Wolffia globosa–Mankai Plant-Based Protein Contains Bioactive Vitamin B12 and Is Well Absorbed in Humans

 , , ,
, , ,  ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Mankai Laboratory Analyses
2.1.1. Plant Sources
Vitamin B12 Detection in Plants Cultivated Under Greenhouse Conditions
Vitamin B12 Detection in Axenic Culture
2.1.2. Vitamin B12 Analyses
Bioassay Method
Liquid Chromatography/Electrospray Ionization Tandem Mass Spectrometry (ESI LC-MS/MS) Assay
2.2. The DIRECT PLUS Dietary Intervention Trial
2.2.1. Study Design
2.2.2. Randomization and Intervention
2.2.3. Outcomes
2.2.4. Statistical Analysis
2.3. Anoxic Gut Microbiome Bioreactors Pilot Experiment
2.3.1. Microbiota Reactors (Human Fecal Mixture)
2.3.2. Media, Anoxic Bioreactor, Mankai Lysate, and Sampling
2.3.3. Chemical and Molecular Analysis
3. Results
3.1. Mankai Plant Analyses
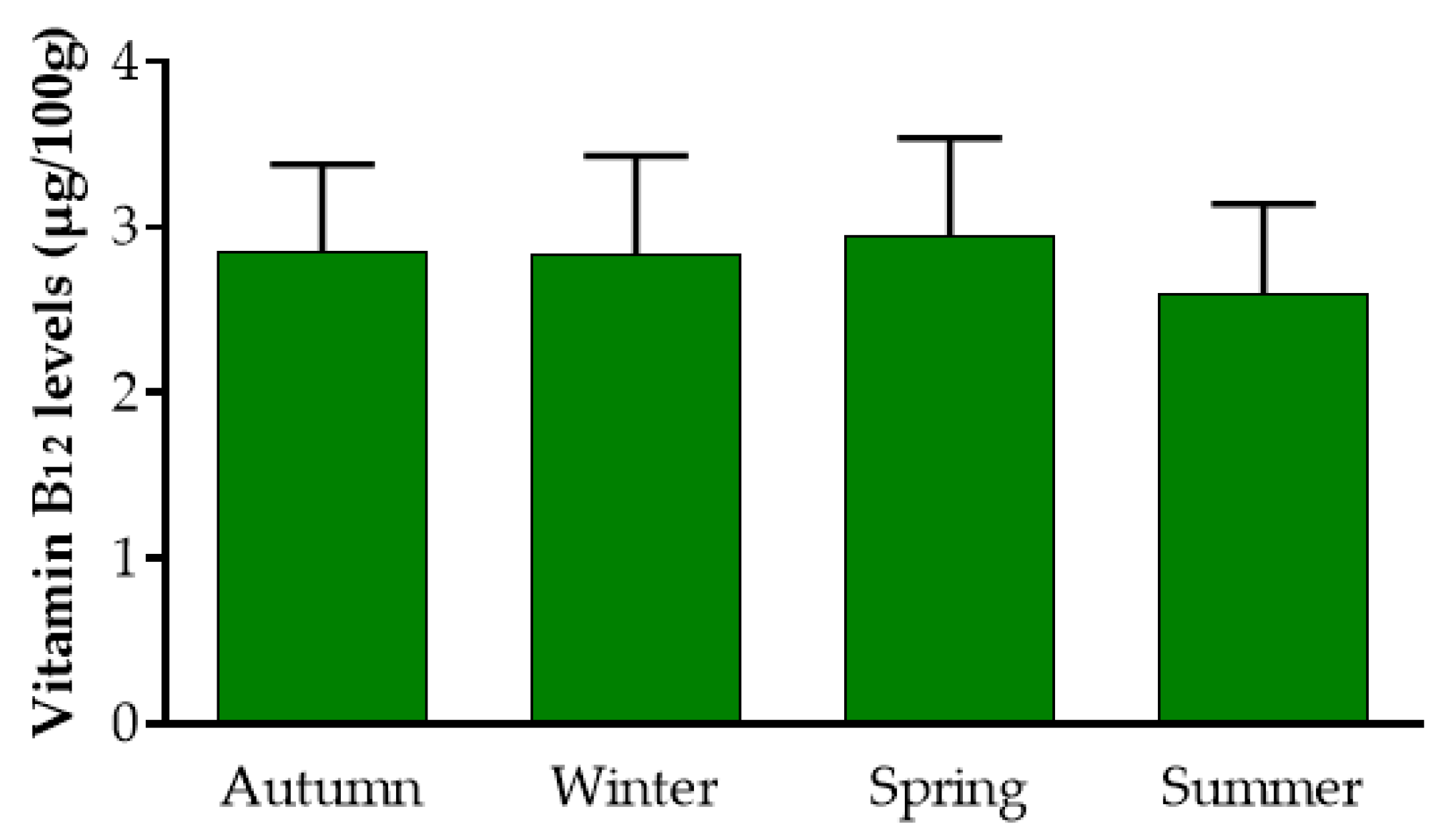
3.1.1. Content and Stability of Vitamin B12 Levels during Different Seasons
3.1.2. Inherent Presence of Vitamin B12 in Mankai Axenic Cultures
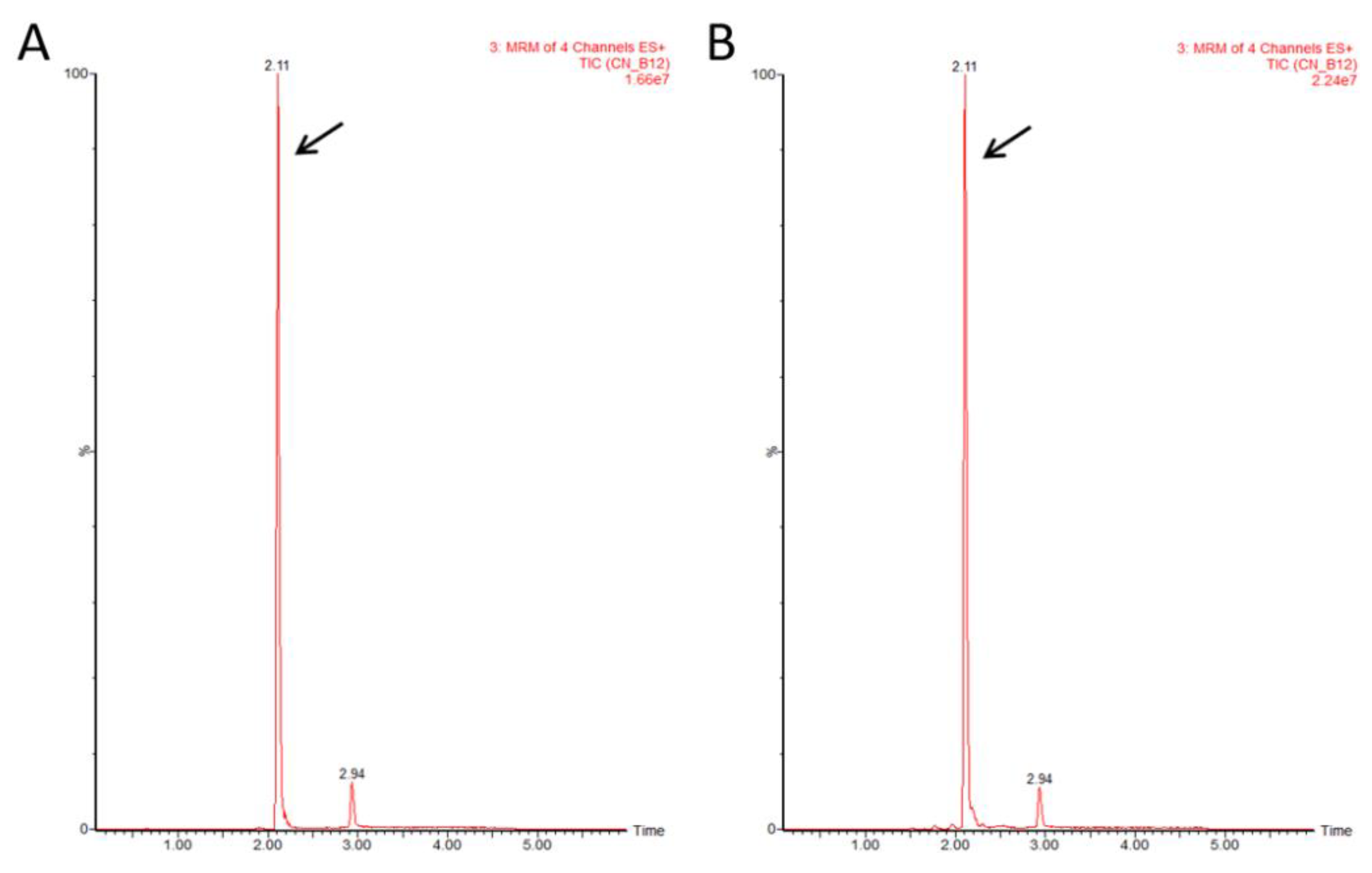
3.1.3. Identification of Vitamin B12 Purified from Mankai
3.1.4. Quantification of Total Vitamin B12 Purified from Mankai
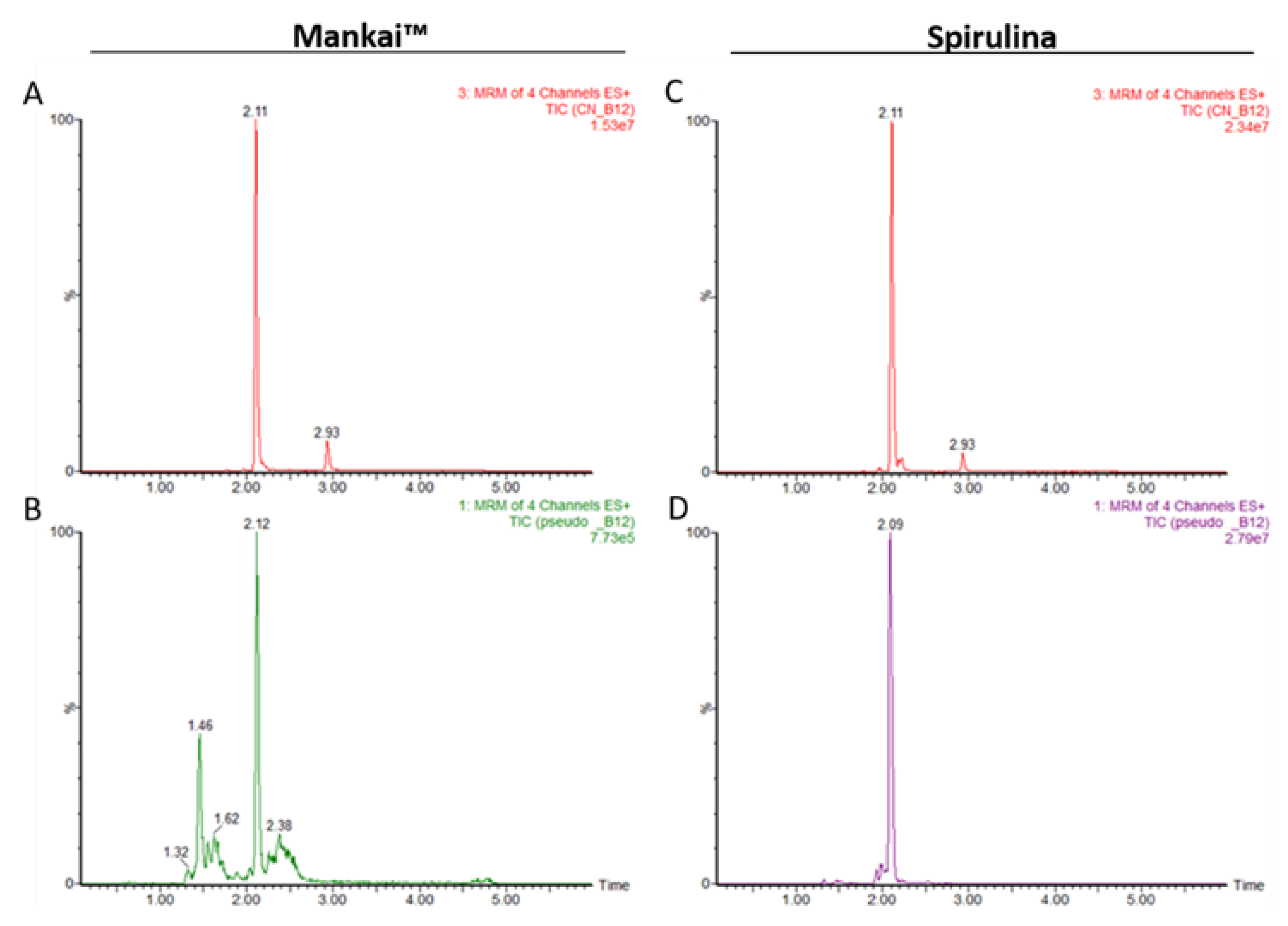
3.1.5. Authentic CN-B12 and Pseudo CN-B12 in Mankai
3.2. DIRECT PLUS Trial
3.2.1. Baseline Characteristics
3.2.2. The Effect of the Intervention on Serum B12 Levels
3.2.3. Changes in chemical and hematological Parameters
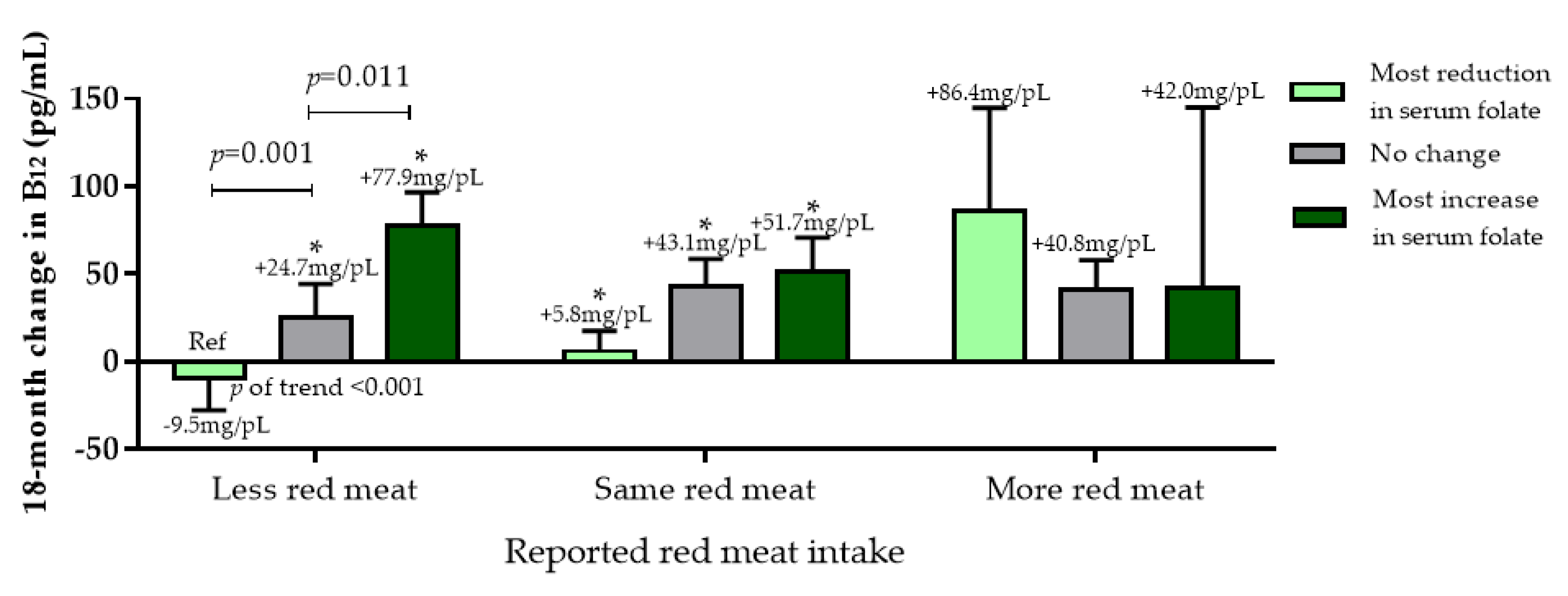
3.2.4. Dietary Vitamin B12 Sources
3.3. Anoxic Bioreactors Pilot Experiment
Predicted Functional Pathways-Gut Bioreactor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Watanabe, F.; Bito, T. Determination of cobalamin and related compounds in foods. J. AOAC Int. 2018, 101, 1308–1313. [Google Scholar] [CrossRef] [PubMed]
- Burgess, C.M.; Smid, E.J.; van Sinderen, D. Bacterial vitamin B2, B11 and B12 overproduction: An overview. Int. J. Food Microbiol. 2009, 133, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Green, R. Vitamin B12 deficiency from the perspective of a practicing hematologist. Blood J. Am. Soc. Hematol. 2017, 129, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, F.; Takenaka, S.; Kittaka-Katsura, H.; Ebara, S.; Miyamoto, E. Characterization and bioavailability of vitamin B12-compounds from edible algae. J. Nutr. Sci. Vitaminol. 2002, 48, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Green, R.; Miller, J.W. Vitamin B12. In Handbook of Vitamins; Zempleni, J., Suttie, J.W., Gregory, J.F., III, Stover, P.J., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 447–489. [Google Scholar]
- Rizzo, G.; Laganà, A.S.; Rapisarda, A.M.C.; Ferrera, L.; Grazia, G.M.; Buscema, M.; Rossetti, P.; Nigro, A.; Muscia, V.; Valenti, G.; et al. Vitamin B12 among vegetarians: Status, assessment and supplementation. Nutrients 2016, 8, 767. [Google Scholar] [CrossRef] [PubMed]
- Crowe, F.L.; Appleby, P.N.; Travis, R.C.; Key, T.J. Risk of hospitalization or death from ischemic heart disease among British vegetarians and nonvegetarians: Results from the EPIC-Oxford cohort study. Am. J. Clin. Nutr. 2013, 97, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Barnard, N.D.; Levin, S.M.; Watanabe, M. Vegetarian diets and glycemic control in diabetes: A systematic review and meta-analysis. Cardiovasc. Diagn. Ther. 2014, 4, 373. [Google Scholar]
- Wang, F.; Zheng, J.; Yang, B.; Jiang, J.; Fu, Y.; Li, D. Effects of vegetarian diets on blood lipids: A systematic review and meta-analysis of randomized controlled trials. J. Am. Heart Assoc. 2015, 4, e002408. [Google Scholar] [CrossRef]
- Yokoyama, Y.; Nishimura, K.; Barnard, N.D.; Miyamoto, Y. Blood pressure and vegetarian diets. In Vegetarian and Plant-Based Diets in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2017; pp. 395–413. [Google Scholar]
- Sukumar, N.; Saravanan, P. Investigating vitamin B12 deficiency. BMJ 2019, 365, l1865. [Google Scholar] [CrossRef]
- Gilbert, S.; Xu, J.; Acosta, K.; Poulev, A.; Lebeis, S.; Lam, E. Bacterial production of indole related compounds reveals their role in association between duckweeds and endophytes. Front. Chem. 2018, 6, 265. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Sree, K.S.; Bog, M.; Ecker, J.; Boehm, V.; Lorkowski, S.; Sommer, K.; Vetter, W.; Tolzin-Banasch, K.; Kirmse, R.; et al. Nutritional value of the duckweed species of the genus Wolffia (Lemnaceae) as human food. Front. Chem. 2018, 6, 483. [Google Scholar] [CrossRef] [PubMed]
- Kawamata, Y.; Shibui, Y.; Takumi, A.; Seki, T.; Shimada, T.; Hashimoto, M.; Inoue, N.; Kobayashi, H.; Narita, T. Genotoxicity and repeated-dose toxicity evaluation of dried Wolffia globosa Mankai. Toxicol. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.; Zelicha, H.; Tsaban, G.; Yaskolka Meir, A.; Rinott, E.; Kovsan, J.; Novack, L.; Thiery, J.; Ceglarek, U.; Burkhardt, R.; et al. Protein bioavailability of Wolffia globosa duckweed, a novel aquatic plant–A randomized controlled trial. Clin. Nutr. 2019, 38. [Google Scholar] [CrossRef]
- Yan, Y.; Candreva, J.; Shi, H.; Ernst, E.; Martienssen, R.; Schwender, J.; Shanklin, J. Survey of the total fatty acid and triacylglycerol composition and content of 30 duckweed species and cloning of a Δ6-desaturase responsible for the production of γ-linolenic and stearidonic acids in Lemna gibba. BMC Plant Biol. 2013, 13, 201. [Google Scholar] [CrossRef]
- Yaskolka Meir, A.; Tsaban, G.; Zelicha, H.; Rinott, E.; Kaplan, A.; Youngster, I.; Rudich, A.; Shelef, I.; Tirosh, A.; Brikner, D.; et al. A Green-mediterranean diet, supplemented with mankai duckweed, preserves iron-homeostasis in humans and is efficient in reversal of anemia in rats. J. Nutr. 2019, 149. [Google Scholar] [CrossRef]
- Ball, G.F.M. Microbiological methods for the determination of the B-group vitamins. In Water-Soluble Vitamin Assays in Human Nutrition; Springer: Berlin, Germany, 1994; pp. 317–364. [Google Scholar]
- Rinott, E.; Youngster, I.; Meir, A.Y.; Tsaban, G.; Zelicha, H.; Kaplan, A.; Knights, D.; Tuohy, K.; Fava, F.; Scholz, M.U.; et al. Effects of diet-modulated autologous fecal microbiota transplantation on weight regain. Gastroenterology 2020. [Google Scholar] [CrossRef]
- Shai, I.; Schwarzfuchs, D.; Henkin, Y.; Shahar, D.R.; Witkow, S.; Greenberg, I.; Golan, R.; Fraser, D.; Bolotin, A.; Vardi, H.; et al. Weight loss with a low-carbohydrate, Mediterranean, or low-fat diet. N. Engl. J. Med. 2008, 359, 229–241. [Google Scholar] [CrossRef]
- Gepner, Y.; Shelef, I.; Schwarzfuchs, D.; Zelicha, H.; Tene, L.; Meir, A.Y.; Tsaban, G.; Cohen, N.; Bril, N.; Rein, M.; et al. Effect of distinct lifestyle interventions on mobilization of fat storage pools: Central magnetic resonance imaging randomized controlled trial. Circulation 2018, 137, 1143–1157. [Google Scholar] [CrossRef]
- Moll, R.; Davis, B. Iron, vitamin B12 and folate. Medicine 2017, 45, 198–203. [Google Scholar] [CrossRef]
- McDonald, J.A.K.; Schroeter, K.; Fuentes, S.; Heikamp-deJong, I.; Khursigara, C.M.; de Vos, W.M.; Allen-Vercoe, E. Evaluation of microbial community reproducibility, stability and composition in a human distal gut chemostat model. J. Microbiol. Methods 2013, 95, 167–174. [Google Scholar] [CrossRef]
- Gutierrez, D.; Weinstock, A.; Antharam, V.C.; Gu, H.; Jasbi, P.; Shi, X.; Dirks, B.; Krajmalnik-Brown, R.; Maldonado, J.; Guinan, J.; et al. Antibiotic-induced gut metabolome and microbiome alterations increase the susceptibility to Candida albicans colonization in the gastrointestinal tract. FEMS Microbiol. Ecol. 2020, 96, fiz187. [Google Scholar] [CrossRef] [PubMed]
- Ilhan, Z.E.; di Baise, J.K.; Dautel, S.E.; Isern, N.G.; Kim, Y.M.; Hoyt, D.W.; Schepmoes, A.A.; Brewer, H.M.; Weitz, K.K.; Metz, T.O.; et al. Temporospatial shifts in the human gut microbiome and metabolome after gastric bypass surgery. NPJ Biofilms Microbiomes 2020, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jägerstad, M.; Arkbåge, K. Cobalamins properties and determination. In Encyclopedia of Food Sciences and Nutrition; Caballero, B., Trugo, L.C., Finglas, P.M., Eds.; Academic: Cambridge, MA, USA, 2003; p. 1419. [Google Scholar]
- Watanabe, F.; Katsura, H.; Takenaka, S.; Fujita, T.; Abe, K.; Tamura, Y.; Nakatsuka, T.; Nakano, Y. Pseudovitamin B12 is the predominant cobamide of an algal health food, spirulina tablets. J. Agric. Food Chem. 1999, 47, 4736–4741. [Google Scholar] [CrossRef] [PubMed]
- Chamlagain, B.; Edelmann, M.; Kariluoto, S.; Ollilainen, V.; Piironen, V. Ultra-high performance liquid chromatographic and mass spectrometric analysis of active vitamin B12 in cells of Propionibacterium and fermented cereal matrices. Food Chem. 2015, 166, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, F.; Yabuta, Y.; Bito, T.; Teng, F. Vitamin B12-containing plant food sources for vegetarians. Nutrients 2014, 6, 1861–1873. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, F. Vitamin B12 sources and bioavailability. Exp. Biol. Med. 2007, 232, 1266–1274. [Google Scholar] [CrossRef]
- Fedosov, S.N.; Fedosova, N.U.; Kräutler, B.; Nexø, E.; Petersen, T.E. Mechanisms of discrimination between cobalamins and their natural analogues during their binding to the specific B12-transporting proteins. Biochemistry 2007, 46, 6446–6458. [Google Scholar] [CrossRef]
- Stupperich, E.; Nexø, E. Effect of the cobalt-N coordination on the cobamide recognition by the human vitamin B12 binding proteins intrinsic factor, transcobalamin and haptocorrin. Eur. J. Biochem. 1991, 199, 299–303. [Google Scholar] [CrossRef]
- Herbert, V.; Drivas, G. Spirulina and vitamin B12. JAMA 1982, 248, 3096–3097. [Google Scholar] [CrossRef]
- Bito, T.; Bito, M.; Hirooka, T.; Okamoto, N.; Harada, N.; Yamaji, R.; Nakano, Y.; Inui, H.; Watanabe, F. Biological activity of pseudovitamin B12 on cobalamin-dependent methylmalonyl-CoA mutase and methionine synthase in mammalian cultured COS-7 Cells. Molecules 2020, 25, 3268. [Google Scholar] [CrossRef] [PubMed]
- Schauer, K.; Rodionov, D.A.; de Reuse, H. New substrates for TonB-dependent transport: Do we only see the ‘tip of the iceberg’? Trends Biochem. Sci. 2008, 33, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xiang, S.; Ye, K.; Zheng, Y.; Feng, X.; Zhu, X.; Chen, J.; Chen, Y. Cobalamin (vitamin B12) induced a shift in microbial composition and metabolic activity in an in vitro colon simulation. Front. Microbiol. 2018, 9, 2780. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Iñiguez, T.; García-Hernandez, E.; Arreguín-Espinosa, R.; Flores, M.E. Role of vitamin B12 on methylmalonyl-CoA mutase activity. J. Zhejiang Univ. Sci. B 2012, 13, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Ragsdale, S.W. The many faces of vitamin B12: Catalysis by cobalamin-dependent enzymes. Annu. Rev. Biochem. 2003, 72, 209–247. [Google Scholar] [CrossRef]
- Buckel, W. Unusual enzymes involved in five pathways of glutamate fermentation. Appl. Microbiol. Biotechnol. 2001, 57, 263–273. [Google Scholar] [CrossRef]
- Van Treuren, W.; Dodd, D. Microbial contribution to the human Metabolome: Implications for health and disease. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 345–369. [Google Scholar] [CrossRef]
- Food and Nutrition Board, Institute of Medicine. Dietary Reference Intakes: Recommended Intakes for Individuals; National Academy of Sciences: Washington, DC, USA, 2004.
- Micha, R.; Penalvo, J.L.; Cudhea, F.; Imamura, F.; Rehm, C.D.; Mozaffarian, D. association between dietary factors and mortality from heart disease, stroke, and Type 2 diabetes in the united states. JAMA 2017, 317, 912–924. [Google Scholar] [CrossRef]
- Obeid, R.; Heil, S.G.; Verhoeven, M.M.A.; van den Heuvel, E.G.H.M.; de Groot, L.C.; Eussen, S.J.P.M. Vitamin B12 intake from animal foods, biomarkers, and health aspects. Front. Nutr. 2019, 6, 93. [Google Scholar] [CrossRef]
- Green, R.; Allen, L.H.; Bjørke-Monsen, A.L.; Brito, A.; Guéant, J.L.; Miller, J.W.; Molloy, A.M.; Nexo, E.; Stabler, S.; Toh, B.H.; et al. Vitamin B 12 deficiency. Nat. Rev. Dis. Prim. 2017, 3, 17040. [Google Scholar] [CrossRef]
- Oussalah, A.; Levy, J.; Berthezène, C.; Alpers, D.H.; Guéant, J.L. Health outcomes associated with vegetarian diets: An umbrella review of systematic reviews and meta-analyses. Clin. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Lebacq, T.; Baret, P.V.; Stilmant, D. Sustainability indicators for livestock farming. A review. Agron. Sustain. Dev. 2013, 33, 311–327. [Google Scholar] [CrossRef]
- Van Zanten, H.H.E.; Herrero, M.; van Hal, O.; Röös, E.; Muller, A.; Garnett, T.; Gerber, P.J.; Schader, C.; Boer, I.J.M. De Defining a land boundary for sustainable livestock consumption. Glob. Chang. Biol. 2018, 24, 4185–4194. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, F.; Yabuta, Y.; Tanioka, Y.; Bito, T. Biologically active vitamin B12 compounds in foods for preventing deficiency among vegetarians and elderly subjects. J. Agric. Food Chem. 2013, 61, 6769–6775. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H. Serum vitamin B12 levels in young vegans who eat brown rice. J. Nutr. Sci. Vitaminol. 1995, 41, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Dagnelie, P.C.; van Staveren, W.A.; van den Berg, H. Vitamin B-12 from algae appears not to be bioavailable. Am. J. Clin. Nutr. 1991, 53, 695–697. [Google Scholar] [CrossRef]
- Schmidt, A.; Call, L.M.; Macheiner, L.; Mayer, H.K. Determination of vitamin B12 in four edible insect species by immunoaffinity and ultra-high performance liquid chromatography. Food Chem. 2019, 281, 124–129. [Google Scholar] [CrossRef]
- Klee, G.G. Cobalamin and folate evaluation: Measurement of methylmalonic acid and homocysteine vs vitamin B12 and folate. Clin. Chem. 2000, 46, 1277–1283. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entire n = 294 | Lowest Tertile n = 99 | Intermediate Tertile n = 98 | Highest Tertile n = 97 | p Between Tertiles 1 | p Between Extreme Tertiles 2 | |
|---|---|---|---|---|---|---|
| Vitamin B12, pg/mL | 420.4 ± 187 | 261.2 ± 46.1 | 385.7 ± 37.9 | 618.1 ±1 92.8 | - | - |
| Age, years | 51.1 ± 10.5 | 51.9 ± 9.6 | 49.7 ± 10.5 | 51.5 ± 11.4 | 0.25 | 0.54 |
| Men, number | 259 | 87 | 86 | 86 | 0.98 | - |
| BMI, kg/m2 | 31.3 ± 4.0 | 31.3 ± 4.3 | 31.3 ± 3.9 | 31.2 ± 3.7 | 0.84 | 0.87 |
| WC, cm | 109.7 ± 9.5 | 110.1 ± 9.7 | 109.6 ± 10.5 | 109.4 ± 8.1 | 0.67 | 0.93 |
| Fasting glucose, mg/dL | 101.9 ± 17.1 | 104.0 ± 19.5 | 101.2 ± 14.7 | 100.6 ± 16.5 | 0.62 | 0.35 |
| Cholesterol, mg/dL | 190.6 ± 33.0 | 190.9 ± 29.5 | 190.9 ± 33.8 | 189.8 ± 35.9 | 0.97 | 0.82 |
| HDL-c, mg/dL | 46.0 ± 11.7 | 45.0 ± 12.4 | 46.2 ± 11.3 | 46.7 ± 11.3 | 0.29 | 0.15 |
| LDL-c, mg/dL | 125.7 ± 30.1 | 125.5 ± 28.6 | 127.0 ± 31.6 | 124.5 ± 32.4 | 0.86 | 0.83 |
| Triglycerides, mg/dL | 146.3 ± 66.8 | 159.4 ± 66.9 | 139.8 ± 60.0 | 139.5 ± 68.8 | 0.02 | 0.01 |
| ALT, U/L | 34.9 ± 16.8 | 34.3 ± 14.4 | 35.4 ± 20.4 | 34.9 ± 15.0 | 0.79 | 0.71 |
| AST, U/L | 25.6 ± 7.7 | 25.5 ± 7.2 | 26.1 ± 8.7 | 25.3 ± 7.3 | 0.90 | 0.74 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sela, I.; Yaskolka Meir, A.; Brandis, A.; Krajmalnik-Brown, R.; Zeibich, L.; Chang, D.; Dirks, B.; Tsaban, G.; Kaplan, A.; Rinott, E.; et al. Wolffia globosa–Mankai Plant-Based Protein Contains Bioactive Vitamin B12 and Is Well Absorbed in Humans. Nutrients 2020, 12, 3067. https://doi.org/10.3390/nu12103067
Sela I, Yaskolka Meir A, Brandis A, Krajmalnik-Brown R, Zeibich L, Chang D, Dirks B, Tsaban G, Kaplan A, Rinott E, et al. Wolffia globosa–Mankai Plant-Based Protein Contains Bioactive Vitamin B12 and Is Well Absorbed in Humans. Nutrients. 2020; 12(10):3067. https://doi.org/10.3390/nu12103067
Chicago/Turabian StyleSela, Ilan, Anat Yaskolka Meir, Alexander Brandis, Rosa Krajmalnik-Brown, Lydia Zeibich, Debbie Chang, Blake Dirks, Gal Tsaban, Alon Kaplan, Ehud Rinott, and et al. 2020. "Wolffia globosa–Mankai Plant-Based Protein Contains Bioactive Vitamin B12 and Is Well Absorbed in Humans" Nutrients 12, no. 10: 3067. https://doi.org/10.3390/nu12103067
APA StyleSela, I., Yaskolka Meir, A., Brandis, A., Krajmalnik-Brown, R., Zeibich, L., Chang, D., Dirks, B., Tsaban, G., Kaplan, A., Rinott, E., Zelicha, H., Arinos, S., Ceglarek, U., Isermann, B., Lapidot, M., Green, R., & Shai, I. (2020). Wolffia globosa–Mankai Plant-Based Protein Contains Bioactive Vitamin B12 and Is Well Absorbed in Humans. Nutrients, 12(10), 3067. https://doi.org/10.3390/nu12103067