High-dose Glycerol Monolaurate Up-Regulated Beneficial Indigenous Microbiota without Inducing Metabolic Dysfunction and Systemic Inflammation: New Insights into Its Antimicrobial Potential





Abstract

1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Experimental Processes
2.3. Histology Analysis
2.4. Quantitative Reverse-Transcription PCR (qRT-PCR) Analysis
2.5. Plasma Parameters Analysis
2.6. Fecal Microbiota Analysis by 16S rRNA Gene Sequencing
2.7. Short-Chain Fatty Acids Composition Analysis
2.8. Statistical Analysis
3. Results
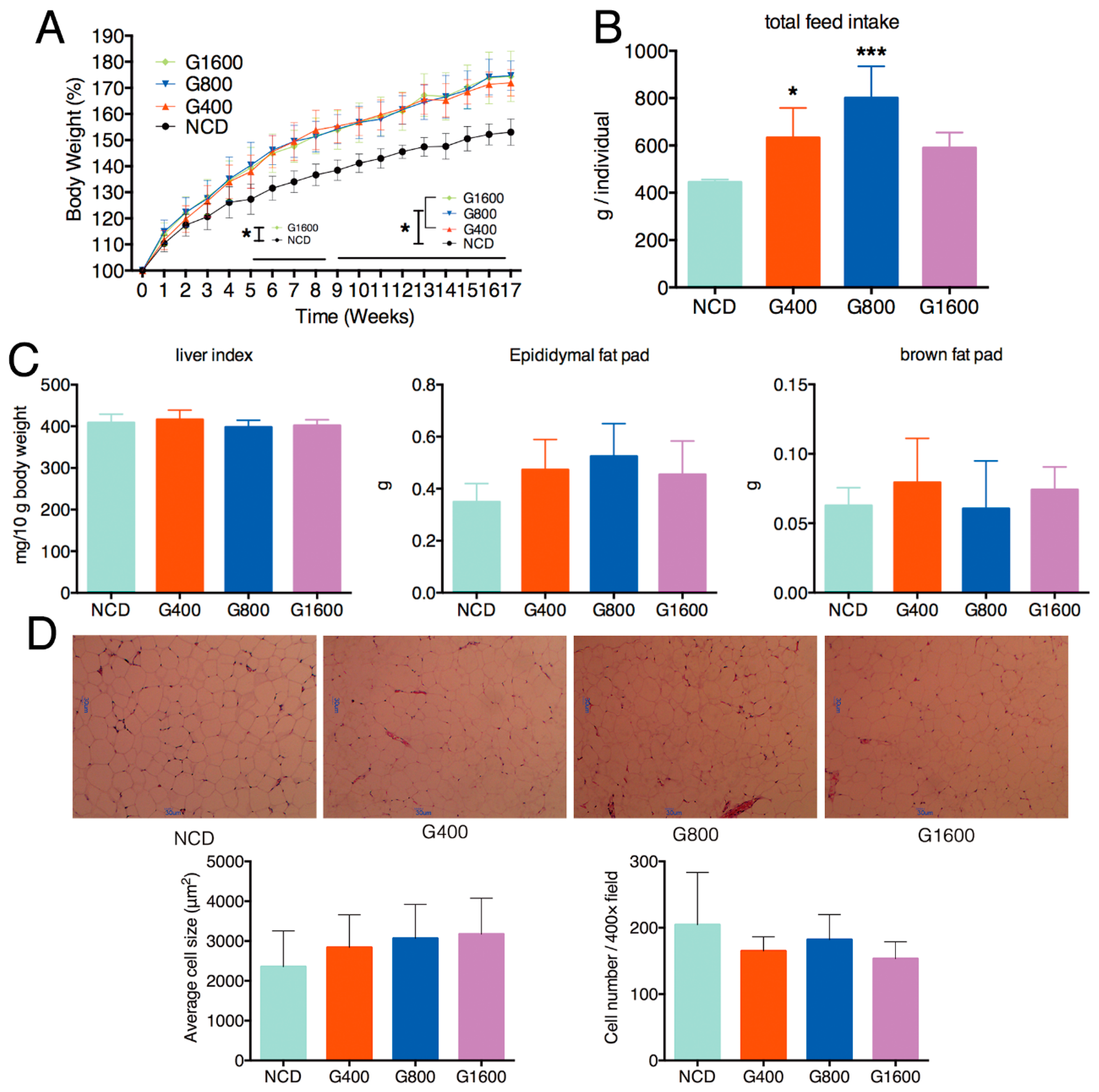
3.1. Effect of GML on the Body Weight, Feed Intake, Liver Index and Adipocyte size
3.2. Effect of GML on the Blood Biochemical Parameters
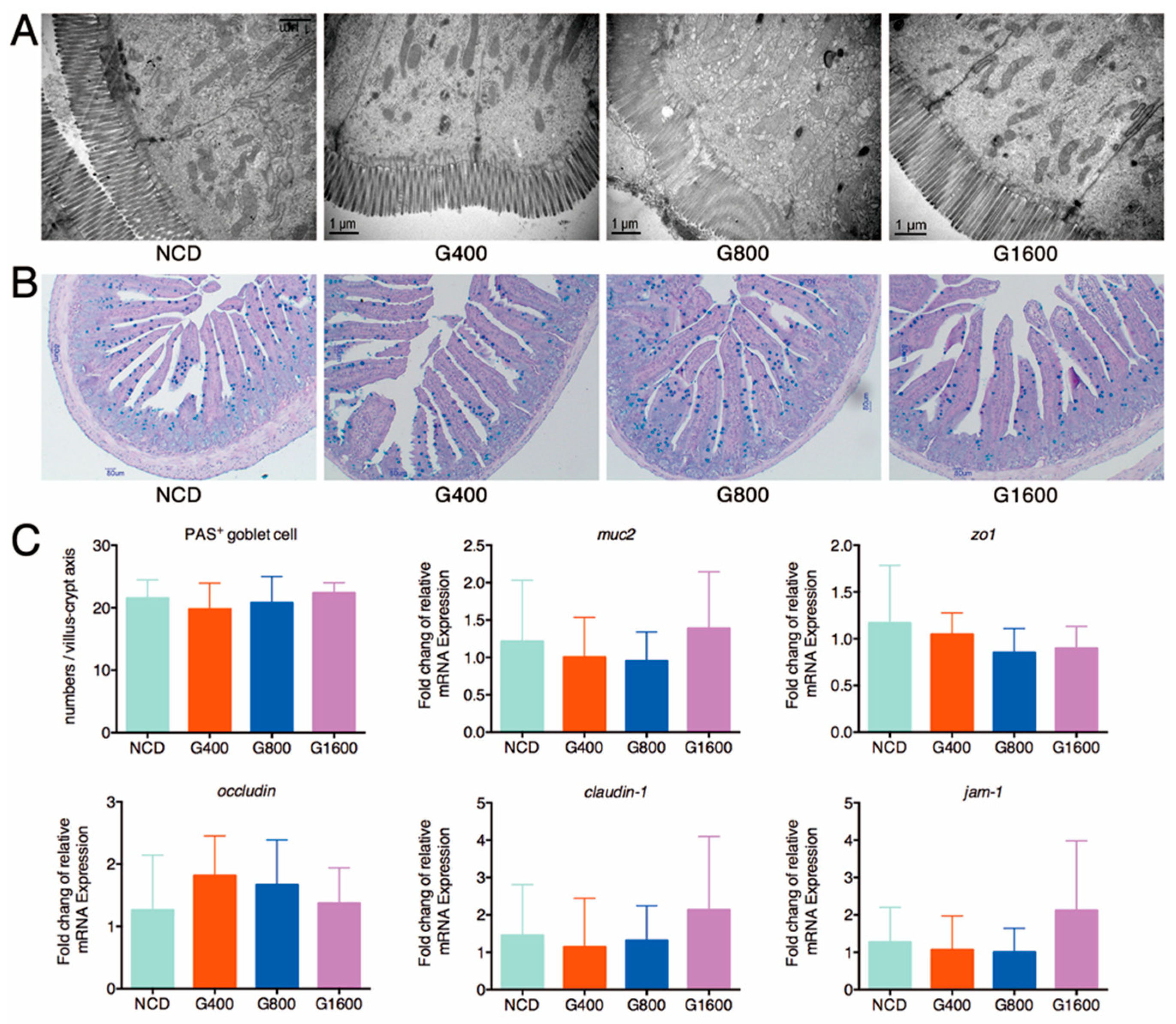
3.3. Effect of GML on the Histological Feature and Barrier Function of Intestine
3.4. Effect of GML on Serum Inflammatory-Related Parameters
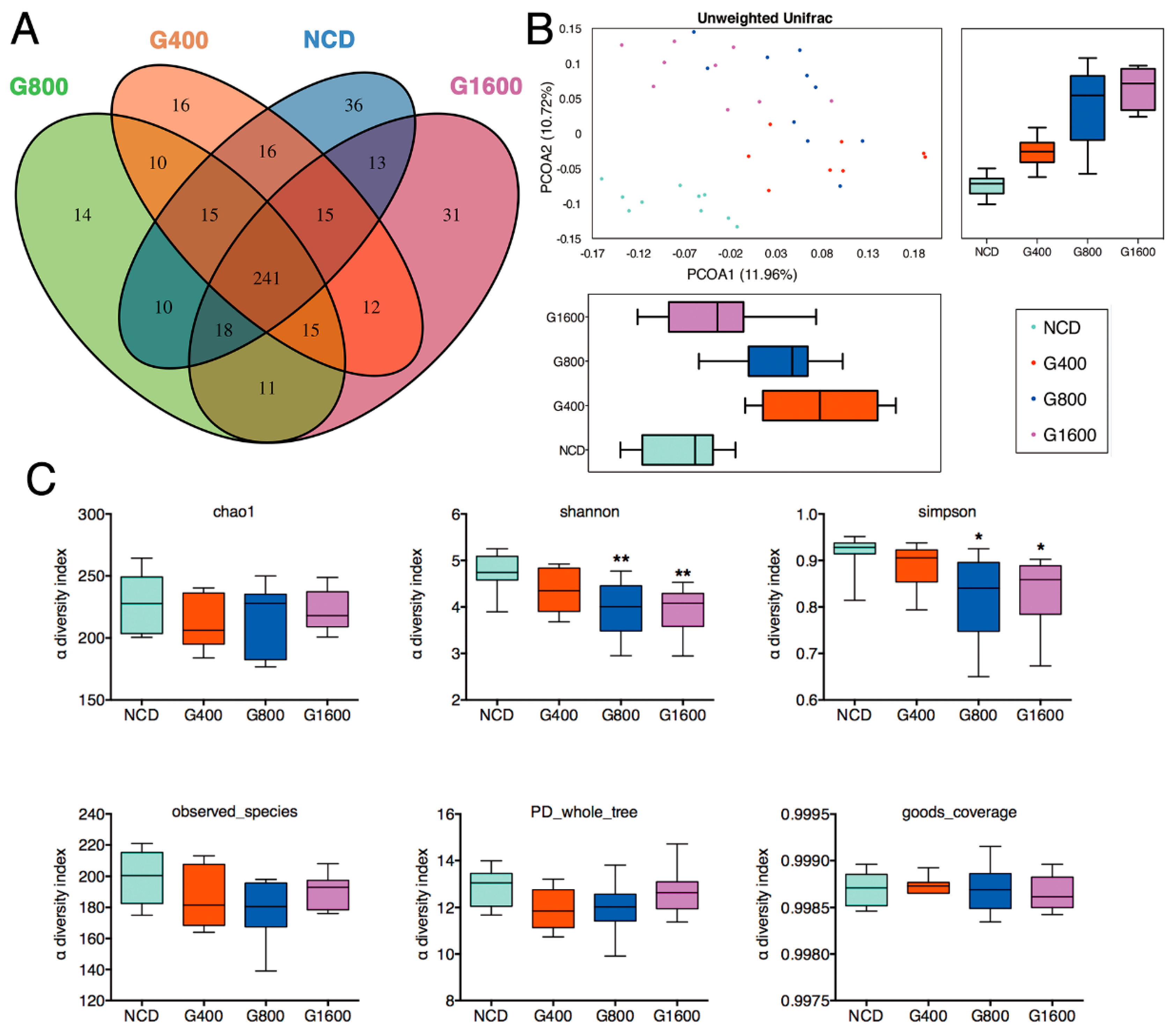
3.5. Effects of GML on Fecal Microbiota
3.6. Effect of GML on Phylotypes
3.7. Effects of GML on Fecal Short Chain Fatty Acid
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schlievert, P.M.; Peterson, M.L. Glycerol Monolaurate Antibacterial Activity in Broth and Biofilm Cultures. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.S.; Estes, J.D.; Schlievert, P.M.; Duan, L.J.; Brosnahan, A.J.; Southern, P.J.; Reilly, C.S.; Peterson, M.L.; Schultz-Darken, N.; Brunner, K.G.; et al. Glycerol monolaurate prevents mucosal SIV transmission. Nature 2009, 458, 1034–1038. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M.; Strandberg, K.L.; Brosnahan, A.J.; Peterson, M.L.; Pambuccian, S.E.; Nephew, K.R.; Brunner, K.G.; Schultz-Darken, N.J.; Haase, A.T. Glycerol Monolaurate Does Not Alter Rhesus Macaque (Macaca mulatta) Vaginal Lactobacilli and Is Safe for Chronic Use. Antimicrob. Agents Chemother. 2008, 52, 4448–4454. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M.; Kilgore, S.H.; Kaus, G.M.; Ho, T.D.; Ellermeier, C.D. Glycerol Monolaurate (GML) and a Nonaqueous Five-Percent GML Gel Kill Bacillus and Clostridium Spores. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, K.L.; Peterson, M.L.; Schaefers, M.M.; Case, L.C.; Pack, M.C.; Chase, D.J.; Schlievert, P.M. Reduction in Staphylococcus aureus Growth and Exotoxin Production and in Vaginal Interleukin 8 Levels Due to Glycerol Monolaurate in Tampons. Clin. Infect. Dis. 2009, 49, 1711–1717. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.S.; Sandouk, A.; Houtman, J.C.D. Glycerol Monolaurate (GML) inhibits human T cell signaling and function by disrupting lipid dynamics. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Itav, S.; Rothschild, D.; Meijer, M.T.; Levy, M.; Moresi, C.; Dohnalova, L.; Braverman, S.; Rozin, S.; Malitsky, S.; et al. Persistent microbiome alterations modulate the rate of post-dieting weight regain. Nature 2016, 540, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.L.; Zhao, M.J.; Zhang, H.; Li, Y.; Liu, M.Y.; Feng, F.Q. Antimicrobial Emulsifier-Glycerol Monolaurate Induces Metabolic Syndrome, Gut Microbiota Dysbiosis, and Systemic Low-Grade Inflammation in Low-Fat Diet Fed Mice. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef]
- Li, Y.; Liu, T.; Zhang, X.; Zhao, M.; Zhang, H.; Feng, F. Lactobacillus plantarum helps to suppress body weight gain, improve serum lipid profile and ameliorate low-grade inflammation in mice administered with glycerol monolaurate. J. Funct. Foods 2019, 53, 54–61. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Kim, M.C.; Cho, N.-J. Spectrum of membrane morphological responses to antibacterial fatty acids and related surfactants. Langmuir 2015, 31, 10223–10232. [Google Scholar] [CrossRef] [PubMed]
- Zentek, J.; Buchheit-Renko, S.; Manner, K.; Pieper, R.; Vahjen, W. Intestinal concentrations of free and encapsulated dietary medium-chain fatty acids and effects on gastric microbial ecology and bacterial metabolic products in the digestive tract of piglets. Arch. Anim. Nutr. 2012, 66, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Dierick, N.A.; Decuypere, J.A.; Molly, K.; Van Beek, E.; Vanderbeke, E. The combined use of triacylglycerols (TAGs) containing medium chain fatty acids (MCFAs) and exogenous lipolytic enzymes as an alternative to nutritional antibiotics in piglet nutrition—II. In Vivo release of MCFAs in gastric cannulated and slaughtered piglets by endogenous and exogenous lipases; effects on the luminal gut flora and growth performance. Livest. Prod. Sci. 2002, 76, 1–16. [Google Scholar] [CrossRef]
- Sprong, R.C.; Hulstein, M.F.; Van der Meer, R. High intake of milk fat inhibits intestinal colonization of Listeria but not of Salmonella in rats. J. Nutr. 1999, 129, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.; Boneh, R.S.; Wine, E. Evolving role of diet in the pathogenesis and treatment of inflammatory bowel diseases. Gut 2018, 67, 1726–1738. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Milard, M.; Laugerette, F.; Bugeat, S.; Plaisancie, P.; Letisse, M.; Meugnier, E.; Loizon, E.; Durand, A.; Buisson, C.; Geloen, A.; et al. Metabolic effects in mice of cream formulation: Addition of both thickener and emulsifier does not alter lipid metabolism but modulates mucus cells and intestinal endoplasmic reticulum stress. J. Dairy Sci. 2018, 101, 10649–10663. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, J.; Chen, Y.; Chen, C.; Chu, B.; Zhang, Y. p-Coumaric acid as a prophylactic measure against normobaric hypoxia induced pulmonary edema in mice. Life Sci. 2018, 211, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jangra, S.; Kuruva, R.S.; Sharma, R.K.; Pothuraju, R.; Mohanty, A.K. Ameliorative effect of fermentable fibres on adiposity and insulin resistance in C57BL/6 mice fed a high-fat and sucrose diet. Food Funct. 2019, 10, 3696–3705. [Google Scholar] [CrossRef] [PubMed]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; De Jesus-Laboy, K.M.; Shen, N.; Cox, L.M.; Amir, A.; Gonzalez, A.; Bokulich, N.A.; Song, S.J.; Hoashi, M.; Rivera-Vinas, J.I.; et al. Partial restoration of the microbiota of cesarean-born infants via vaginal microbial transfer. Nat. Med. 2016, 22, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Yamanishi, S.; Cox, L.; Methe, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, D.; Cadwell, K. Intrinsic Defense Mechanisms of the Intestinal Epithelium. Cell Host Microbe 2016, 19, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Holder, M.K.; Peters, N.V.; Whylings, J.; Fields, C.T.; Gewirtz, A.T.; Chassaing, B.; de Vries, G.J. Dietary emulsifiers consumption alters anxiety-like and social-related behaviors in mice in a sex-dependent manner. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Denis, M.C.; Roy, D.; Yeganeh, P.R.; Desjardins, Y.; Varin, T.; Haddad, N.; Amre, D.; Sane, A.T.; Garofalo, C.; Furtos, A.; et al. Apple peel polyphenols: A key player in the prevention and treatment of experimental inflammatory bowel disease. Clin. Sci. 2016, 130, 2217–2237. [Google Scholar] [CrossRef]
- Weiss, G.A.; Chassard, C.; Hennet, T. Selective proliferation of intestinal Barnesiella under fucosyllactose supplementation in mice. Br. J. Nutr. 2014, 111, 1602–1610. [Google Scholar] [CrossRef]
- Willing, B.P.; Dicksved, J.; Halfvarson, J.; Andersson, A.F.; Lucio, M.; Zheng, Z.; Järnerot, G.; Tysk, C.; Jansson, J.K.; Engstrand, L. A Pyrosequencing Study in Twins Shows That Gastrointestinal Microbial Profiles Vary with Inflammatory Bowel Disease Phenotypes. Gastroenterology 2011, 139, 1844–1854.e1. [Google Scholar] [CrossRef]
- Zhang, C.H.; Zhang, M.H.; Pang, X.Y.; Zhao, Y.F.; Wang, L.H.; Zhao, L.P. Structural resilience of the gut microbiota in adult mice under high-fat dietary perturbations. ISME J. 2012, 6, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.T.; Kong, J.Y.; Stothard, P.; Willing, B.P. Defining the role of Parasutterella, a previously uncharacterized member of the core gut microbiota. ISME J. 2019, 13, 1520–1534. [Google Scholar] [CrossRef] [PubMed]
- Kreutzer, C.; Peters, S.; Schulte, D.M.; Fangmann, D.; Turk, K.; Wolff, S.; van Eimeren, T.; Ahrens, M.; Beckmann, J.; Schafmayer, C.; et al. Hypothalamic Inflammation in Human Obesity is Mediated by Environmental and Genetic Factors. Diabetes 2017, 66, 2407–2415. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Nigro, G.; Sansonetti, P.J. Microbiota and gut stem cells cross-talks: A new view of epithelial homeostasis. Curr. Stem Cell Rep. 2015, 1, 48–52. [Google Scholar] [CrossRef]
- Rowan, F.E.; Docherty, N.G.; Coffey, J.C.; O’Connell, P.R. Sulphate-reducing bacteria and hydrogen sulphide in the aetiology of ulcerative colitis. Br. J. Surg. 2009, 96, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Portune, K.J.; Benitez-Paez, A.; Del Pulgar, E.M.G.; Cerrudo, V.; Sanz, Y. Gut microbiota, diet, and obesity-related disorders-The good, the bad, and the future challenges. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; ter Horst, R.; Jansen, T.; Jacobs, L.; Bonder, M.J.; et al. Linking the Human Gut Microbiome to Inflammatory Cytokine Production Capacity. Cell 2016, 167, 1897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.W.; Jin, G.; Yang, X.D.; Zhou, H.L. Regulating dyslipidemia effect of polysaccharides from Pleurotus ostreatus on fat-emulsion-induced hyperlipidemia rats. Int. J. Biol. Macromol. 2017, 101, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Bharosay, A.; Bharosay, V.V.; Bandyopadhyay, D.; Choubey, R.; Varma, V.; Saxena, K.; Varma, A. Neurological worsening and association between LDL, HDL ratio, mean platelet volume and platelet count in cerebrovascular ischemic stroke. Int. J. Health Sci. Res. 2016, 6, 209–215. [Google Scholar]
- Cox, C.; Mann, J.; Sutherland, W.; Chisholm, A.; Skeaff, M. Effects of Coconut Oil, Butter, and Safflower Oil on Lipids and Lipoproteins in Persons with Moderately Elevated Cholesterol Levels. J. Lipid Res. 1995, 36, 1787–1795. [Google Scholar] [CrossRef]
- Cox, C.; Sutherland, W.; Mann, J.; de Jong, S.; Chisholm, A.; Skeaff, M. Effects of dietary coconut oil, butter and safflower oil on plasma lipids, lipoproteins and lathosterol levels. Eur. J. Clin. Nutr. 1998, 52, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.S. Characterizing how Glycerol Monolaurate (GML) Affects Human T Cell Signaling and Function. Ph.D. Thesis, University of Iowa, Iowa City, IA, USA, 2018. [Google Scholar]
- Lv, X.C.; Guo, W.L.; Li, L.; Yu, X.D.; Liu, B. Polysaccharide peptides from Ganoderma lucidum ameliorate lipid metabolic disorders and gut microbiota dysbiosis in high-fat diet-fed rats. J. Funct. Foods 2019, 57, 48–58. [Google Scholar] [CrossRef]
- Hua, P.P.; Yu, Z.Y.; Xiong, Y.; Liu, B.; Zhao, L.N. Regulatory Efficacy of Spirulina platensis Protease Hydrolyzate on Lipid Metabolism and Gut Microbiota in High-Fat Diet-Fed Rats. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Li, T.T.; Tong, A.J.; Liu, Y.Y.; Huang, Z.R.; Wan, X.Z.; Pan, Y.Y.; Jia, R.B.; Liu, B.; Chen, X.H.; Zhao, C. Polyunsaturated fatty acids from microalgae Spirulina platensis modulates lipid metabolism disorders and gut microbiota in high-fat diet rats. Food Chem. Toxicol. 2019, 131, 110558. [Google Scholar] [CrossRef] [PubMed]
- Boulange, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016, 8. [Google Scholar] [CrossRef]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.Y.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome-brain-beta-cell axis to promote metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef]
- Sugimoto, K.; Ogawa, A.; Mizoguchi, E.; Shimomura, Y.; Andoh, A.; Bhan, A.K.; Blumberg, R.S.; Xavier, R.J.; Mizoguchi, A. IL-22 ameliorates intestinal inflammation in a mouse model of ulcerative colitis. J. Clin. Investig. 2008, 118, 534–544. [Google Scholar] [CrossRef]
- Waldner, M.J.; Neurath, M.F. Mechanisms of Immune Signaling in Colitis-Associated Cancer. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Dudakov, J.A.; Hanash, A.M.; van den Brink, M.R.M. Interleukin-22: Immunobiology and Pathology. Annu. Rev. Immunol. 2015, 33, 747–785. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Chen, S.S.; Chen, Y. Unraveling the biological functions of Smad7 with mouse models. Cell Biosci. 2011, 1. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | NCD | G400 | G800 | G1600 |
|---|---|---|---|---|
| TC (mmol L−1) | 3.71 ± 0.47 | 3.81 ± 0.76 | 3.91 ± 0.86 | 3.66 ± 0.53 |
| TG (mmol L−1) | 0.82 ± 0.08 | 1.21 ± 0.25 * | 1.03 ± 0.19 | 0.85 ± 0.32 |
| LDL-C (mmol L−1) | 0.93 ± 0.56 | 0.88 ± 0.37 | 0.75 ± 0.51 | 0.42 ± 0.34 |
| HDL-C (mmol L−1) | 1.75 ± 0.39 | 1.99 ± 0.38 | 1.88 ± 0.51 | 2.38 ± 0.77 |
| LDL-C/HDL-C | 0.69 ± 0.34 | 0.54 ± 0.13 | 0.56 ± 0.25 | 0.29 ± 0.06 # |
| atherogenic index | 1.20 ± 0.45 | 0.92 ± 0.20 | 1.28 ± 1.01 | 0.60 ± 0.30 |
| fasting Glu (mmol L−1) | 3.43 ± 0.72 | 4.54 ± 0.51 | 4.77 ± 1.47 | 4.39 ± 0.65 |
| insulin (mU L−1) | 7.20 ± 1.26 | 6.36 ± 0.69 | 6.93 ± 0.85 | 5.85 ± 0.53 |
| HOMA-IR score | 1.09 ± 0.27 | 1.28 ± 0.11 | 1.49 ± 0.56 | 1.14 ± 0.18 |
| adiponectin (pg mL−1) | 16.32 ± 3.23 | 13.03 ± 4.51 | 18.06 ± 2.93 | 14.13 ± 3.31 |
| leptin (pg mL−1) | 70.20 ± 16.82 | 63.51 ± 7.17 | 99.44 ± 43.24 | 97.51 ± 43.70 |
| FFA (mmol L−1) | 145.85 ± 70.79 | 89.77 ± 25.73 | 102.05 ± 20.60 | 125.48 ± 28.36 |
| GOT (U L−1) | 12.70 ± 2.46 | 14.73 ± 4.86 | 16.64 ± 4.76 | 14.40 ± 2.72 |
| GPT (U L−1) | 11.83 ± 10.26 | 15.15 ± 5.94 | 17.04 ± 2.00 | 15.52 ± 2.60 |
| AKP (U L−1) | 7.95 ± 0.85 | 6.77 ± 1.23 | 7.27 ± 1.39 | 8.76 ± 1.51 |
| Parameters | NCD | G400 | G800 | G1600 |
|---|---|---|---|---|
| TGF-β1 (ng mL−1) | 14.34 ± 6.98 | 14.08 ± 7.11 | 13.72 ± 7.25 | 23.32 ± 8.87 * |
| IL-22 (pg mL−1) | 25.42 ± 5.03 | 23.63 ± 1.60 | 21.52 ± 1.31 | 31.09 ± 2.59 * |
| IL-10 (pg mL−1) | 269.21 ± 131.29 | 369.26 ± 149.80 | 219.96 ± 183.82 | 147.65 ± 43.92 |
| TNFα (pg mL−1) | 22.23 ± 6.24 | 17.99 ± 7.98 | 19.63 ± 5.87 | 20.66 ± 7.63 |
| IL-6 (pg mL−1) | 28.95 ± 8.05 | 24.56 ± 8.01 | 29.19 ± 13.95 | 26.90 ± 12.56 |
| IL-1β (pg mL−1) | 13.20 ± 10.23 | 4.57 ± 3.80 | 8.03 ± 6.94 | 24.21 ± 12.26 |
| IL-12/p70 (pg mL−1) | 33.99 ± 9.78 | 32.10 ± 10.70 | 39.12 ± 14.85 | 37.95 ± 6.43 |
| IFNγ (pg mL−1) | 36.70 ± 2.00 | 38.97 ± 2.91 | 39.86 ± 3.33 | 40.28 ± 2.62 |
| LPS (U mL−1) | 11.18 ± 0.44 | 9.92 ± 4.72 | 11.11 ± 0.93 | 10.53 ± 0.77 |
| LBP (μg mL−1) | 1.59 ± 1.19 | 1.54 ± 0.81 | 1.55 ± 0.47 | 1.53 ± 0.58 |
| Parameters | NCD | G400 | G800 | G1600 |
|---|---|---|---|---|
| acetic acid | 98.15 ± 5.03 | 68.73 ± 12.14 ** | 64.95 ± 5.17 *** | 75.84 ± 12.55 ** |
| propionic acid | 6.97 ± 2.15 | 6.83 ± 0.64 | 7.79 ± 1.39 | 6.60 ± 0.99 |
| butyric acid | 5.93 ± 5.42 | 3.33 ± 0.93 | 6.72 ± 3.65 | 7.90 ± 4.11 |
| isobutyric acid | 0.60 ± 0.30 | 0.68 ± 0.27 | 0.79 ± 0.30 | 0.82 ± 0.27 |
| valeric acid | 0.84 ± 0.35 | 0.63 ± 0.29 | 0.83 ± 0.42 | 0.68 ± 0.33 |
| isovaleric acid | 1.19 ± 0.30 | 0.96 ± 0.17 | 1.29 ± 0.33 | 1.37 ± 0.31 |
| hexanoic acid | 0.30 ± 0.09 | 0.35 ± 0.04 | 0.36 ± 0.07 | 0.43 ± 0.18 |
| total SCFAs | 113.98 ± 12.83 | 81.50 ± 11.75 ** | 82.73 ± 8.46 ** | 93.65 ± 15.05 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, Q.; Fu, A.; Deng, L.; Zhao, M.; Li, Y.; Zhang, H.; Feng, F. High-dose Glycerol Monolaurate Up-Regulated Beneficial Indigenous Microbiota without Inducing Metabolic Dysfunction and Systemic Inflammation: New Insights into Its Antimicrobial Potential. Nutrients 2019, 11, 1981. https://doi.org/10.3390/nu11091981
Mo Q, Fu A, Deng L, Zhao M, Li Y, Zhang H, Feng F. High-dose Glycerol Monolaurate Up-Regulated Beneficial Indigenous Microbiota without Inducing Metabolic Dysfunction and Systemic Inflammation: New Insights into Its Antimicrobial Potential. Nutrients. 2019; 11(9):1981. https://doi.org/10.3390/nu11091981
Chicago/Turabian StyleMo, Qiufen, Aikun Fu, Lingli Deng, Minjie Zhao, Yang Li, Hui Zhang, and Fengqin Feng. 2019. "High-dose Glycerol Monolaurate Up-Regulated Beneficial Indigenous Microbiota without Inducing Metabolic Dysfunction and Systemic Inflammation: New Insights into Its Antimicrobial Potential" Nutrients 11, no. 9: 1981. https://doi.org/10.3390/nu11091981
APA StyleMo, Q., Fu, A., Deng, L., Zhao, M., Li, Y., Zhang, H., & Feng, F. (2019). High-dose Glycerol Monolaurate Up-Regulated Beneficial Indigenous Microbiota without Inducing Metabolic Dysfunction and Systemic Inflammation: New Insights into Its Antimicrobial Potential. Nutrients, 11(9), 1981. https://doi.org/10.3390/nu11091981