Understanding the Fate of Almond (Prunus dulcis (Mill.) D.A. Webb) Oleosomes during Simulated Digestion

 ,
,  ,
, 

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
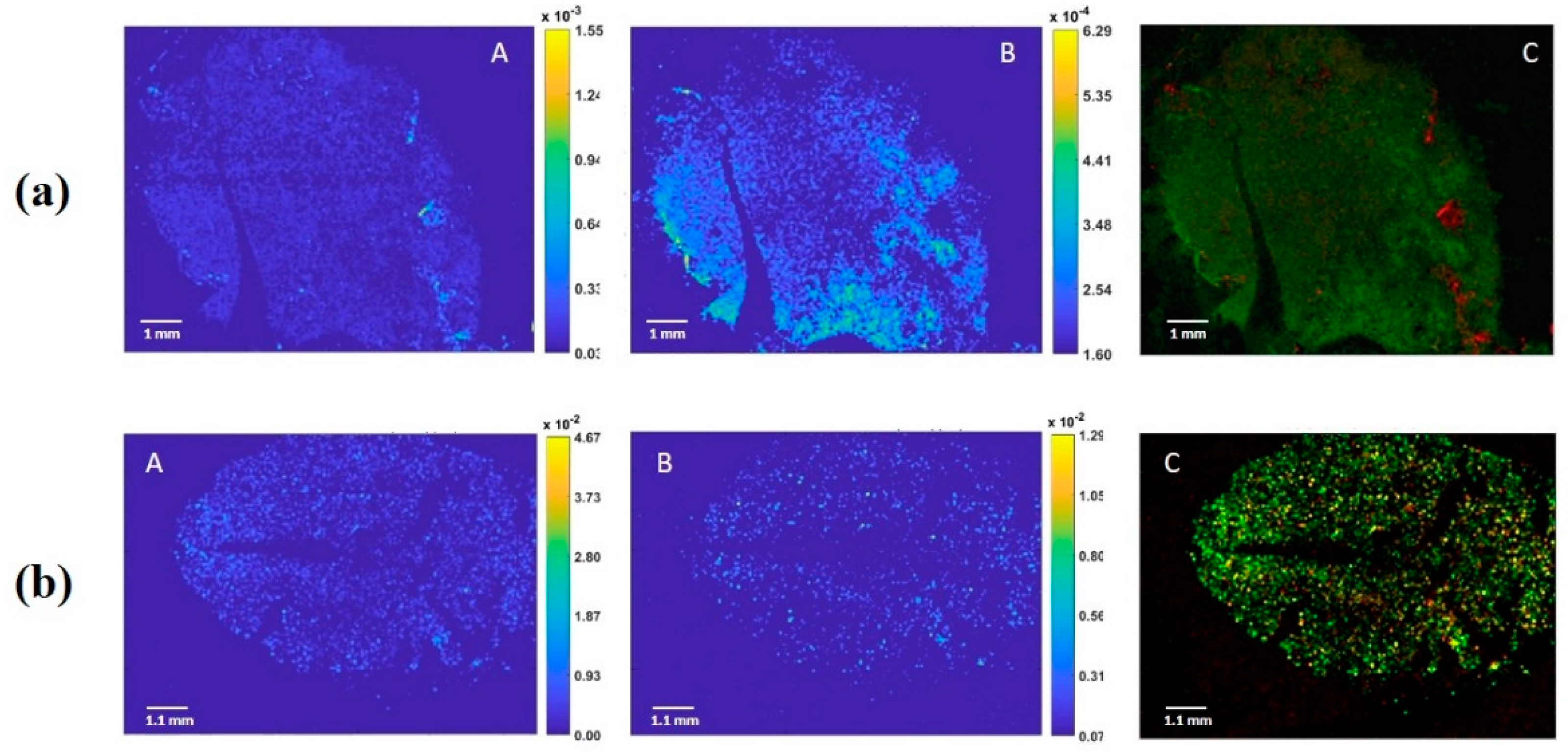
2.1. Imaging Mass Spectrometry Analysis
2.2. Extraction of Almond Oleosomes
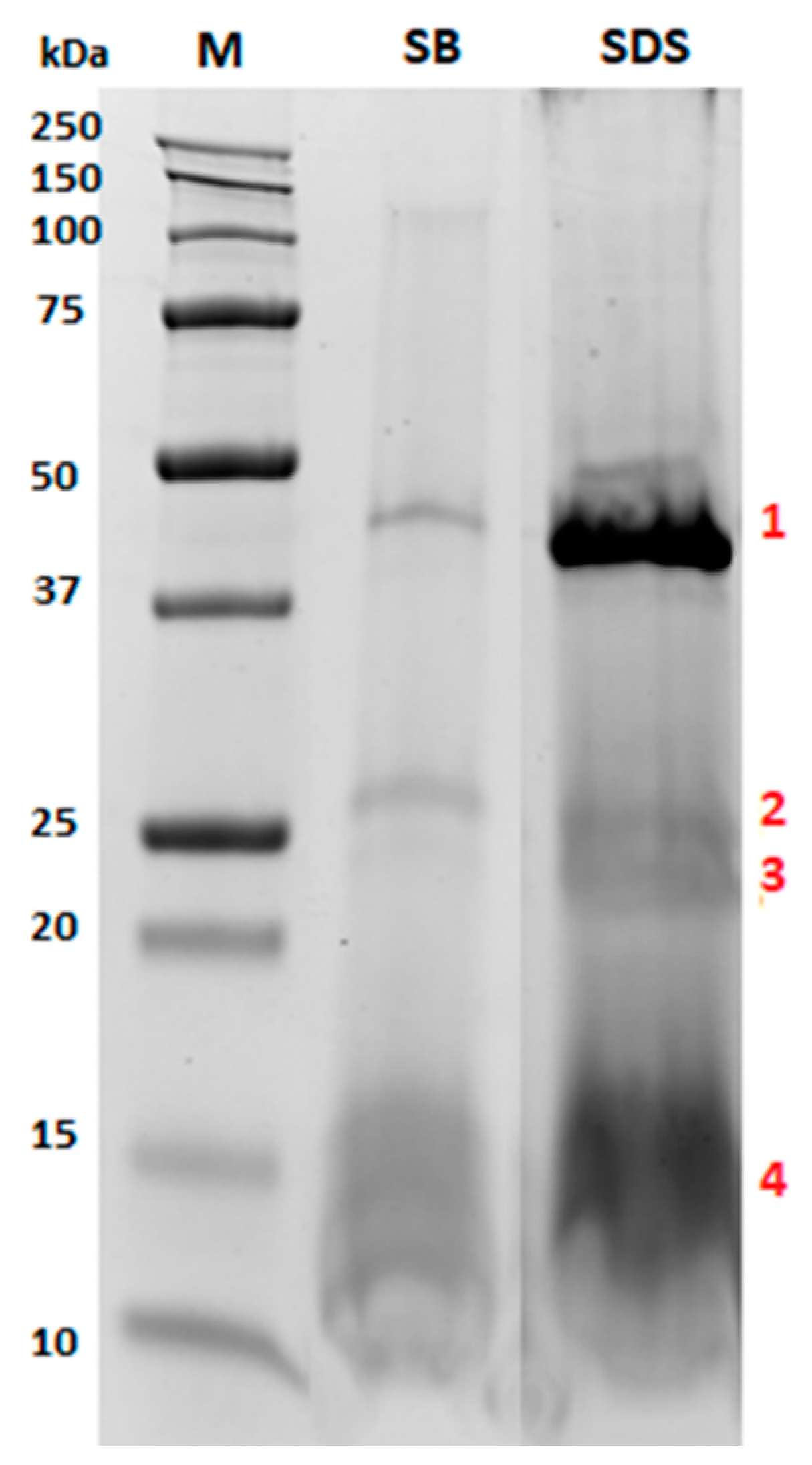
2.3. Characterization of the Protein Fraction Embedding in Phospholipid Layer
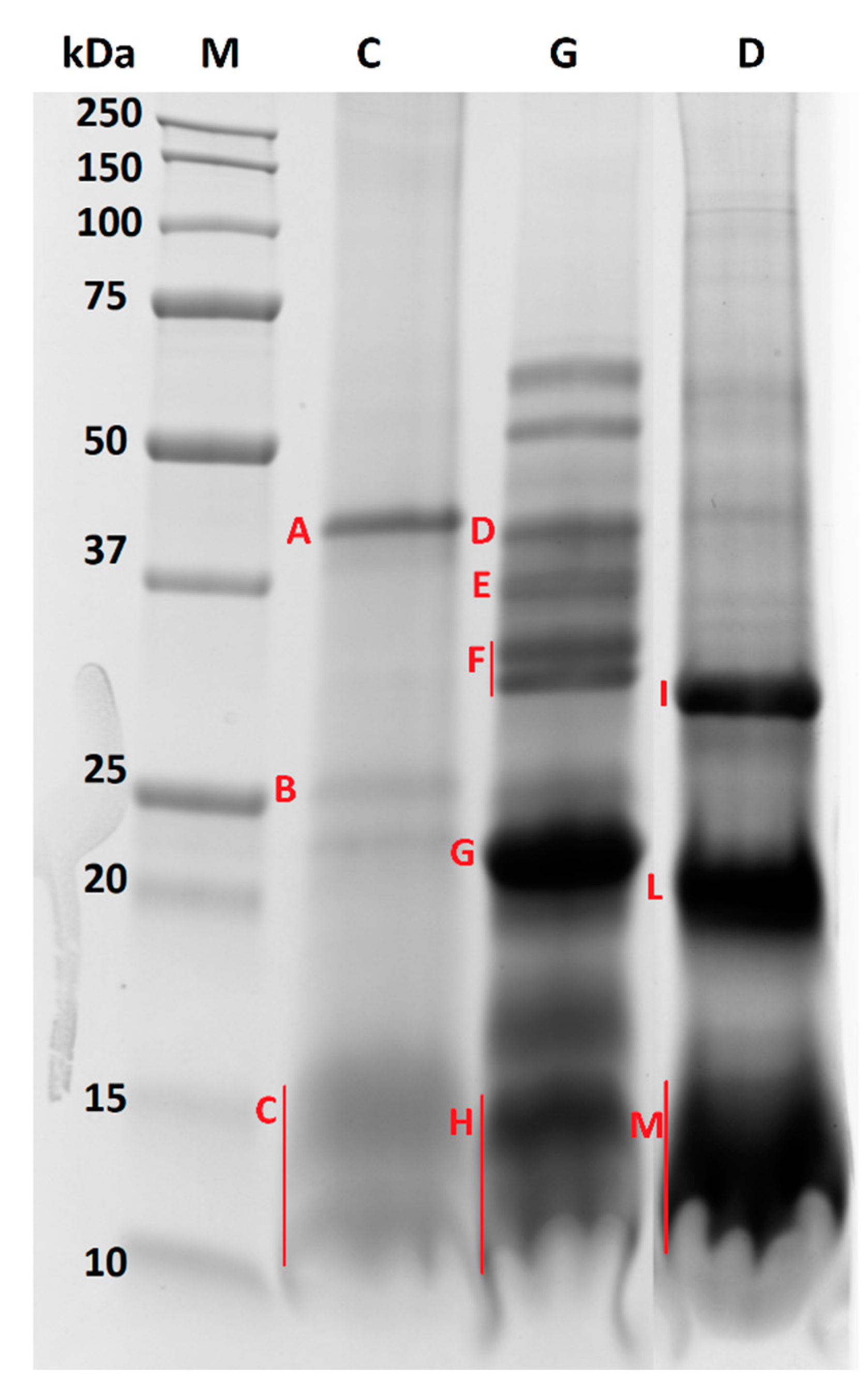
2.4. In-Gel Tryptic Digestion
2.5. Liquid Chromatography High-Resolution Tandem Mass Spectrometry (LC-HR-MS/MS) Analysis and Software-Based Identification
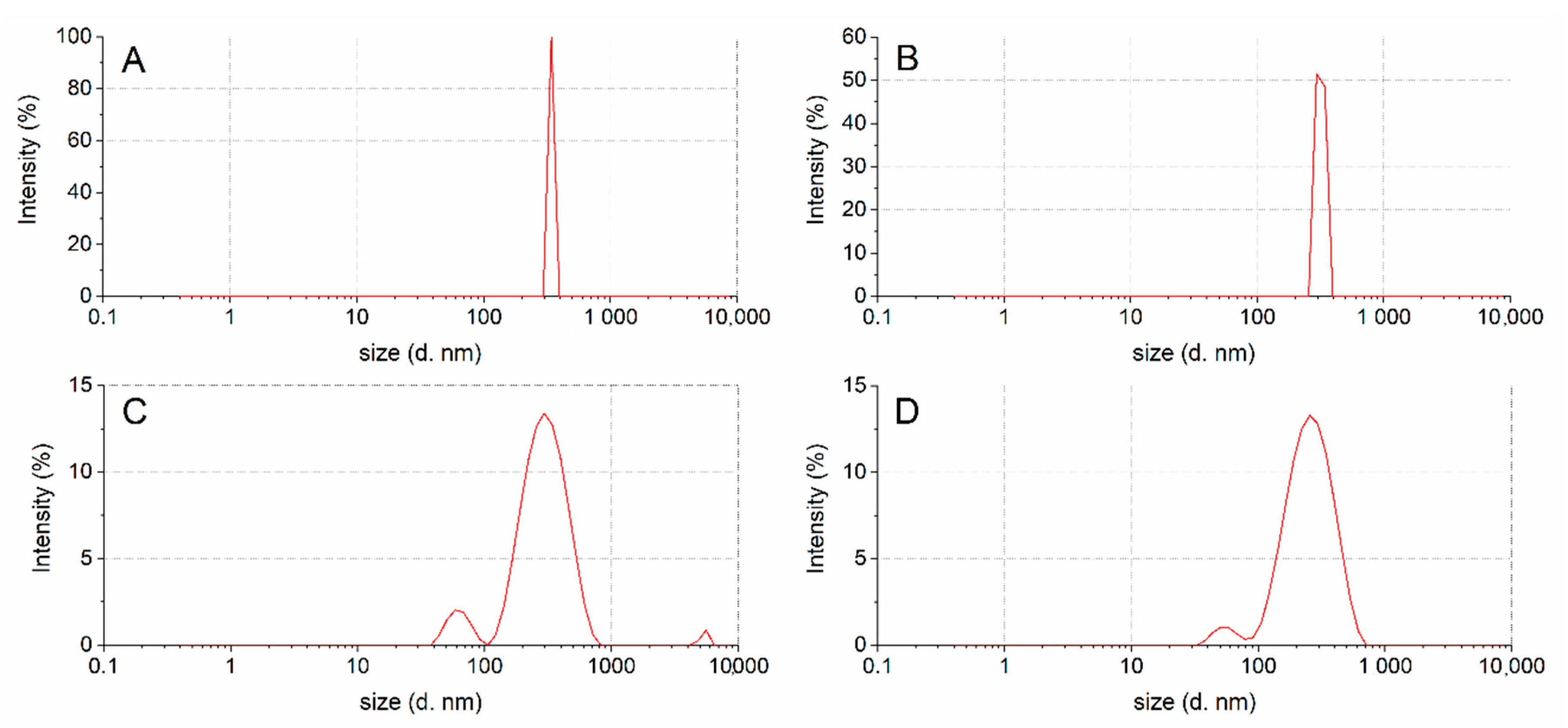
2.6. Droplet Size Distribution
2.7. Simulated Human Digestion
2.7.1. Chemicals and Enzymes
2.7.2. Chewing
2.7.3. Gastric Digestion
2.7.4. Duodenal Digestion
2.8. Post Digestion Analyses
3. Results
3.1. Oleosomes Distribution within Almond Seeds
3.2. Protein Profile
3.3. Droplet Size Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, A.H.C. Structure of plant seed oil bodies. Curr. Opin. Struct. Biol. 1994, 4, 493–498. [Google Scholar] [CrossRef]
- Nikiforidis, C.V. Structure and functions of oleosomes (oil bodies). Adv. Colloid Interface Sci. 2019, 274, 102039. [Google Scholar] [CrossRef]
- Beisson, F.; Ferte, N.; Bruley, S.; Voultoury, R.; Verger, R.; Arondel, V. Oil-bodies as substrates for lipolytic enzymes. Biochim. Biophys. 2001, 1531, 47–58. [Google Scholar] [CrossRef]
- Mandalari, G.; Grundy, M.M.; Grassby, T.; Parker, M.L.; Cross, K.L.; Chessa, S.; Bisignano, C.; Barreca, D.; Bellocco, E.; Laganà, G.; et al. The effects of processing and mastication on almond lipid bioaccessibility using novel methods of in vitro digestion modelling and micro-structural analysis. Br. J. Nutr. 2014, 112, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Parker, M.L.; Grundy, M.M.; Grassby, T.; Smeriglio, A.; Bisignano, C.; Raciti, R.; Trombetta, D.; Baer, D.J.; Wilde, P.J. Understanding the effect of particle size and processing on almond lipid bioaccessibility through microstructural analysis: From mastication to faecal collection. Nutrients 2018, 10, 213. [Google Scholar] [CrossRef] [PubMed]
- Novotny, J.A.; Gebauer, S.K.; Baer, D.J. Discrepancy between the Atwater factor predicted and empirically measured energy values of almonds in human diets. Am. J. Clin. Nutr. 2012, 96, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.R.; Kendall, C.W.; Ren, Y.; Parker, C.; Pacy, J.F.; Waldron, K.W.; Jenkins, D.J. Role of cell walls in the bioaccessibility of lipids in almond seeds. Am. J. Clin. Nutr. 2004, 80, 604–613. [Google Scholar] [CrossRef]
- Grundy, M.M.; Grassby, T.; Mandalari, G.; Waldron, K.W.; Butterworth, P.J.; Berry, S.E.; Ellis, P.R. Effect of mastication on lipid bioaccessibility of almonds in a randomized human study and its implications for digestion kinetics, metabolizable energy, and postprandial lipemia. Am. J. Clin. Nutr. 2015, 101, 25–33. [Google Scholar] [CrossRef]
- Berry, S.E.; Tydeman, E.A.; Lewis, H.B.; Phalora, R.; Rosborough, J.; Picout, D.R.; Ellis, P.R. Manipulation of lipid bioaccessibility of almond seeds influences postprandial lipemia in healthy human subjects. Am. J. Clin. Nutr. 2008, 88, 922–929. [Google Scholar] [CrossRef]
- Grundy, M.M.L.; Carrière, F.; Mackie, A.R.; Gray, D.A.; Butterworth, P.J.; Ellis, P.R. The role of plant cell wall encapsulation and porosity in regulating lipolysis during the digestion of almond seeds. Food Funct. 2016, 7, 69–78. [Google Scholar] [CrossRef]
- Barreca, D.; Nabavi, S.M.; Sureda, A.; Rasekhian, M.; Raciti, R.; Silva, A.S.; Annunziata, G.; Arnone, A.; Tenore, G.C.; Süntar, İ.; et al. Almonds (Prunus Dulcis Mill. D. A. Webb): A Source of Nutrients and Health-Promoting Compounds. Nutrients 2020, 12, 672. [Google Scholar] [CrossRef] [PubMed]
- Sturtevant, D.; Lee, Y.J.; Chapman, K.D. Matrix assisted laser desorption/ionization-mass spectrometry imaging (MALDI-MSI) for direct visualization of plant metabolites in situ. Curr. Opin. Biotechnol. 2016, 37, 53–60. [Google Scholar] [CrossRef]
- Bouschen, W.; Schulz, O.; Eikel, D.; Spengler, B. Matrix vapor deposition/recrystallization and dedicated spray preparation for high-resolution scanning microprobe matrix-assisted laser desorption/ionization imaging mass spectrometry (SMALDI-MS) of tissue and single cells. Rap. Comm. Mass Spec. 2010, 24, 355–364. [Google Scholar] [CrossRef]
- Guenther, S.; Koestler, M.; Schulz, O.; Spengler, B. Laser spot size and laser power dependence of ion formation in high resolution MALDI imaging. Int. J. Mass Spec. 2010, 294, 7–15. [Google Scholar] [CrossRef]
- De Chirico, S.; di Bari, V.; Foster, T.; Gray, D. Enhancing the recovery of oilseed rape seed oil bodies (oleosomes) using bicarbonate-based soaking and grinding media. Food Chem. 2018, 241, 419–426. [Google Scholar] [CrossRef]
- De Angelis, E.; Pilolli, R.; Bavaro, S.L.; Monaci, L. Insight into the gastro-duodenal digestion resistance of soybean proteins and potential implications for residual immunogenicity. Food Funct. 2017, 8, 1599–1610. [Google Scholar] [CrossRef]
- De Angelis, E.; Bavaro, S.L.; Forte, G.; Pilolli, G.; Monaci, L. Heat and Pressure Treatments on Almond Protein Stability and Change in Immunoreactivity after Simulated Human Digestion. Nutrients 2018, 10, 1679. [Google Scholar] [CrossRef]
- Bavaro, S.L.; Di Stasio, L.; Mamone, G.; De Angelis, E.; Nocerino, R.; Canani, R.B.; Logrieco, A.F.; Montemurro, N.; Monaci, L. Effect of thermal/pressure processing and simulated human digestion on the immunoreactivity of extractable peanut allergens. Food Res. Int. 2018, 109, 126–137. [Google Scholar] [CrossRef]
- Monaci, L.; Pilolli, R.; De Angelis, E.; Godula, M.; Visconti, A. Multi-allergen detection in food by micro high-performance liquid chromatography coupled to a dual cell linear ion trap mass spectrometry. J. Chromat. A 2014, 1358, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Pilolli, R.; De Angelis, E.; Monaci, L. Streamlining the analytical workflow for multiplex MS/MS allergen detection in processed foods. Food Chem. 2017, 221, 1747–1753. [Google Scholar] [CrossRef]
- Bernardi, D.; Pereira, S.; Maciel, T.A.; Bortoloto, J.; Viera, G.S.; Oliveira, G.C.; Rocha-Filho, P.A. Formation and stability of oil-in-water nanoemulsions containing rice bran oil: In vitro and in vivo assessments. J. Nanobiotech. 2011, 9, 44. [Google Scholar] [CrossRef]
- Shen, Q.; Dong, W.; Yang, M.; Li, L.; Cheung, H.Y.; Zhang, Z. Lipidomic fingerprint of almonds (Prunus dulcis L. cv Nonpareil) using TiO₂ nanoparticle based matrix solid-phase dispersion and MALDI-TOF/MS and its potential in geographical origin verification. J. Agric. Food Chem. 2013, 61, 7739–7748. [Google Scholar] [CrossRef]
- ISO. International Standard ISO 22412:2017. Particle Size Analysis-Dynamic Light Scattering (DLS); International Organisation for Standardisation (ISO): Geneva, Switzerland, 2017. [Google Scholar]
- Liu, Z.; Wang, W.; Huang, G.; Zhang, W.; Ni, L. In vitro and in vivo evaluation of the prebiotic effect of raw and roasted almonds (Prunus amygdalus). J. Sci. Food Agric. 2016, 96, 1836–1843. [Google Scholar] [CrossRef]
- Hollingworth, S.; Dalton, M.; Blundell, J.; Finlayson, G. Evaluation of the Influence of Raw Almonds on Appetite Control: Satiation, Satiety, Hedonics and Consumer Perceptions. Nutrients 2019, 11, 2030. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, M.; Teuber, S.S.; Roux, K.H.; Sathe, S.K. Effects of 478 roasting, blanching, autoclaving, and microwave heating on antigenicity of almond 479 (Prunus dulcis L.) proteins. J. Agric. Food Chem. 2002, 50, 3544–3548. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band | Accession | Description (OS Prunus dulcis) | Coverage (%) | Peptides (Unique) | PSMs | MW (kDa) | Score |
|---|---|---|---|---|---|---|---|
| 1 | A0A5E4EAH6 | PREDICTED: basic 7S globulin | 58.7 | 21 (21) | 107 | 46.9 | 70.2 |
| 2 | A0A5E4EKE0 | PREDICTED: peroxygenase | 15.9 | 3 (2) | 13 | 26.5 | 5.9 |
| A0A4Y1QWY8 | Beta-tonoplast intrinsic protein | 8.9 | 2 (2) | 8 | 27.2 | 2.7 | |
| 3 | A0A5E4FFS0 | PREDICTED: legumin | 8.5 | 4 (4) | 14 | 63 | 5.6 |
| A0A4Y1QWY8 | Beta-tonoplast intrinsic protein | 23.4 | 4 (4) | 15 | 27.2 | 4.4 | |
| 4 | A0A5E4ET55 | Oleosin | 50.6 | 5 (4) | 27 | 16.6 | 9.5 |
| A0A5E4FFS0 | PREDICTED: legumin | 5.6 | 2 (2) | 4 | 63 | 1.3 | |
| A0A5E4EAT1 | Oleosin | 52.0 | 4 (4) | 16 | 15.6 | 4.1 |
| Digestion Step | Band | Accession | Description (OS = Prunus dulcis) | Coverage (%) | Peptides (Unique) | PSMs | MW (kDa) | Score |
|---|---|---|---|---|---|---|---|---|
| Chew | A | A0A5E4EAH6 | PREDICTED: basic 7S globulin. OS = Prunus dulcis. | 77.0 | 43 (42) | 230 | 46.9 | 88.8 |
| P00766 | Chymotrypsinogen A | 30.6 | 2 (1) | 20 | 25.7 | 5.2 | ||
| B | A0A5E4EKE0 | PREDICTED: peroxygenase OS = Prunus dulcis. | 59.0 | 16 (13) | 75 | 26.5 | 36.5 | |
| A0A5E4ET55 | Oleosin OS = Prunus dulcis | 23.7 | 2 (2) | 6 | 16.6 | 1.3 | ||
| A0A5E4FV72 | PREDICTED: vicilin OS = Prunus dulcis. | 6.4 | 2 (2) | 8 | 60 | 9.7 | ||
| C | A0A5E4ET55 | Oleosin OS = Prunus dulcis | 57.0 | 5 (4) | 27 | 16.6 | 18.0 | |
| A0A5E4EAT1 | Oleosin OS = Prunus dulcis | 43.2 | 4 (4) | 22 | 15.6 | 6.3 | ||
| P00766 | Chymotrypsinogen A | 29.0 | 5 (3) | 16 | 25.7 | 10.6 | ||
| A0A5E4EYX0 | PREDICTED: vicilin antimicrobial peptides 2-2 OS = Prunus dulcis. | 19.7 | 5 (5) | 19 | 31.1 | 7.9 | ||
| A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 19.2 | 3 (3) | 17 | 63 | 1.4 | ||
| Gastric | D | A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 40.3 | 16 (16) | 75 | 63 | 24.5 |
| P00766 | Chymotrypsinogen A OS = Bos Taurus | 24.5 | 2 (2) | 6 | 25.7 | 3.7 | ||
| E | A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 54.4 | 21 (21) | 144 | 63 | 76.6 | |
| F | A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 68.6 | 43 (39) | 272 | 63 | 156.8 | |
| A0A5E4FK23 | PREDICTED: legumin OS = Prunus dulcis. | 25.5 | 4 (2) | 25 | 35.9 | 7.8 | ||
| G | A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 76.4 | 63 (59) | 408 | 63 | 272.5 | |
| A0A5E4FK23 | PREDICTED: legumin OS = Prunus dulcis. | 44.5 | 8 (4) | 84 | 35.9 | 36.2 | ||
| A0A5E4FV72 | PREDICTED: vicilin OS = Prunus dulcis. | 18.5 | 3 (3) | 19 | 60 | 5.7 | ||
| A0A5E4F5V3 | PREDICTED: putative disease OS = Prunus dulcis. | 2.7 | 2 (1) | 17 | 128.4 | 2.6 | ||
| A0A5E4G4H3 | PREDICTED: STRUBBELIG-RECEPTOR FAMILY OS = Prunus dulcis. | 4.1 | 2 (1) | 6 | 80.4 | 1.3 | ||
| A0A5E4G2W8 | PREDICTED: TCM_014128 isoform OS = Prunus dulcis | 1.0 | 2 (2) | 5 | 119.9 | 3.1 | ||
| A0A5E4FDL3 | PREDICTED: ferric OS = Prunus dulcis. | 4.3 | 2 (1) | 10 | 74 | 1.5 | ||
| A0A5E4ETP4 | PREDICTED: kinesin OS = Prunus dulcis. | 2.4 | 2 (1) | 9 | 158.6 | 1.3 | ||
| P00766 | Chymotrypsinogen A OS = Bos Taurus | 26.9 | 4 (4) | 13 | 25.7 | 3.7 | ||
| A0A5E4ERF0 | WAT1-related protein OS = Prunus dulcis. | 5.0 | 2 (1) | 6 | 39.3 | 1.3 | ||
| H | A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 60.0 | 29 (29) | 180 | 63 | 75.9 | |
| A0A5E4EAT1 | Oleosin OS = Prunus dulcis | 44.6 | 3 (3) | 11 | 15.6 | 7.4 | ||
| Duodenal | I | A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 47.6 | 58 (56) | 366 | 63 | 268.1 |
| A0A5E4FK23 | PREDICTED: legumin OS = Prunus dulcis. | 41.6 | 13 (12) | 88 | 35.9 | 44.5 | ||
| A0A5E4FDR1 | PREDICTED: legumin OS = Prunus dulcis. | 22.4 | 3 (2) | 25 | 20.5 | 9.9 | ||
| A0A5E4F2T7 | PREDICTED: vicilin OS = Prunus dulcis. | 14.8 | 3 (3) | 16 | 62.1 | 6.5 | ||
| A0A5E4FH19 | PREDICTED: SMG7 OS = Prunus dulcis. | 4.0 | 2 (1) | 18 | 126.2 | 2.9 | ||
| A0A5E4GE42 | Uncharacterized protein OS = Prunus dulcis | 54.7 | 15 (12) | 74 | 34 | 47.7 | ||
| P00766 | Chymotrypsinogen A OS = Bos Taurus | 31.0 | 5 (2) | 17 | 25.7 | 8.5 | ||
| L | A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 37.0 | 50 (47) | 318 | 63 | 253.7 | |
| A0A5E4FK23 | PREDICTED: legumin OS = Prunus dulcis. | 36.6 | 9 (7) | 59 | 35.9 | 32.5 | ||
| A0A5E4FV72 | PREDICTED: vicilin OS = Prunus dulcis. | 27.5 | 4 (4) | 30 | 60 | 4.7 | ||
| A0A5E4EZP4 | PREDICTED: vicilin OS = Prunus dulcis. | 11.3 | 3 (3) | 15 | 93.7 | 3.2 | ||
| A0A5E4F3E3 | DNA-directed RNA polymerase subunit beta. | 2.3 | 2 (1) | 3 | 129.8 | 2.1 | ||
| M | P00766 | Chymotrypsinogen A OS = Bos taurus | 60.8 | 13 (8) | 51 | 25.7 | 34.9 | |
| A0A5E4FFS0 | PREDICTED: legumin OS = Prunus dulcis. | 48.8 | 22 (18) | 154 | 63 | 74.8 | ||
| A0A5E4EAT1 | Oleosin OS = Prunus dulcis | 35.1 | 5 (4) | 21 | 15.6 | 8.4 | ||
| P17538 | Chymotrypsinogen B OS = Homo sapiens | 25.8 | 5 (1) | 27 | 27.7 | 17.2 | ||
| A0A5E4FK23 | PREDICTED: legumin OS = Prunus dulcis. | 19.9 | 2 (2) | 24 | 35.9 | 5.3 | ||
| A0A5E4FDR1 | PREDICTED: legumin OS = Prunus dulcis. | 14.7 | 3 (1) | 15 | 20.5 | 4.9 | ||
| A0A5E4EU98 | WAT1-related protein OS = Prunus dulcis. | 3.0 | 2 (1) | 4 | 40.7 | 3.2 | ||
| A0A5E4EZP4 | PREDICTED: vicilin OS = Prunus dulcis. | 8.1 | 3 (1) | 14 | 93.7 | 1.3 |
| Sample | DH (nm ± SD) (%) b | Polydispersity Index |
|---|---|---|
| Untreated | 343 ± 12 (100%) | 0.07 |
| Chewed | 318 ± 23 (100%) | 0.14 |
| Gastric | 308 ± 57 (91%) 77 ± 19 (9%) | 0.37 ≤0.4 |
| Duodenal | 246 ± 47 (96%) 56 ± 11 (4%) | 0.38 0.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trombetta, D.; Smeriglio, A.; Denaro, M.; Zagami, R.; Tomassetti, M.; Pilolli, R.; De Angelis, E.; Monaci, L.; Mandalari, G. Understanding the Fate of Almond (Prunus dulcis (Mill.) D.A. Webb) Oleosomes during Simulated Digestion. Nutrients 2020, 12, 3397. https://doi.org/10.3390/nu12113397
Trombetta D, Smeriglio A, Denaro M, Zagami R, Tomassetti M, Pilolli R, De Angelis E, Monaci L, Mandalari G. Understanding the Fate of Almond (Prunus dulcis (Mill.) D.A. Webb) Oleosomes during Simulated Digestion. Nutrients. 2020; 12(11):3397. https://doi.org/10.3390/nu12113397
Chicago/Turabian StyleTrombetta, Domenico, Antonella Smeriglio, Marcella Denaro, Roberto Zagami, Mara Tomassetti, Rosa Pilolli, Elisabetta De Angelis, Linda Monaci, and Giuseppina Mandalari. 2020. "Understanding the Fate of Almond (Prunus dulcis (Mill.) D.A. Webb) Oleosomes during Simulated Digestion" Nutrients 12, no. 11: 3397. https://doi.org/10.3390/nu12113397
APA StyleTrombetta, D., Smeriglio, A., Denaro, M., Zagami, R., Tomassetti, M., Pilolli, R., De Angelis, E., Monaci, L., & Mandalari, G. (2020). Understanding the Fate of Almond (Prunus dulcis (Mill.) D.A. Webb) Oleosomes during Simulated Digestion. Nutrients, 12(11), 3397. https://doi.org/10.3390/nu12113397