Milk and Meat Allergens from Bos taurus β-Lactoglobulin, α-Casein, and Bovine Serum Albumin: An In-Vivo Study of the Immune Response in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
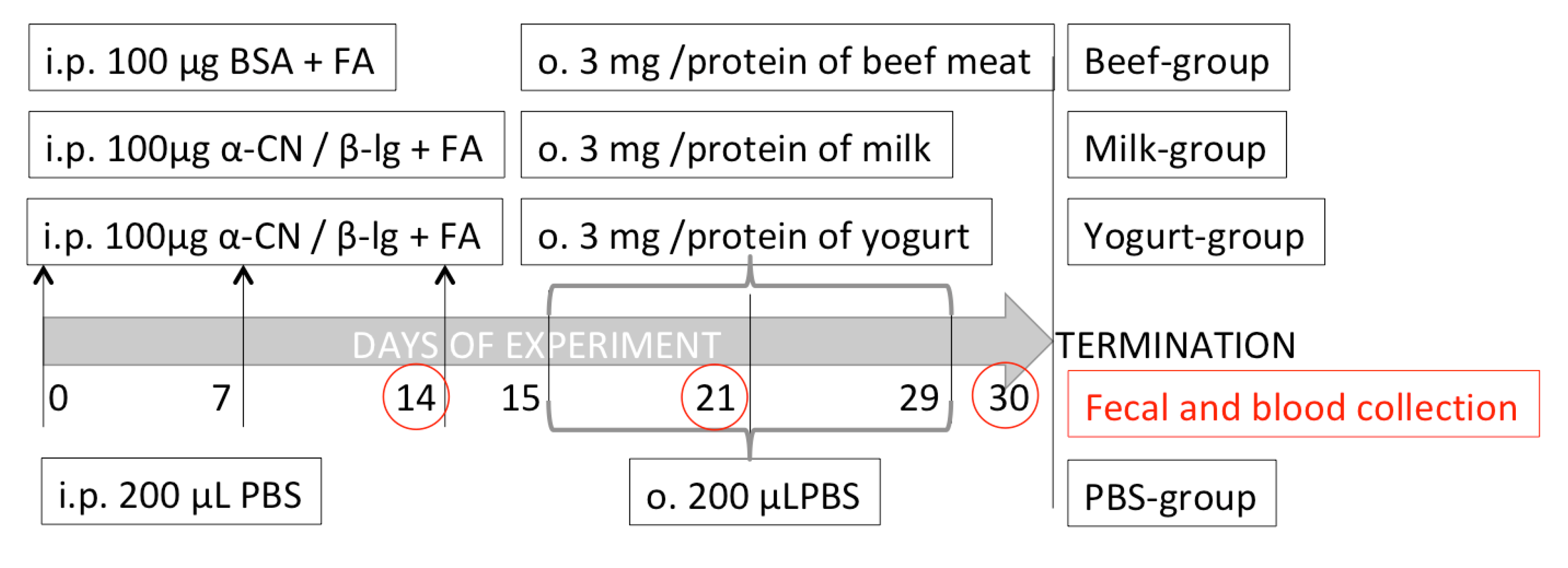
2.1. Mouse Diet Suplementation Products
2.2. Animals
2.3. Experimental Protocol
2.4. Isolation of Lymphocytes
2.5. Splenocyte Culture and Cytokine Measurement
2.6. Lymphocyte Phenotyping
2.7. Antibody Measurements
2.8. Quantification of Fecal Microbiota by qPCR
2.9. Statistical Analysis
3. Results
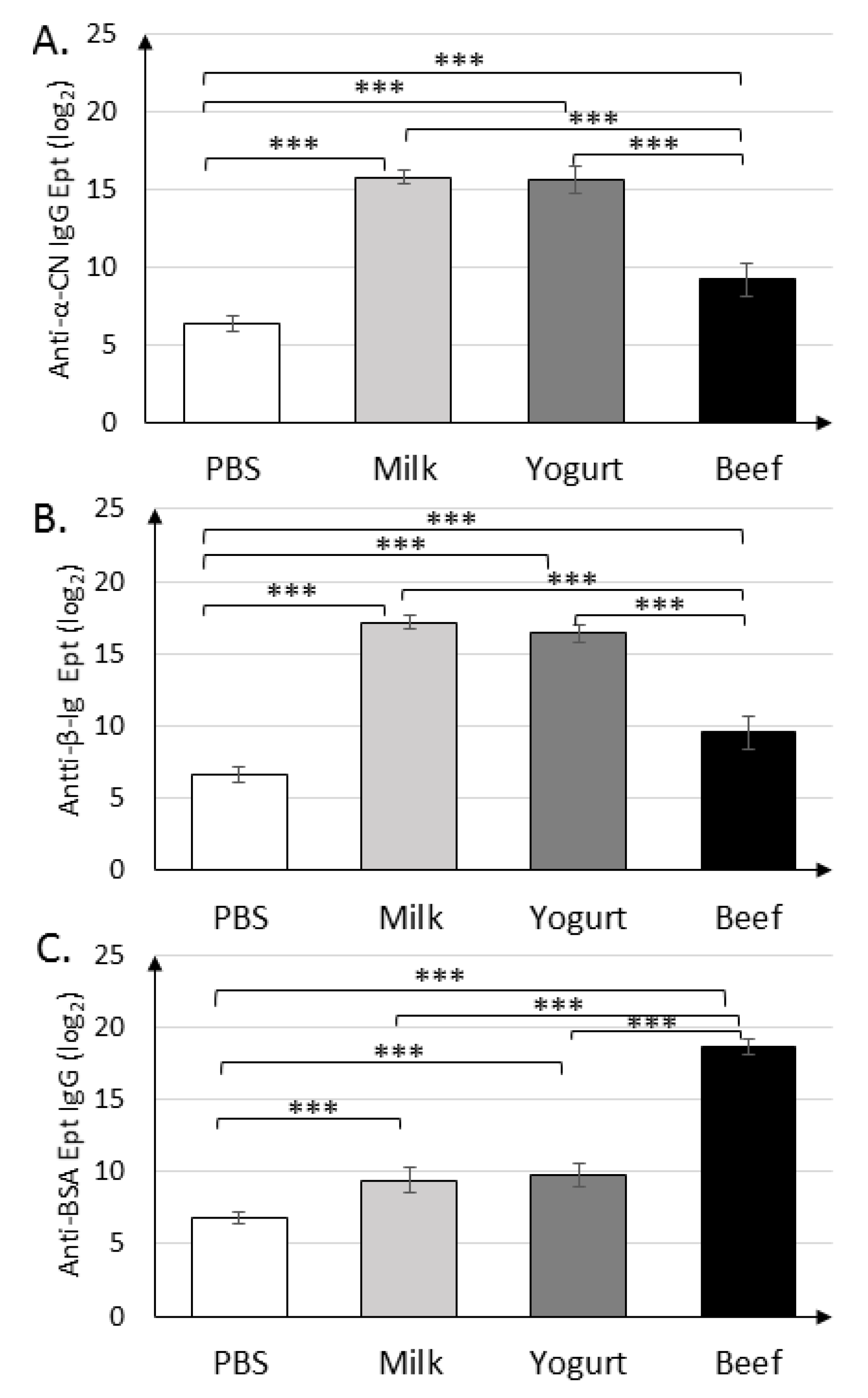
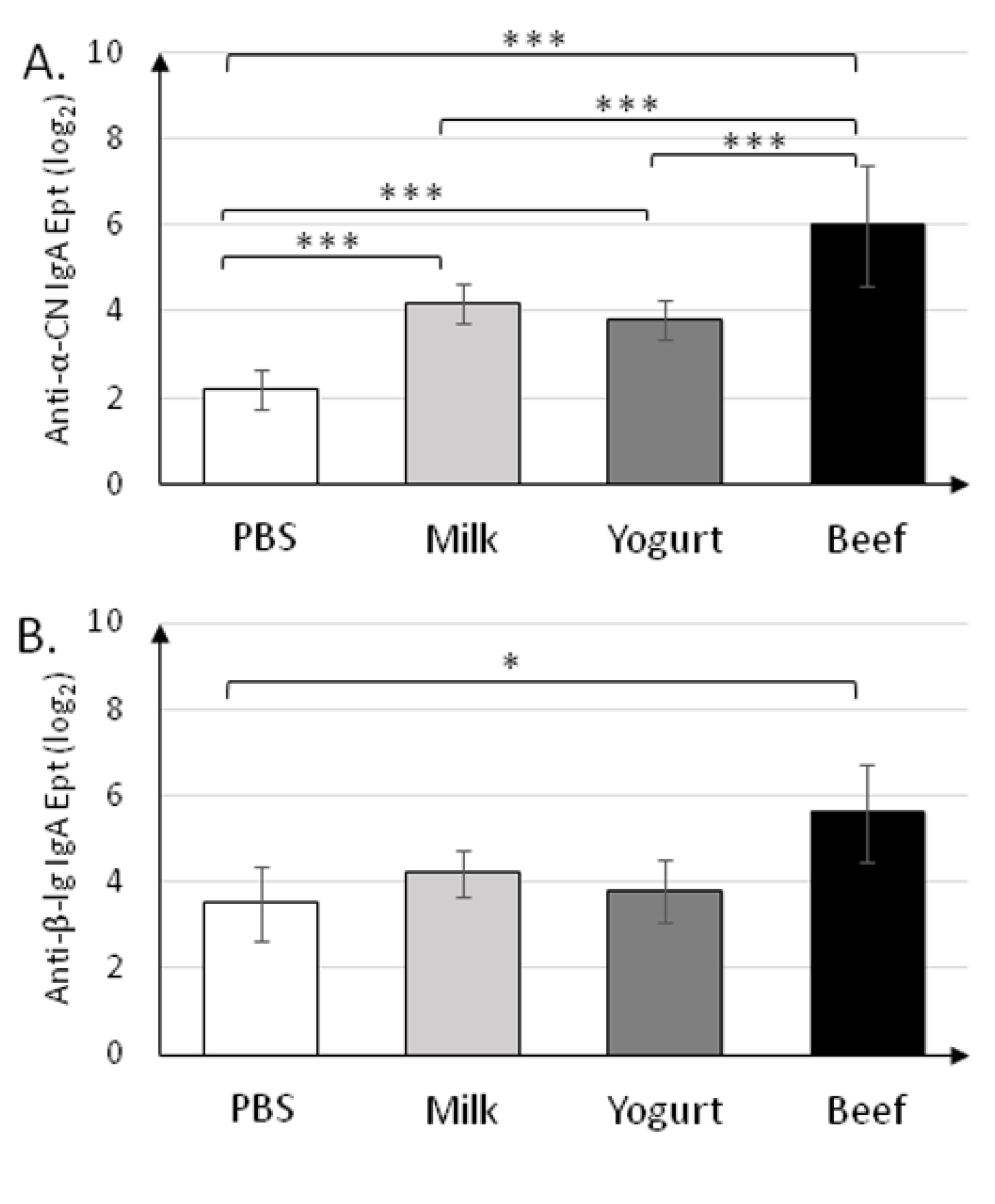
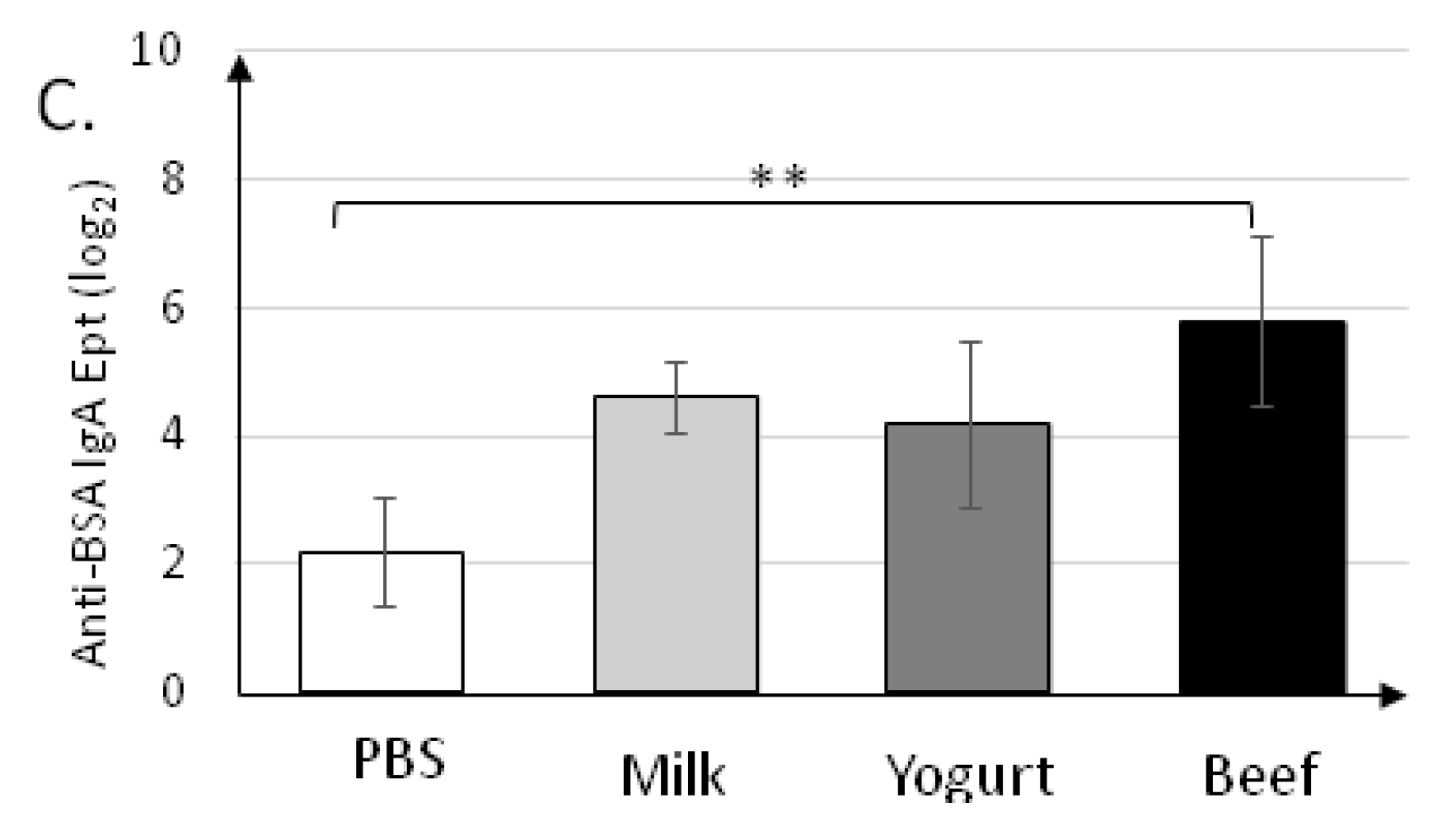
3.1. Yogurt Suplementation Decreases Humoral Response
3.2. Cellular Immune Responses
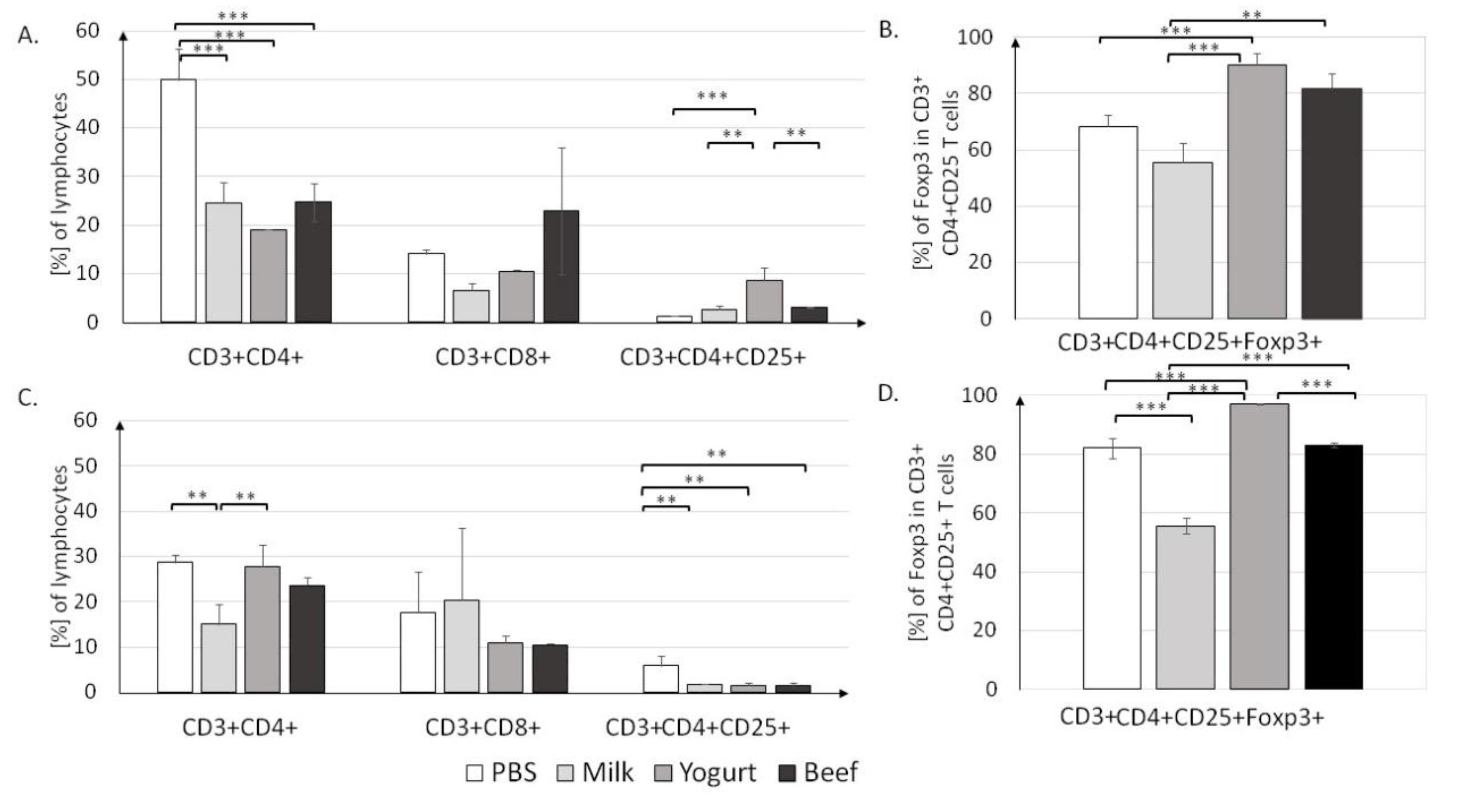
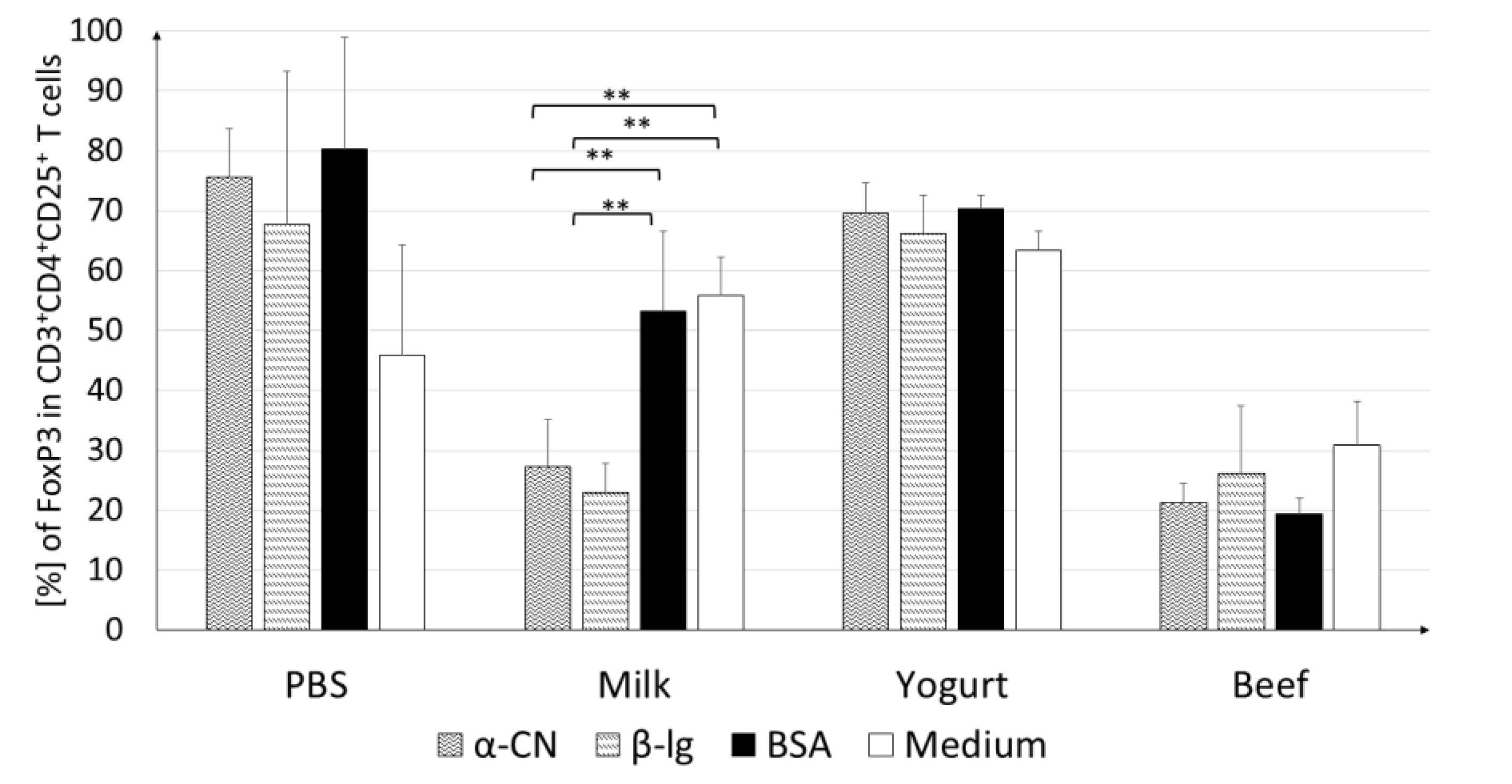
3.2.1. T Lymphocyte Profile in Lymphoid Tissues
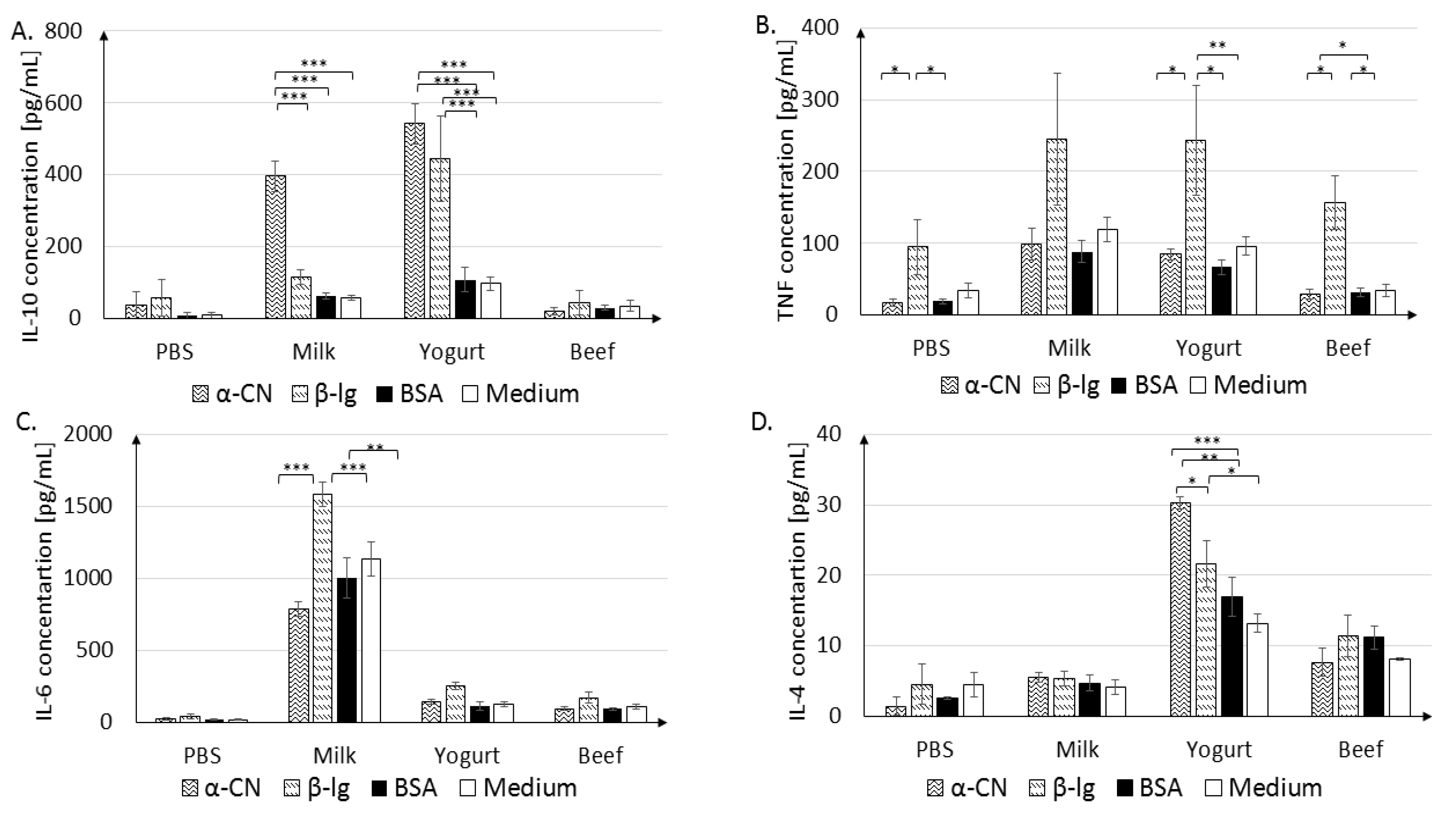
3.2.2. In Vitro Cytokine Secretion
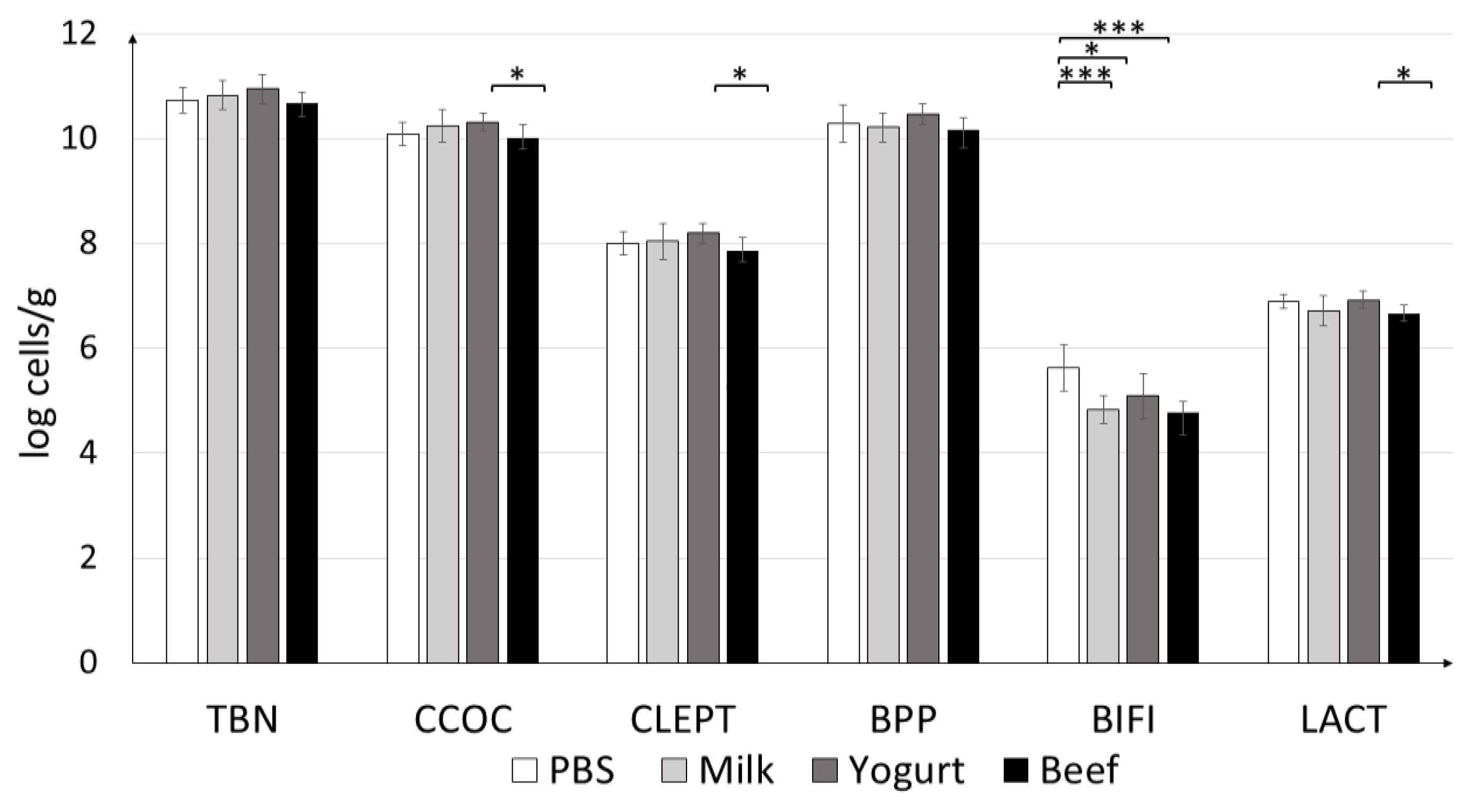
3.2.3. Quantification of Colonic Microbiota
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CMA | Cow’s milk allergy |
| α-CN | α-casein |
| β-lg | β-lactoglobulin |
| BSA | Bovine serum albumin |
| PBS | 0.1 M phosphate-buffered saline, pH = 7.4 |
| MLN | Mesenteric lymph nodes |
| PP | Payer’s patch |
| HNLN | Head and neck lymph nodes |
| SPL | Spleen |
| Ept | Endpoint titer |
References
- Cianferoni, A.; Spergel, J.M. Food Allergy: Review, classification and diagnosis. Allergol. Int. 2009, 58, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Tsabouri, S.; Douros, K.; Priftis, K.N. Cow’s milk allergenicity. Endocr. Metab. Immune Disord. Drug Targets 2014, 14, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Denney, L.; Afeiche, M.C.; Eldridge, A.L.; Villalpando-Carrión, S. Food Sources of Energy and Nutrients in Infants, toddlers and young children from the Mexican National Health and Nutrition Survey. Nutrients 2017, 9, 494. [Google Scholar] [CrossRef] [PubMed]
- Wróblewska, B.; Juśkiewicz, J.; Kroplewski, B.; Jurgoński, A.; Wasilewska, E.; Złotkowska, D.; Markiewicz, L.H. The effect of whey and soy proteins on growth performance, gastroinestinal digestion and selected physiological responses in rats. Food Funct. 2018, 9, 1500–1509. [Google Scholar] [CrossRef]
- Mierzejewska, D.; Kubicka, E. Effect of temperature on immumorective properties of cow milk whey protein. I. alpha-lactoalbumin. Milchwiss. Milk Sci. Int. 2003, 61, 237–246. [Google Scholar]
- Zlotkowska, D.; Kozlowska, J.; Rudnicka, B.; Kostyra, H. Immune responses of mouse mucosal system to glycated and hydrolysed OVA. FASEB J. 2010, 24 (Suppl. 1), 723-19. [Google Scholar]
- Bu, G.; Luo, Y.; Chen, F. Milk processing as a toll to reduce cow’s milk allergenicity: A Mini-Review. Dairy Sci. Technol. 2013, 93, 211–223. [Google Scholar] [CrossRef]
- Wróblewska, B.; Kaliszewska-Suchodoła, A.; Kołakowski, P.; Troszyńska, A. The effect of microbial transglutaminase on the immunoreactive and sensory properties of fermented milk beverages. Int. J. Food Sci. Tech. 2012, 48, 1007–1017. [Google Scholar] [CrossRef]
- Umetsu, D.T.; DeKruyff, R.H. Th1 and Th2 CD4+ cells in human allergic diseases. J. Allergy Clin. Immunol. 1997, 100, 1–6. [Google Scholar] [CrossRef]
- Orozco, E.M.; Norte-Munoz, M.; Martinez-Gracia, J. Regulatory T cells in allergy and asthma. Front. Pediatr. 2017, 5, 117. [Google Scholar] [CrossRef]
- Miyahara, N.; Takeda, K.; Kodoma, T.; Joetham, A.; Taube, C.; Park, J.W.; Miyahara, S.; Balhorn, A.; Dakhama, A.; Gelfand, E.W. Contribution of Antigen-primed CD8+T cells to the development of airway hyperresponsiveness and inflammation is associated with IL-13. J. Immunol. 2004, 172, 2549–2558. [Google Scholar] [CrossRef] [PubMed]
- Allergen Nomenclature Sub-committee. Available online: www.allergen.org (accessed on 10 June 2019).
- Villa, C.; Costa, J.; Oliveira, M.B.P.P.; Mafra, I. Bovine Milk Allergens: A comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 137–164. [Google Scholar] [CrossRef]
- Kelly, P.; Woonton, B.W.; Smither, G.W. Improving the sensory quality, shelf-life and functionality of milk. In Functional and Speciality Beverage Technology; Woodhead Publishing: Sawston, UK, 2009; pp. 170–231. [Google Scholar] [CrossRef]
- Refaat, M.; Kamal, A.; Fares, M.; Ossman, E.; Attia, M.; Elokda, A. Beef meat allergy in Cow’s milk allergenic adults. Food Nutr. Sci. 2011, 2, 891–894. [Google Scholar] [CrossRef]
- Hochwallner, H.; Schulmeister, U.; Swoboda, I.; Spitzauer, S.; Valenta, R. Cow’s milk allergy: From allergens to new forms of diagnosis, therapy and prevention. Methods 2014, 66, 22–23. [Google Scholar] [CrossRef] [PubMed]
- Restani, P.; Fiocchi, A.; Beretta, B.; Velona, T.; Giovannini, M.; Galli, C.L. Meat allergy III—Proteins involved and cross-reactivity between different animal species. J. Am. Coll. Nutr. 1997, 16, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Fiocchi, A.; Brozek, J.; Schünemann, H.; Bahna, S.L.; von Berg, A.; Beyer, K.; Bozzola, M.; Bradsher, J.; Compalati, E.; Ebisawa, M.; et al. World Allergy Organization (WAO) Diagnosis and Rationale for Action against Cow’s Milk allergy (DRACMA) Guidelines. Pediatr. Allergy Immunol. 2010, 21, 1–125. [Google Scholar] [CrossRef]
- Chruszcz, M.; Mikołajczak, K.; Mank, N.; Majorek, K.A.; Porębski, P.J.; Minor, W. Serum albumins-unusual allergens. Biochim. Biophys. Acta-Gen. Subj. 2013, 1830, 5375–5381. [Google Scholar] [CrossRef]
- Marteli, A.; de Chiara, A.; Corvo, M.; Restani, P.; Fiocchi, A. Beff Allergy in children with Cows’s milk allergy; Cows’s milk allergy in children with Beef allergy. Ann. Allergy Asthma Immunol. 2002, 89, 38–43. [Google Scholar] [CrossRef]
- AOAC. Available online: http://cem.com/media/wysiwyg/sprint/AOACMethod2011_04_Protein_in_Meat.pdf (accessed on 28 March 2019).
- Lynch, J.M.; Barbano, D.M. Kjeldahl Nitrogen Analysis as a Reference Method for protein Determination in dairy Products. J. AOAC Int. 1999, 82, 1389–1398. [Google Scholar]
- Mierzejewska, D.; Mitrowska, P.; Rudnicka, B.; Kubicka, E.; Kostyra, H. Effect of non enzymatic glycosylation of pea albumins on their immunorective properties. Food Chem. 2008, 111, 127–131. [Google Scholar] [CrossRef]
- Fuc, E.; Zlotkowska, D.; Wróblewska, B. Immunoreactive properties of α-casein and κ-casein: Ex Vivo and In Vivo studies. J. Dairy Sci. 2018, 101, 10703–10713. [Google Scholar] [CrossRef] [PubMed]
- Fotschki, J.; Szyc, A.M.; Laparra, J.M.; Markiewicz, L.H.; Wróblewska, B. Immune-modulating properties of horse milk administered to mice sensitized to cow milk. J. Dairy Sci. 2016, 99, 9395–9404. [Google Scholar] [CrossRef] [PubMed]
- Drabińska, N.; Jarocka-Cyrta, E.; Markiewicz, L.H.; Krupa-Kozak, U. The Effect of Oligofructose-Enriched Inulin of Feacal Bacterial Counts and microbiota-Associated Characteristicsm in Celiac Disease Children Following a Gluten-Free Diet: Results of a Randomized, Placebo-Controlled Trial. Nutrients 2018, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Garcia, B.E.; Lizaso, M.T. Cross-reactivity syndromes in food allergy. J. Investig. Allergol. Clin. Immunol. 2011, 21, 162–170. [Google Scholar] [PubMed]
- Gocki, J.; Bartuzi, Z. Role of Immunoglobulin G Antibodies in Diagnosis of Food Allergy. Postęp. Dermatol. Alergol. 2016, 33, 253–256. [Google Scholar] [CrossRef]
- Wachholz, P.A.; Durham, S.R. Mechanisms of immunotherapy: IgG revisited. Curr. Opin. Allergy Clin. Immunol. 2004, 4, 313–318. [Google Scholar] [CrossRef]
- Aramburo-Galvez, J.G.; Sotelo-Cruz, N.; Flores-Mendoza, L.K.; Gracia-Valenzuela, M.H.; Chiquete-Elizalde, F.I.R.; Espinoza-Alderete, J.G.; Trejo-Martinez, H.; Canizalez-Roman, V.A.; Ontiveros, N.; Cabrera-Chavez, F. Assessment of the sensitizng potential of proteins in BALB/c mice: Comparison of three protocols of Intraperitoneal sensitization. Nutrients 2018, 10, 903. [Google Scholar] [CrossRef]
- Corthésy, B. Roundtrip Ticket for Secretory IgA: Role in Mucosal Homoestasis? J. Immunol. 2007, 178, 27–32. [Google Scholar] [CrossRef]
- Frossard, C.P.; Tropia, L.; Hauser, C.; Eigenmann, P.A. Lymphocytes in Payer patches regulate clinical tolerance in a murine model of food allergy. J. Allergy Clin. Immunol. 2004, 113, 958–964. [Google Scholar] [CrossRef]
- Wróblewska, B.; Kaliszewska, A.; Markiewicz, L.H.; Szyc, A.M.; Wasilewska, E. Whey prefermented with beneficial microbes modulates immune response and lowers responsiveness to milk allergens in mouse model. J. Funct. Foods 2019, 54, 41–52. [Google Scholar] [CrossRef]
- Kaminogawa, S.; Hachimura, S.; Nakajima-Adachi, H.; Totsuka, M. Food allergens and mucosal immune system with special reference to recognition of food allergens by gut associated lyphoid tissue. Allergol. Int. 1999, 48, 15–23. [Google Scholar] [CrossRef]
- Calzada, D.; Baos, S.; Cremades-Jimeno, L.; Cardaba, B. Immunological mechanism in allergic diseases and allergen tolerance: The role of Treg cells. J. Immunol Res. 2018. [Google Scholar] [CrossRef]
- Noval Rivas, M.; Chatila, C. Regulatory T Cells in Allergic Diseases. J. Allergy Clin. Immunol. 2016, 138, 639–652. [Google Scholar] [CrossRef]
- Kondělková, K.; Vokurková, D.; Krejsek, J.; Borská, L.; Fiala, Z.; Andrýs, A. Regulatory T cells (TREG) and their roles in immune system with respectm to immunopathological disorders. Acta Med. 2010, 53, 73–77. [Google Scholar]
- Mucida, D.; Kutchukhidze, N.; Erza, A.; Risso, M.; Lafaille, J.J.; Curotto de Lafaille, M.A. Oral tolerance in the absence of naturally occurings Tregs. J. Clin. Investig. 2005, 115, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Couper, K.N.; Blount, D.G.; Riley, E.M. Il-10: The Master Regulator of Immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef] [PubMed]
- Feehley, T.; Stefka, A.T.; Cai, S.; Nagler, C.R. Microbial regulation of allergic responses to food. Semin. Immunopathol. 2012, 34, 671–688. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Björkstén, B.; Naaber, P.; Seep, E.; Mikelsaar, M. The intestinal microflora in allergic Estonian and Swedish 2-year-old children. Clin. Exp. Allergy 1999, 29, 342–346. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.T.; Saito, T.; Ohba, Y.; et al. Induction of colonic Regultory T Cells by indigenous Clostridium Species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuc, E.; Złotkowska, D.; Wróblewska, B. Milk and Meat Allergens from Bos taurus β-Lactoglobulin, α-Casein, and Bovine Serum Albumin: An In-Vivo Study of the Immune Response in Mice. Nutrients 2019, 11, 2095. https://doi.org/10.3390/nu11092095
Fuc E, Złotkowska D, Wróblewska B. Milk and Meat Allergens from Bos taurus β-Lactoglobulin, α-Casein, and Bovine Serum Albumin: An In-Vivo Study of the Immune Response in Mice. Nutrients. 2019; 11(9):2095. https://doi.org/10.3390/nu11092095
Chicago/Turabian StyleFuc, Ewa, Dagmara Złotkowska, and Barbara Wróblewska. 2019. "Milk and Meat Allergens from Bos taurus β-Lactoglobulin, α-Casein, and Bovine Serum Albumin: An In-Vivo Study of the Immune Response in Mice" Nutrients 11, no. 9: 2095. https://doi.org/10.3390/nu11092095
APA StyleFuc, E., Złotkowska, D., & Wróblewska, B. (2019). Milk and Meat Allergens from Bos taurus β-Lactoglobulin, α-Casein, and Bovine Serum Albumin: An In-Vivo Study of the Immune Response in Mice. Nutrients, 11(9), 2095. https://doi.org/10.3390/nu11092095