobacter formigenes-associated host features and microbial community structures examined using the American Gut Project. Microbiome 2017, 5, 108. [Google Scholar] [CrossRef] [PubMed]
- Campieri, C.; Campieri, M.; Bertuzzi, V.; Swennen, E.; Matteuzzi, D.; Stefoni, S.; Pirovano, F.; Centi, C.; Ulisse, S.; Famularo, G.; et al. Reduction of oxaluria after an oral course of lactic acid bacteria at high concentration. Kidney Int. 2001, 60, 1097–1105. [Google Scholar] [CrossRef]
- Duncan, S.H.; Richardson, A.J.; Kaul, P.; Holmes, R.P.; Allison, M.J.; Stewart, C.S. Oxalobacter formigenes and its potential role in human health. Appl. Environ. Microbiol. 2002, 68, 3841–3847. [Google Scholar] [CrossRef]
- Lieske, J.C.; Goldfarb, D.S.; De Simone, C.; Regnier, C. Use of a probiotic to decrease enteric hyperoxaluria. Kidney Int. 2005, 68, 1244–1249. [Google Scholar] [CrossRef]
- Goldfarb, D.S.; Modersitzki, F.; Asplin, J.R. A randomized, controlled trial of lactic acid bacteria for idiopathic hyperoxaluria. Clin. J. Am. Soc. Nephrol. 2007, 2, 745–749. [Google Scholar] [CrossRef]
- Okombo, J.; Liebman, M. Probiotic-induced reduction of gastrointestinal oxalate absorption in healthy subjects. Urol. Res. 2010, 38, 169–178. [Google Scholar] [CrossRef]
- Hoppe, B.; Groothoff, J.W.; Hulton, S.A.; Cochat, P.; Niaudet, P.; Kemper, M.J.; Deschênes, G.; Unwin, R.; Milliner, D. Efficacy and safety of Oxalobacter formigenes to reduce urinary oxalate in primary hyperoxaluria. Nephrol. Dial. Transplant. 2011, 26, 3609–3615. [Google Scholar] [CrossRef] [PubMed]
- Al-Wahsh, I.; Wu, Y.; Liebman, M. Acute probiotic ingestion reduces gastrointestinal oxalate absorption in healthy subjects. Urol. Res. 2012, 40, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Siener, R.; Bade, D.J.; Hesse, A.; Hoppe, B. Dietary hyperoxaluria is not reduced by treatment with lactic acid bacteria. J. Transl. Med. 2013, 11, 306. [Google Scholar] [CrossRef] [PubMed]
- Suryavanshi, M.; Bhute, S.S.; Jadhav, S.D.; Bhatia, M.S.; Gune, R.P.; Shouche, Y.S. Hyperoxaluria leads to dysbiosis and drives selective enrichment of oxalate metabolizing bacterial species in recurrent kidney stone formers. Sci. Rep. 2016, 6, 34712. [Google Scholar] [CrossRef]
- Stern, J.M.; Moazami, S.; Qiu, Y.; Kurland, I.; Chen, Z.; Agalliu, I.; Burk, R.; Davies, K.P. Evidence for a distinct gut microbiome in kidney stone formers compared to non-stone formers. Urolithiasis 2016, 44, 399–407. [Google Scholar] [CrossRef]
- Tang, R.; Jiang, Y.; Tan, A.; Ye, J.; Xian, X.; Xie, Y.; Wang, Q.; Yao, Z.; Mo, Z. 16S rRNA gene sequencing reveals altered composition of gut microbiota in individuals with kidney stones. Urolithiasis 2018, 46, 503–514. [Google Scholar] [CrossRef]
- Ticinesi, A.; Milani, C.; Guerra, A.; Allegri, F.; Lauretani, F.; Nouvenne, A.; Mancabelli, L.; Lugli, G.A.; Turroni, F.; Duranti, S.; et al. Understanding the gut-kidney axis in nephrolithiasis: An analysis of the gut microbiota composition and functionality of stone formers. Gut 2018, 67, 2097–2106. [Google Scholar] [CrossRef]
- Suryavanshi, M.V.; Bhute, S.S.; Gune, R.P.; Shouche, Y.S. Functional eubacteria species along with trans-domain gut inhabitants favour dysgenic diversity in oxalate stone disease. Sci. Rep. 2018, 8, 16598. [Google Scholar] [CrossRef]
- Miller, A.W.; Choi, D.; Penniston, K.L.; Lange, D. Inhibition of urinary stone disease by a multi-species bacterial network ensures healthy oxalate homeostasis. Kidney Int. 2019, 96, 180–188. [Google Scholar] [CrossRef]
- Zampini, A.; Nguyen, A.H.; Rose, E.; Monga, M.; Miller, A.W. Defining dysbiosis in patients with urolithiasis. Sci. Rep. 2019, 9, 5425. [Google Scholar] [CrossRef]
- Dornbier, R.A.; Bajic, P.; Van Kuiken, M.; Jardaneh, A.; Lin, H.; Gao, X.; Knudsen, B.; Dong, Q.; Wolfe, A.J.; Schwaderer, A.L. The microbiome of calcium-based urinary stones. Urolithiasis 2019. [Google Scholar] [CrossRef] [PubMed]
- PeBenito, A.M.; Liu, M.; Nazzal, L.; Blaser, M.J. Development of humanized murine model for the study of Oxalobacter formigenes intestinal colonization. J. Infect. Dis. 2019, 220, 1848–1858. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Goldfarb, D.S. The use of antibiotics and risk of kidney stones. Curr. Opin. Nephrol. Hypertens. 2019, 28, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Tasian, G.E.; Jemielita, T.; Goldfarb, D.S.; Copelovitch, L.; Gerber, J.S.; Wu, Q.; Denburg, M.R. Oral antibiotic exposure and kidney stone disease. J. Am. Soc. Nephrol. 2018, 29, 1731–1740. [Google Scholar] [CrossRef]
- Ferraro, P.M.; Curhan, G.C.; Gambaro, G.; Taylor, E.N. Antibiotic use and risk of incident kidney stones in female nurses. Am. J. Kidney Dis. 2019, 74, 736–741. [Google Scholar] [CrossRef]
- Schmidt, T.S.B.; Raes, J.; Bork, P. The human gut microbiome: From association to modulation. Cell 2018, 172, 1198–1215. [Google Scholar] [CrossRef]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S.; et al. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 2016, 352, 565–569. [Google Scholar] [CrossRef]
- Al Zahrani, H.; Norman, R.W.; Thompson, C.; Weerasinghe, S. The dietary habits of idiopathic calcium stone-formers and normal control subjects. BJU Int. 2000, 85, 616–620. [Google Scholar] [CrossRef]
- Meschi, T.; Nouvenne, A.; Ticinesi, A.; Prati, B.; Guerra, A.; Allegri, F.; Pigna, F.; Soldati, L.; Vezzoli, G.; Gambaro, G.; et al. Dietary habits in women with idiopathic calcium nephrolithiasis. J. Transl. Med. 2012, 10, 63. [Google Scholar] [CrossRef]
- Sorensen, M.D.; Kahn, A.J.; Reiner, A.P.; Tseng, T.Y.; Shikany, J.M.; Wallace, R.B.; Chi, T.; Wactawski-Wende, J.; Jackson, R.D.; O’Sullivan, M.J.; et al. Impact of nutritional factors on incident kidney stone formation: A report from the WHI OS. J. Urol. 2012, 187, 1645–1649. [Google Scholar] [CrossRef]
- Ticinesi, A.; Nouvenne, A.; Maalouf, N.M.; Borghi, L.; Meschi, T. Salt and nephrolithiasis. Nephrol. Dial. Transplant. 2016, 31, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Borghi, L.; Schianchi, T.; Meschi, T.; Guerra, A.; Allegri, F.; Maggiore, U.; Novarini, A. Comparison of two diets for the prevention of recurrent stones in idiopathic hypercalciuria. N. Engl. J. Med. 2002, 346, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Nouvenne, A.; Meschi, T.; Prati, B.; Guerra, A.; Allegri, F.; Vezzoli, G.; Soldati, L.; Gambaro, G.; Maggiore, U.; Borghi, L. Effects of a low-salt diet on idiopathic hypercalciuria in calcium-oxalate stone formers: A 3-mo randomized controlled trial. Am. J. Clin. Nutr. 2010, 91, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, M.; Capasso, G.; Di Leo, V.A.; Di Santo, N.G. A history of salt. Am. J. Nephrol. 1994, 14, 426–431. [Google Scholar]
- Derkach, A.; Sampson, J.; Joseph, J.; Playdon, M.C.; Stolzenberg-Solomon, R.Z. Effects of dietary sodium on metabolites: The Dietary Approaches to Stop Hypertension (DASH)-Sodium Feeding Study. Am. J. Clin. Nutr. 2017, 106, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Luo, H.; Wang, J.; Tang, W.; Lu, L.; Wu, S.; Xiong, Z.; Yang, G.; Chen, Z.; Lan, T.; et al. Enteric dysbiosis-linked gut barrier disruption triggers early renal injury induced by chronic high salt feeding in mice. Exp. Mol. Med. 2017, 49, e370. [Google Scholar] [CrossRef] [PubMed]
- Wilck, N.; Matus, M.G.; Kearney, S.M.; Olesen, S.W.; Forslund, K.; Bartolomaeus, H.; Haase, S.; Mӓhler, A.; Balogh, A.; Markó, L.; et al. Salt-responsive gut commensal modulates TH17 axis and disease. Nature 2017, 551, 585–589. [Google Scholar] [CrossRef]
- Bielinska, K.; Radkowski, M.; Grochowska, M.; Perlejewski, K.; Huc, T.; Jaworska, K.; Motooka, D.; Nakamura, S.; Ufnal, M. High salt intake increases plasma trimethylamine N-oxide (TMAO) concentration and produces gut dysbiosis in rats. Nutrition 2018, 54, 33–39. [Google Scholar] [CrossRef]
- Wang, H.; Li, S.; Fang, S.; Yang, X.; Feng, J. Betaine improves intestinal functions by enhancing digestive enzymes, ameliorating intestinal morphology, and enriching intestinal microbiota in high-salt stressed rats. Nutrients 2018, 10, 907. [Google Scholar] [CrossRef]
- Seck, E.H.; Senghor, B.; Merhej, V.; Bachar, D.; Cadoret, F.; Robert, C.; Azhar, E.I.; Yasir, M.; Bibi, F.; Jiman-Fatani, A.A.; et al. Salt in stools is associated with obesity, gut halophilic microbiota and Akkermansia muciniphila depletion in humans. Int. J. Obes. 2019, 43, 862–871. [Google Scholar] [CrossRef]
- Smiljanec, K.; Lennon, S.L. Sodium, hypertension, and the gut: Does the microbiota go salty? Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H1173–H1182. [Google Scholar] [CrossRef] [PubMed]
- Amanzadeh, J.; Gitomer, W.L.; Zerwekh, J.E.; Preisig, P.A.; Moe, O.W.; Pak, C.Y.C.; Levi, M. Effect of high protein diet on stone-forming propensity and bone loss in rats. Kidney Int. 2003, 64, 2142–2149. [Google Scholar] [CrossRef] [PubMed]
- Vezzoli, G.; Dogliotti, E.; Terranegra, A.; Arcidiacono, T.; Macrina, L.; Tavecchia, M.; Pivari, F.; Mingione, A.; Brasacchio, C.; Nouvenne, A.; et al. Dietary style and acid load in an Italian population of calcium kidney stone formers. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Ferraro, P.M.; Mandel, E.I.; Curhan, G.C.; Gambaro, G.; Taylor, E.N. Dietary protein and potassium, diet-dependent net acid load, and risk of incident kidney stones. Clin. J. Am. Nephrol. 2016, 11, 1834–1844. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Hughes, B.; Harris, S.S.; Rasmussen, H.M.; Dallal, G.E. Comparative effects of oral aromatic and branched-chain amino acids on urine calcium excretion in humans. Osteoporos. Int. 2007, 18, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Bihuniak, J.D.; Sullivan, R.R.; Simpson, C.A.; Caseria, D.M.; Huedo-Medina, T.B.; O’Brien, K.O.; Kerstetter, J.E.; Insogna, K.L. Supplementing a low-protein diet with dibasic amino acids increases urinary calcium excretion in young women. J. Nutr. 2014, 144, 282–288. [Google Scholar] [CrossRef]
- Ince, B.A.; Anderson, E.J.; Neer, R.M. Lowering dietary protein to U.S. Recommended Dietary Allowance levels reduces urinary calcium excretion and bone resorption in young women. J. Clin. Endocrinol. Metab. 2004, 89, 3801–3807. [Google Scholar] [CrossRef]
- Mu, C.; Yang, Y.; Luo, Z.; Guan, L.; Zhu, W. The colonic microbiome and epithelial transcriptome are altered in rats fed a high-protein diet compared with a normal-protein diet. J. Nutr. 2016, 146, 474–483. [Google Scholar] [CrossRef]
- Kostovcikova, K.; Coufal, S.; Galanova, N.; Fajstova, A.; Hudcovic, T.; Kostovcik, M.; Prochazkova, P.; Jiraskova Zakosteslka, Z.; Cermakova, M.; Sediva, B.; et al. Diet rich in animal protein promotes pro-inflammatory macrophage response and exacerbates colitis in mice. Front. Immunol. 2019, 10, 919. [Google Scholar] [CrossRef]
- Umair Ijaz, M.; Ijaz Ahmed, M.; Zou, X.; Hussain, M.; Zhang, M.; Zhao, F.; Xu, X.; Zhou, G.; Li, C. Beef, casein, and soy proteins differently affect lipid metabolism, triglycerides accumulation and gut microbiota of high-fat diet-fed C57BL/6J mice. Front. Immunol. 2018, 9, 2200. [Google Scholar]
- Zhao, F.; Zhou, G.; Liu, X.; Song, S.; Xu, X.; Hooiveld, G.; Müller, M.; Liu, L.; Kristiansen, K.; Li, C. Dietary protein sources differentially affect the growth of Akkermansia muciniphila and maintenance of the gut mucus barrier in mice. Mol. Nutr. Food Res. 2019, 63, 1900589. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Shi, X.; Lin, X.; Ye, K.; Xu, X.; Li, C.; Zhou, G. Beef, chicken, and soy proteins in diets induce different gut microbiota and metabolites in rats. Front. Microbiol. 2017, 8, 1395. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Xu, W.; Guo, M.; Chen, S.; Liu, Y.; He, X.; Huang, K. Rice- or pork-based diets with similar calorie and content result in different rat gut microbiota. Int. J. Food Sci. Nutr. 2017, 68, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lin, X.; Zhao, F.; Shi, X.; Li, H.; Li, Y.; Zhu, W.; Xu, X.; Li, C.; Zhou, G. Meat, dairy and plant proteins alter bacterial composition of rat gut bacteria. Sci. Rep. 2015, 5, 15220. [Google Scholar] [CrossRef]
- Zhu, Y.; Lin, X.; Li, H.; Li, Y.; Shi, X.; Zhao, F.; Xu, X.; Li, C.; Zhou, G. Intake of meat proteins substantially increased the relative abundance of genus Lactobacillus in rat feces. PLoS ONE 2016, 11, e0152678. [Google Scholar] [CrossRef]
- Vidal-Lletjós, S.; Andriamihaja, M.; Blais, A.; Grauso, M.; Lepage, P.; Davila, A.M.; Viel, R.; Gaudichon, C.; Leclerc, M.; Blachier, F.; et al. Dietary protein intake level modulates mucosal healing and mucosa-adherent microbiota in mouse model of colitis. Nutrients 2019, 11, 514. [Google Scholar] [CrossRef]
- Kiilerich, P.; Secher Myrmel, L.; Fjære, E.; Hao, Q.; Hugenholz, F.; Brask Sonne, S.; Derrien, M.; Møller Pedersen, L.; Koefoed Petersen, R.; Mortensen, A.; et al. Effect of a long-term high-protein diet on survival, obesity development, and gut microbiota in mice. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E886–E899. [Google Scholar] [CrossRef]
- Blachier, F.; Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Audebert, M.; Khodorova, N.; Andriamihaja, M.; Airinei, G.; Benamouzig, R.; et al. High-protein diets for weight management: Interactions with the intestinal microbiota and consequences for gut health. A position paper by the my new gut study group. Clin. Nutr. 2019, 38, 1012–1022. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef]
- Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Cerrudo, V.; Audebert, M.; Dumont, F.; Mancano, G.; Khodorova, N.; Andriamihaja, M.; et al. Quantity and source of dietary protein influence metabolite production by gut microbiota and rectal mucosa gene expression: A randomized, parallel, double-blind trial in overweight humans. Am. J. Clin. Nutr. 2017, 106, 1005–1019. [Google Scholar] [CrossRef]
- Wei, R.; Ross, A.B.; Su, M.M.; Wang, J.; Guiraud, S.P.; Fogarty Draper, C.; Beaumont, M.; Jia, W.; Martin, F.P. Metabotypes related to meat and vegetable intake reflect microbial, lipid and amino acid metabolism in healthy people. Mol. Nutr. Food Res. 2018, 62, 1800583. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on gut microbiota associated to a Mediterranean diet adherence and specific dietary intakes on general adult population. Front. Microbiol. 2018, 9, 890. [Google Scholar] [CrossRef]
- Leone, A.; Fernández-Montero, A.; de la Fuente-Arrillaga, C.; Martínez-González, M.A.; Bertoli, S.; Battezzati, A.; Bes-Rastrollo, M. Adherence to the Mediterranean dietary pattern and incidence of nephrolithiasis in the Seguimiento Universidad de Navarra Follow-up (SUN) cohort. Am. J. Kidney Dis. 2017, 70, 778–786. [Google Scholar] [CrossRef]
- Taylor, E.N.; Fung, T.T.; Curhan, G.C. DASH-style diet associates with reduced risk for kidney stones. J. Am. Soc. Nephrol. 2009, 20, 2253–2259. [Google Scholar] [CrossRef]
- Taylor, E.N.; Stampfer, M.J.; Mount, D.B.; Curhan, G.C. DASH-style diet and 24-hour urine composition. Clin. J. Am. Soc. Nephrol. 2010, 5, 2315–2322. [Google Scholar] [CrossRef]
- Siener, R.; Ebert, D.; Nicolay, C.; Hesse, A. Dietary risk factors for hyperoxaluria in calcium oxalate stone formers. Kidney Int. 2003, 63, 1037–1043. [Google Scholar] [CrossRef]
- Lange, J.N.; Wood, K.D.; Mufarrij, P.W.; Callahan, M.F.; Easter, L.; Knight, J.; Holmes, R.P.; Assimos, D.G. The impact of dietary calcium and oxalate ratios on stone risk. Urology 2012, 79, 1226–1229. [Google Scholar] [CrossRef]
- Knight, J.; Jiang, J.; Assimos, D.G.; Holmes, R.P. Hydroxyproline ingestion and urinary oxalate and glycolate excretion. Kidney Int. 2006, 70, 1929–1934. [Google Scholar] [CrossRef]
- Bergsland, K.J.; Zisman, A.L.; Asplin, J.R.; Worcester, E.M.; Coe, F.L. Evidence for net renal tubule oxalate secretion in patients with calcium kidney stones. Am. J. Physiol. Renal Physiol. 2011, 300, F311–F318. [Google Scholar] [CrossRef]
- Taylor, E.N.; Curhan, G.C. Oxalate intake and risk for nephrolithiasis. J. Am. Soc. Nephrol. 2007, 18, 2198–2204. [Google Scholar] [CrossRef]
- Khan, S.R.; Glenton, P.A.; Byer, K.J. Dietary oxalate and calcium oxalate nephrolithiasis. J. Urol. 2007, 178, 2191–2196. [Google Scholar] [CrossRef]
- Thomas, E.; von Unruh, G.E.; Hesse, A. Influence of a low- and high-oxalate vegetarian diet on intestinal oxalate absorption and urinary excretion. Eur. J. Clin. Nutr. 2008, 62, 1090–1097. [Google Scholar] [CrossRef]
- Nouvenne, A.; Meschi, T.; Guerra, A.; Allegri, F.; Prati, B.; Fiaccadori, E.; Maggiore, U.; Borghi, L. Diet to reduce mild hyperoxaluria in patients with idiopathic calcium oxalate stone formation: A pilot study. Urology 2009, 73, 725–730. [Google Scholar] [CrossRef]
- Noori, N.; Honarkar, E.; Goldfarb, D.S.; Kalantar-Zadeh, K.; Taheri, M.; Shakhssaklim, N.; Parvin, M.; Basiri, A. Urinary lithogenic risk profile in recurrent stone formers with hyperoxaluria: A randomized controlled trial comparing DASH (Dietary Approaches to Stop Hypertension)-style and low-oxalate diets. Am. J. Kidney Dis. 2014, 63, 456–463. [Google Scholar] [CrossRef]
- Miller, A.W.; Kohl, K.D.; Dearing, D. The gastrointestinal tract of the white-throated woodrat (Neotoma albigula) harbors distinct consortia of oxalate-degrading bacteria. Appl. Environ. Microbiol. 2014, 80, 1595–1601. [Google Scholar] [CrossRef]
- Miller, A.W.; Oakeson, K.F.; Dale, C.; Dearing, M.D. Effect of dietary oxalate on the gut microbiota of the mammalian herbivore Neotoma albigula. Appl. Environ. Microbiol. 2016, 82, 2669–2675. [Google Scholar] [CrossRef]
- Miller, A.W.; Dale, C.; Dearing, M.D. Microbiota diversification and crash induced by dietary oxalate in the mammalian herbivore Neotoma albigula. mSphere 2017, 2, e00428-17. [Google Scholar] [CrossRef]
- Ukhanova, M.; Wang, X.; Baer, D.J.; Novotny, J.A.; Fredborg, M.; Mai, V. Effect of almond and pistachio consumption on gut microbiota composition in a randomised cross-over human feeding study. Br. J. Nutr. 2014, 111, 2146–2152. [Google Scholar] [CrossRef]
- Holscher, H.D.; Taylor, A.M.; Swanson, K.S.; Novotny, J.A.; Baer, D.J. Almond consumption and processing affects the composition of gastrointestinal microbiota of healthy adult men and women: A randomized controlled trial. Nutrients 2018, 10, 126. [Google Scholar] [CrossRef]
- Holscher, H.D.; Guetterman, H.M.; Swanson, K.S.; An, R.; Matthan, N.R.; Lichtenstein, A.H.; Novotny, J.A.; Baer, D.J. Walnut consumption alters the gastrointestinal microbiota, microbially derived secondary bile acids, and health markers in healthy adults: A randomized controlled trial. J. Nutr. 2018, 148, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Bamberger, C.; Rossmeier, A.; Lechner, K.; Wu, L.; Waldmann, E.; Fischer, S.; Stark, R.G.; Altenhofer, J.; Henze, K.; Parhofer, K.G. A walnut-enriched diet affects gut microbiome in healthy Caucasian subjects: A randomized, controlled trial. Nutrients 2018, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Byerley, L.O.; Samuelson, D.; Blanchard IV, E.; Luo, M.; Lorenzen, B.N.; Banks, S.; Ponder, M.A.; Welsh, D.A.; Taylor, C.M. Changes in the gut microbial communities following addiction of walnuts to diet. J. Nutr. Biochem. 2017, 48, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Gargari, G.; Deon, V.; Taverniti, V.; Gardana, C.; Denina, M.; Riso, P.; Guardamagna, O.; Guglielmetti, S. Evidence of dysbiosis in the intestinal microbial ecosystem of children and adolescents with primary hyperlipidemia and the potential role of regular hazelnut intake. FEMS Microbiol. Ecol. 2018, 94, fiy045. [Google Scholar] [CrossRef] [PubMed]
- Calani, L.; Dall’Asta, M.; Derlindati, E.; Scazzina, F.; Bruni, R.; Del Rio, D. Colonic metabolism of polyphenols from coffee, green tea, and hazelnut skins. J. Clin. Gastroenterol. 2012, 46, S95–S99. [Google Scholar] [CrossRef]
- Haaskjold, Y.L.; Drotningsvik, A.; Leh, S.; Marti, H.P.; Svarstad, E. Renal failure due to excessive intake of almonds in the absence of Oxalobacter formigenes. Am. J. Med. 2015, 128, e29–e30. [Google Scholar] [CrossRef]
- Taylor, E.N.; Curhan, G.C. Dietary calcium from dairy and nondairy sources, and risk of symptomatic kidney stones. J. Urol. 2013, 190, 1255–1259. [Google Scholar] [CrossRef]
- Heller, H.J.; Doerner, M.F.; Brinkley, L.J.; Adams-Huet, B.; Pak, C.Y.C. Effect of dietary calcium on stone forming propensity. J. Urol. 2003, 169, 470–474. [Google Scholar] [CrossRef]
- Mourad, B.; Fadwa, N.; Mounir, T.; Abdelhamid, E.; Fadhel, N.M.; Rachid, S. Influence of hypercalcic and/or hyperoxalic diet on calcium oxalate renal stone formation in rats. Scand. J. Urol. Nephrol. 2006, 40, 187–191. [Google Scholar] [CrossRef]
- Domrongkitchaiporn, S.; Sopassathit, W.; Stitchantrakul, W.; Prapaipanich, S.; Ingsathit, A.; Rajatanavin, R. Schedule of taking calcium supplement and the risk of nephrolithiasis. Kidney Int. 2004, 65, 1835–1841. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Taylor, E.N.; Curhan, G.C. Demographic, dietary, and urinary factors and 24-h urinary calcium excretion. Clin. J. Am. Soc. Nephrol. 2009, 4, 1980–1987. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, M.D.; Eisner, B.H.; Stone, K.L.; Kahn, A.J.; Lui, L.Y.; Sadetsky, N.; Stoller, M.L. Impact of calcium intake and intestinal calcium absorption on kidney stones in older women: The study of osteoporotic fractures. J. Urol. 2012, 187, 1287–1292. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vezzoli, G.; Macrina, L.; Rubinacci, A.; Spotti, D.; Arcidiacono, T. Intestinal calcium absorption among hypercalciuric patients with or without calcium kidney stones. Clin. J. Am. Soc. Nephrol. 2016, 11, 1450–1455. [Google Scholar] [CrossRef][Green Version]
- Chaplin, A.; Parra, P.; Laraichi, S.; Serra, F.; Palou, A. Calcium supplementation modulates gut microbiota in a prebiotic manner in dietary obese mice. Mol. Nutr. Food Res. 2016, 60, 468–480. [Google Scholar] [CrossRef]
- Li, P.; Tang, T.; Chang, X.; Fan, X.; Chen, X.; Wang, R.; Fan, C.; Qi, K. Abnormality in maternal dietary calcium intake during pregnancy and lactation promotes body weight gain by affecting the gut microbiota in mouse offspring. Mol. Nutr. Food Res. 2019, 63, 1800399. [Google Scholar] [CrossRef]
- Krupa-Kozak, U.; Markiewicz, L.H.; Lamparski, G.; Juśkiewicz, J. Administration of inulin-supplemented gluten-free diet modified calcium absorption and caecal microbiota in rats in a calcium-dependent manner. Nutrients 2017, 9, 702. [Google Scholar] [CrossRef]
- Sorensen, M.D.; Hsi, R.S.; Chi, T.; Shara, N.; Wactawski-Wende, J.; Kahn, A.J.; Wang, H.; Hou, L.; Stoller, M.L. Dietary intake of fiber, fruit, and vegetables decrease the risk of incident kidney stones in women: A Women’s Health Initiative (WHI) report. J. Urol. 2014, 192, 1694–1699. [Google Scholar] [CrossRef]
- Meschi, T.; Maggiore, U.; Fiaccadori, E.; Schianchi, T.; Bosi, S.; Adorni, G.; Ridolo, E.; Guerra, A.; Allegri, F.; Novarini, A.; et al. The effects of fruits and vegetables on urinary stone risk factors. Kidney Int. 2004, 66, 2402–2410. [Google Scholar] [CrossRef]
- Saw, N.K.; Chow, K.; Rao, P.N.; Kavanagh, J.P. Effects of inositol hexaphosphate (phytate) on calcium binding, calcium oxalate crystallization and in vitro stone growth. J. Urol. 2007, 177, 2366–2370. [Google Scholar] [CrossRef]
- Grases, F.; Saez-Torres, C.; Rodriguez, A.; Costa-Bauza, A.; Rodrigo, D.; Frontera, G.; Berga, F.; Fakier, S. Urinary phytate (myo-inositol hexaphosphate) in healthy school children and risk of nephrolithiasis. J. Ren. Nutr. 2014, 24, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Massey, L.K.; Palmer, R.G.; Horner, H.T. Oxalate content of soybean seeds (Glycine max: Leguminosae), soyfoods, and other edible legumes. J. Agric. Food Chem. 2001, 49, 4262–4266. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Zhao, A.; Liu, A.; You, L.; Wang, P. Dietary factors and risk of kidney stones: A case-control study in southern China. J. Ren. Nutr. 2013, 23, e21–e28. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.S.F.; Walker, A.W.; Louis, P.; Parkhill, J.; Vermeiren, J.; Bosscher, D.; Duncan, S.H.; Flint, H.J. Modulation of the human gut microbiota by dietary fibres occurs at the species level. BMC Biol. 2016, 14, 3. [Google Scholar] [CrossRef]
- Lin, D.; Peters, B.A.; Friedlander, C.; Freiman, H.J.; Goedert, J.J.; Sinha, R.; Miller, G.; Bernstein, M.A.; Hayes, R.B.; Ahn, J. Association of dietary fibre intake and gut microbiota in adults. Br. J. Nutr. 2018, 120, 1014–1022. [Google Scholar] [CrossRef]
- Tornova, A.; Bukovsky, I.; Rembert, E.; Yonas, W.; Alwarith, J.; Barnard, N.D.; Kahleova, H. The effects of vegetarian and vegan diets on gut microbiota. Front. Nutr. 2019, 6, 47. [Google Scholar]
- Tap, J.; Furet, J.P.; Bensaada, M.; Philippe, C.; Roth, H.; Rabot, S.; Lakhdari, O.; Lombard, V.; Henrissat, B.; Corthier, G.; et al. Gut microbiota richness promotes its stability upon increased dietary fibre intake in healthy adults. Environ. Microbiol. 2015, 17, 4954–4964. [Google Scholar] [CrossRef]
- Hiel, S.; Bindels, L.B.; Pachikian, B.D.; Kalala, G.; Broers, V.; Zamariola, G.; Chang, B.I.P.; Kambashi, B.; Rodriguez, J.; Cani, P.D.; et al. Effects of a diet based on inulin-rich vegetables on gut health and nutritional behavior in healthy humans. Am. J. Clin. Nutr. 2019, 109, 1683–1695. [Google Scholar] [CrossRef]
- Chambers, E.S.; Byrne, C.S.; Morrison, D.J.; Murphy, K.G.; Preston, T.; Tedford, C.; Garcia-Perez, I.; Fountana, S.; Serrano-Contreras, J.I.; Holmes, E.; et al. Dietary supplementation with inulin-propionate ester or inulin improves insulin sensitivity in adults with overweight and obesity with distinct effects on the gut microbiota, plasma metabolome and systemic inflammatory response: A randomized cross-over trial. Gut 2019, 68, 1430–1438. [Google Scholar]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Menni, C.; Jackson, M.A.; Pallister, T.; Steves, C.J.; Spector, T.D.; Valdes, A.M. Gut microbiome diversity and high-fibre intake are related to lower long-term weight gain. Int. J. Obes. 2017, 41, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Pallister, T.; Jackson, M.A.; Martin, T.C.; Glastonbury, C.A.; Jennings, A.; Beaumont, M.; Mohney, R.P.; Small, K.S.; MacGregor, A.; Steves, C.J.; et al. Untangling the relationship between diet and visceral fat mass through blood metabolomics and gut microbiome profiling. Int. J. Obes. 2017, 41, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Mayengbam, S.; Lambert, J.E.; Parnell, J.A.; Tunnicliffe, J.M.; Nicolucci, A.C.; Han, J.; Sturzenegger, T.; Shearer, J.; Mickiewicz, B.; Vogel, H.J.; et al. Impact of dietary fiber supplementation on modulating microbiota-host-metabolic axes in obesity. J. Nutr. Biochem. 2019, 64, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Klinder, A.; Shen, Q.; Heppel, S.; Lovegrove, J.A.; Rowland, I.; Tuohy, K.M. Impact of increasing fruit and vegetables and flavonoid intake on the human gut microbiota. Food Funct. 2016, 7, 1788–1796. [Google Scholar] [CrossRef]
- Guerra, A.; Ticinesi, A.; Allegri, F.; Nouvenne, A.; Prati, B.; Pinelli, S.; Merli, R.; Tana, C.; Lauretani, F.; Aloe, R.; et al. Insights about urinary hippuric and citric acid as biomarkers of fruit and vegetable intake in patients with kidney stones: The role of age and sex. Nutrition 2019, 59, 83–89. [Google Scholar] [CrossRef]
- Ticinesi, A.; Nouvenne, A.; Borghi, L.; Meschi, T. Water and other fluids in nephrolithiasis: State of the art and future challenges. Crit. Rev. Food Sci. Nutr. 2017, 57, 963–974. [Google Scholar] [CrossRef]
- Choi, Y.J.; Lee, D.H.; Kim, H.S.; Kim, Y.K. An exploratory study on the effect of daily fruits and vegetable juice on human gut microbiota. Food Sci. Biotechnol. 2018, 27, 1377–1386. [Google Scholar] [CrossRef]
- Lima, A.C.D.; Cecatti, C.; Fidélix, M.P.; Adorno, M.A.T.; Sakamoto, I.K.; Cesar, T.B.; Sivieri, K. Effect of daily consumption of orange juice on the levels of blood glucose, lipids, and gut microbiota metabolites: Controlled clinical trials. J. Med. Food 2019, 2, 202–210. [Google Scholar] [CrossRef]
- Brasili, E.; Aymoto Hassimotto, N.M.; Del Chierico, F.; Marini, F.; Quagliariello, A.; Sciubba, F.; Miccheli, A.; Putignani, L.; Lajolo, F. Daily consumption of orange juice from Citrus sinensis L. Osbeck cv. Cara Cara and cv. Bahia differently affects gut microbiota profiling as unveiled by an integrated meta-omics approach. J. Agric. Food Chem. 2019, 67, 1381–1391. [Google Scholar] [CrossRef]
- Henning, S.M.; Yang, J.; Shao, P.; Lee, R.P.; Huang, J.; Ly, A.; Hsu, M.; Lu, Q.Y.; Thames, G.; Heber, D.; et al. Health benefit of vegetable/fruit juice-based diet: Role of microbiome. Sci. Rep. 2017, 7, 2167. [Google Scholar] [CrossRef]
- Mosele, J.I.; Gosalbes, M.J.; Macià, A.; Rubiò, L.; Vazquez-Castellanos, J.F.; Jimenez Hernandez, N.; Moya, A.; Latorre, A.; Motilva, M.J. Effect of daily intake of pomegranate juice on fecal microbiota and feces metabolites from healthy volunteers. Mol. Nutr. Food Res. 2015, 59, 1942–1953. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Krishnan, H.B.; Pham, Q.; Yu, L.L.; Wang, T.T.Y. Soy and gut microbiota: Interaction and implication for human health. J. Agric. Food Chem. 2016, 64, 8695–8709. [Google Scholar] [CrossRef] [PubMed]
- Cross, T.W.L.; Zidon, T.M.; Welly, R.J.; Park, Y.M.; Britton, S.L.; Koch, L.G.; Rottinghaus, G.E.; Cattai de Godoy, M.R.; Padilla, J.; Swanson, K.S.; et al. Soy improves cardiometabolic health and cecal microbiota in female low-fit rats. Sci. Rep. 2017, 7, 9261. [Google Scholar] [CrossRef] [PubMed]
- Siva, N.; Johnson, C.R.; Richard, V.; Jesch, E.D.; Whiteside, W.; Abood, A.A.; Thavarajah, P.; Duckett, S.; Thavarajah, D. Lentil (Lens culinaris Medikus) diet affects the gut microbiome and obesity markers in rat. J. Agric. Food Chem. 2018, 66, 8805–8813. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, L.; Flórez, A.B.; Guadamuro, L.; Mayo, B. Effect of soy isoflavones on growth of representative bacterial species from the human gut. Nutrients 2017, 9, 727. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.F.; Reis, M.P.; Acurcio, L.B.; Carmo, A.O.; Diamantino, C.F.; Motta, A.M.; Kalapothakis, E.; Nicoli, J.R.; Nascimento, A.M.A. Changes in mouse gut bacterial community in response to different types of drinking water. Water Res. 2018, 132, 79–89. [Google Scholar] [CrossRef]
- Johnson, T.B.; Langin, L.M.; Zhao, J.; Weimer, J.M.; Pearce, D.A.; Kovács, A.D. Changes in motor behavior, neuropathology, and gut microbiota of a Batten disease mouse model following administration of acidified drinking water. Sci. Rep. 2019, 9, 14962. [Google Scholar] [CrossRef]
- Barnett, J.A.; Gibson, D.L. H2Oh no! The importance of reporting your water source in your in vivo microbiome studies. Gut Microbes 2019, 10, 261–269. [Google Scholar] [CrossRef]
- Murakami, S.; Goto, Y.; Ito, K.; Hayasaka, S.; Kurihara, S.; Soga, T.; Tomita, M.; Fukuda, S. The consumption of bicarbonate-rich mineral water improves glycemic control. Evid. Based Complement. Alternat. Med. 2015, 2015, 824395. [Google Scholar] [CrossRef]
- Hansen, T.H.; Thomassen, M.T.; Madsen, M.L.; Kern, T.; Bak, E.G.; Kashani, A.; Allin, K.H.; Hansen, T.; Pedersen, O. The effect of drinking water pH on the human gut microbiota and glucose regulation: Results of a randomized controlled cross-over intervention. Sci. Rep. 2018, 8, 16626. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| AUTHOR, YEAR [REF] | METHODS OF MICROBIOTA ANALYSIS | PARTICIPANTS | MAIN RESULTS | NOTES |
|---|---|---|---|---|
| Sidhu H et al., 1998 [26] | Culture + PCR if cultures negative | 43 children with cystic fibrosis, 21 healthy children | Prevalence of Oxalobacter 16% in patients and 71% in controls; patients without Oxalobacter had hyperoxaluria and high stone risk | None of the participants had kidney stones. |
| Sidhu H et al. 1999 [27] | Culture + PCR | 51 adult idiopathic calcium oxalate SFs, 44 healthy volunteers | Prevalence of Oxalobacter: - 75% in controls - 80% in first time stone formers - 38% in recurrent stone formers - 13% in highly recurrent stone formers | Cases and controls inhomogeneous for age and geographical location. |
| Kumar R, et al. 2004 [28] | PCR | 37 ulcerative colitis, 11 Crohn’s disease, 87 calcium SFs, 48 healthy controls | Prevalence of Oxalobacter: 10% in IBD, 29% in stone formers, 56% in controls. Oxalate excretion significantly higher in those without Oxalobacter colonization | The study focused on IBD-associated forms of calcium stones. |
| Kaufman DW, et al. 2008 [29] | Culture | 247 calcium SFs, 259 age-, sex- and location-matched controls | Prevalence of Oxalobacter: 17% in stone formers, 38% in controls. No association between Oxalobacter and oxalate excretion. | Absence of genomic methods of Oxalobacter detection |
| Siener R, et al. 2013 [32] | Culture + PCR | 37 calcium SFs | Prevalence of Oxalobacter: 30%. In colonized subjects, oxalate excretion is lower only under controlled dietary oxalate intake. | Study focused on oxalate metabolism; no controls enrolled. |
| Tavasoli S, et al. 2020 [30] | PCR | 29 SFs with hyperoxaluria, 29 SFs without hyperoxaluria, 29 controls | Oxalobacter more prevalent and abundant in controls and inversely related to oxaluria | Investigated also Oxalobacter abundance in feces |
| AUTHOR, YEAR [REF] | PROBIOTIC | DESIGN, PARTICIPANTS AND FOLLOW-UP DURATION | KEY FINDINGS |
|---|---|---|---|
| Campieri C, et al. 2001 [36] | Lactobacilli (L. acidophilus, L. plantarum, L. brevis) + Bifidobacterium infantis | Prospective single-arm intervention, 6 calcium stone formers, 4-week of follow-up | All participants experienced redution of urinary oxalate excretion (average 40%) |
| Duncan SH, et al. 2002 [37] | Oxalobacter formigenes strain HC1 isolated from human feces | Prospective single-arm intervention, 2 healthy volunteers, 6-h follow-up | Decrease of urinary oxalate excretion after a dietary oxalate load following the probiotic administration. |
| Lieske JC, et al. 2005 [38] | Oxadrop® (L. acidophilus, L. brevis, S. thermophilus, B. infantis) | Prospective single-arm intervention, 10 stone formers with intestinal malabsorption, 1-month follow-up | Decrease of urinary oxalate excretion shown in 7 participants over 10 (average effect size: −19%). |
| Goldfarb DS, et al. 2007 [39] | Oxadrop® (L. acidophilus, L. brevis, S. thermophilus, B. infantis) | RCT, 20 calcium oxalate stone formers with hyperoxaluria, 4-week follow-up | No significant variation of urinary oxalate excretion after treatment in both intervention and control arm. |
| Okombo J, et al. 2010 [40] | VSL#3 (L. acidophilus, L. gasseri, B. lactis) | Prospective single-arm intervention, 11 stone-free volunteers, 4-week follow-up | Reduction of fractional oxalate absorption after a dietary oxalate load (from 31% to 12%). |
| Hoppe B, et al. 2011 [41] | Oxabact® (Oxalobacter formigenes) | RCT, 42 adolescents with primary hyperoxaluria, 24-week follow-up | Reduction of urinary oxalate excretion in both intervention and control arm (average effect size 20% in both groups) |
| Al-Wahsh I, et al. 2012 [42] | VSL#3 (L. acidophilus, L. gasseri, B. lactis) | Prospective single-arm intervention, 11 healthy stone-free volunteers, 24-h follow-up | Reduction of urinary oxalate excretion after a standardized dietary oxalate load |
| Siener R, et al. 2013 [43] | Oxadrop® (L. acidophilus, L. brevis, S. thermophilus, B. infantis) | Randomized cross-over trial, 20 stone-free healthy volunteers under high-oxalate diet, 5 week follow-up | No significant variation of oxaluria detected. |
| AUTHOR, YEAR [REF] | PARTICIPANTS | STONE TYPES | COUNTRY | SAMPLES | MAIN FINDINGS IN STONE FORMERS | TAXA DEPLETED IN STONE FORMERS |
|---|---|---|---|---|---|---|
| Suryavanshi et al., 2016 [44] | 24 recurrent KSF 15 controls | Calcium oxalate | India | Feces | Gut microbiota dysbiosis with different clusterization of composition and functionality. Urinary oxalate excretion correlated with the abundance of 12 taxa. Several taxa harboring oxalate-degrading functionalities identified in both KSF and controls. | Several species, including Faecalibacterium prausnitzii |
| Stern et al., 2016 [45] | 23 KSF 6 controls | Calcium Uric acid | United States | Feces | Different microbiome composition with the prevalence of Bacteroides over Prevotella. | Prevotella |
| Tang et al., 2018 [46] | 13 multiple KSF 13 controls | Radio-opaque | China | Feces | Trend towards reduced biodiversity. Different microbiome composition clusters between KSF and controls. | Eubacterium, Dorea, Ruminiclostridium, Anaerostipes, Fusicatenibacter, Subdoligranulum, Holdemania, Dialister, Ruminococcus, Parasutterella, Bilophila |
| Ticinesi et al., 2018 [47] | 52 recurrent KSF 48 controls | Calcium | Italy | Feces | Reduced fecal microbiota biodiversity. Separate clusterization of KSF and controls. Urinary oxalate excretion correlated with the abundance of 5 taxa. Reduced representation of bacterial genes involved in oxalate degradation. Oxalate-degrading functionalities harbored in several species. | Dorea, Enterobacter, Faecalibacterium prausnitzii |
| Suryavanshi et al., 2018 [48] | 24 recurrent KSF 48 controls | Calcium oxalate | India | Feces | Dysbiosis not limited to eubacteria and also involving archaea and eukaryotes. Species able to metabolize oxalate and produce butyrate were depleted in KSF not colonized with Oxalobacter. | Several species with oxalate-metabolizing properties and butyrate producers, including Prevotella and Ruminococcus |
| Miller et al., 2019 [49] | 17 KSF 17 controls | Calcium oxalate Uric acid Struvite Cystine | Canada | Feces | KSF has reduced representation of a network of bacteria directly involved in oxalate degradation or co-occurring with Oxalobacter in network analysis. These taxa include those stimulated by oxalate intake in rodent models. | 103 bacterial taxa, including Ruminococcus and Oscillospira |
| Zampini et al., 2019 [50] | 24 KSF43 controls | Calcium Uric acid Mixed calcium+ uric acid | United States | Feces Urine Stones | Fecal microbiota similar in KSF and controls. Urinary microbiota had composition independent of stool microbiota. KSF had urinary microbiota dysbiosis correlated with antibiotic treatments, sex and family history. Stones harbor an independent microbiota population. | Lactobacillus underrepresented in urinary samples |
| Dornbier et al., 2019 [51] | 52 KSF | Any composition | United States | Urine Stones | In 20% of KSF, stone samples exhibit microbial communities with a composition independent of urine. Main components of these communities: Staphylococcus, Veillonella, Streptococcus, Enterobacter, Escherichia. | No comparison with controls provided in the study |
| NUTRITIONAL IMBALANCE | EFFECT ON URINE CHEMISTRY AND LITHOGENIC RISK | EFFECT ON GUT MICROBIOTA COMPOSITION |
|---|---|---|
| High salt intake | Increase in urine calcium Decrease in urine citrate Increased risk of CaOx lithiasis | Depletion of Lactobacillus, Akkermansia, Bifidobacterium |
| High animal protein intake | Increase in urine calcium Increase in urine uric acid Decrease in urine pH Increase of renal acid load Increased risk of CaOx and AcUr lithiasis | Depletion of Firmicutes, Bifidobacterium, Roseburia, Prevotella Increased representation of bile-tolerant bacteria Increased representation of Faecalibacterium |
| High oxalate intake | Increase in urine oxalate Mild increased risk of CaOx lithiasis | Expansion of oxalate-degrading species Increased representation of Lachnospira, Roseburia, Dialister, Faecalibacterium and Lactobacillus (probably due to other nutrients contained in oxalate-rich foods) |
| Low calcium intake | Increase in urine oxalate Increase in urine calcium from bones Increased risk of CaOx lithiasis | Reduced biodiversity with depletion of species producing SCFAs (in mice) |
| Low FAV intake | Decrease in urinary inhibitors of lithogenesis Decrease in urinary volume | Depletion of lactic acid bacteria Depletion of species producing SCFAs Depletion of Bifidobacterium, Faecalibacterium, Lactobacillus |
| Poor hydration | Decrease in urinary volume | Alterations of representation of some selected taxa, including Ruminococcaceae, Prevotella, Bifidobacteriaceae and Oxalobacteriaceae depending on the pH of ingested fluids |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ticinesi, A.; Nouvenne, A.; Chiussi, G.; Castaldo, G.; Guerra, A.; Meschi, T. Calcium Oxalate Nephrolithiasis and Gut Microbiota: Not just a Gut-Kidney Axis. A Nutritional Perspective. Nutrients 2020, 12, 548. https://doi.org/10.3390/nu12020548
Ticinesi A, Nouvenne A, Chiussi G, Castaldo G, Guerra A, Meschi T. Calcium Oxalate Nephrolithiasis and Gut Microbiota: Not just a Gut-Kidney Axis. A Nutritional Perspective. Nutrients. 2020; 12(2):548. https://doi.org/10.3390/nu12020548
Chicago/Turabian StyleTicinesi, Andrea, Antonio Nouvenne, Giulia Chiussi, Giampiero Castaldo, Angela Guerra, and Tiziana Meschi. 2020. "Calcium Oxalate Nephrolithiasis and Gut Microbiota: Not just a Gut-Kidney Axis. A Nutritional Perspective" Nutrients 12, no. 2: 548. https://doi.org/10.3390/nu12020548
APA StyleTicinesi, A., Nouvenne, A., Chiussi, G., Castaldo, G., Guerra, A., & Meschi, T. (2020). Calcium Oxalate Nephrolithiasis and Gut Microbiota: Not just a Gut-Kidney Axis. A Nutritional Perspective. Nutrients, 12(2), 548. https://doi.org/10.3390/nu12020548