Olive Leaf Extract (OleaVita) Suppresses Inflammatory Cytokine Production and NLRP3 Inflammasomes in Human Placenta



 ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Olive Leaf Extract (OleaVita)
2.2. Tissue Collection
2.3. Experimental Conditions
2.4. Determination of Cytokines
2.5. Real-Time RT-PCR
2.6. Determination of Cytokines and Lactate Dehydrogenase (LDH)
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
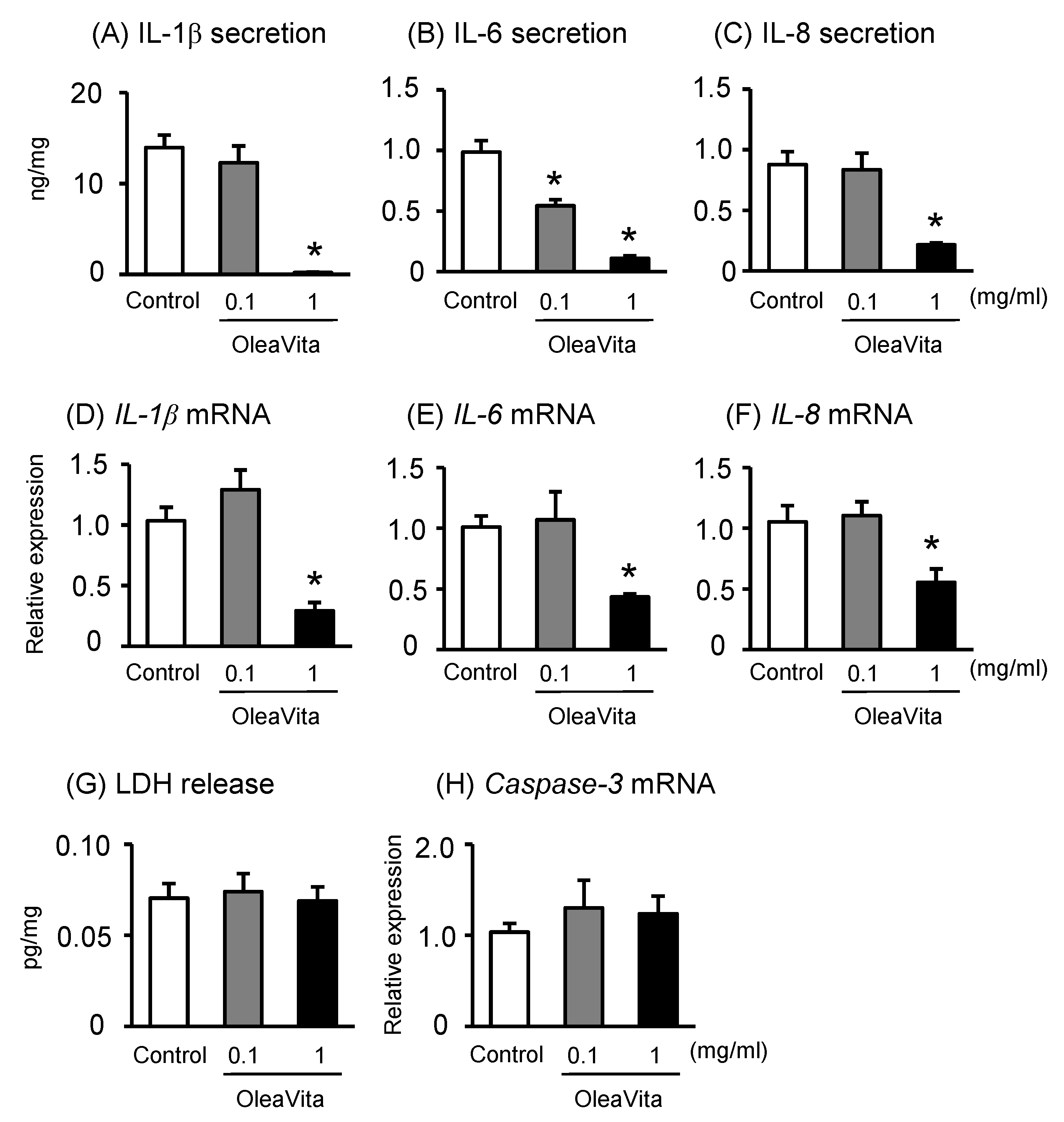
3.1. Effects of OleaVita on Inflammatory Cytokine Production in Human Placental Tissues
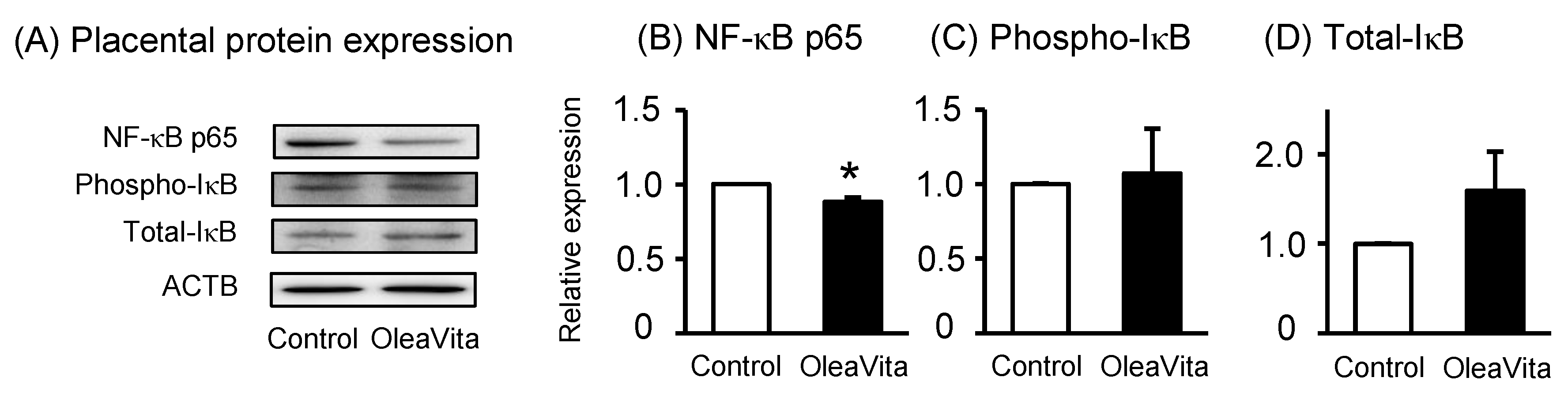
3.2. Effects of OleaVita on the NF-κB Pathway in Human Placental Tissues
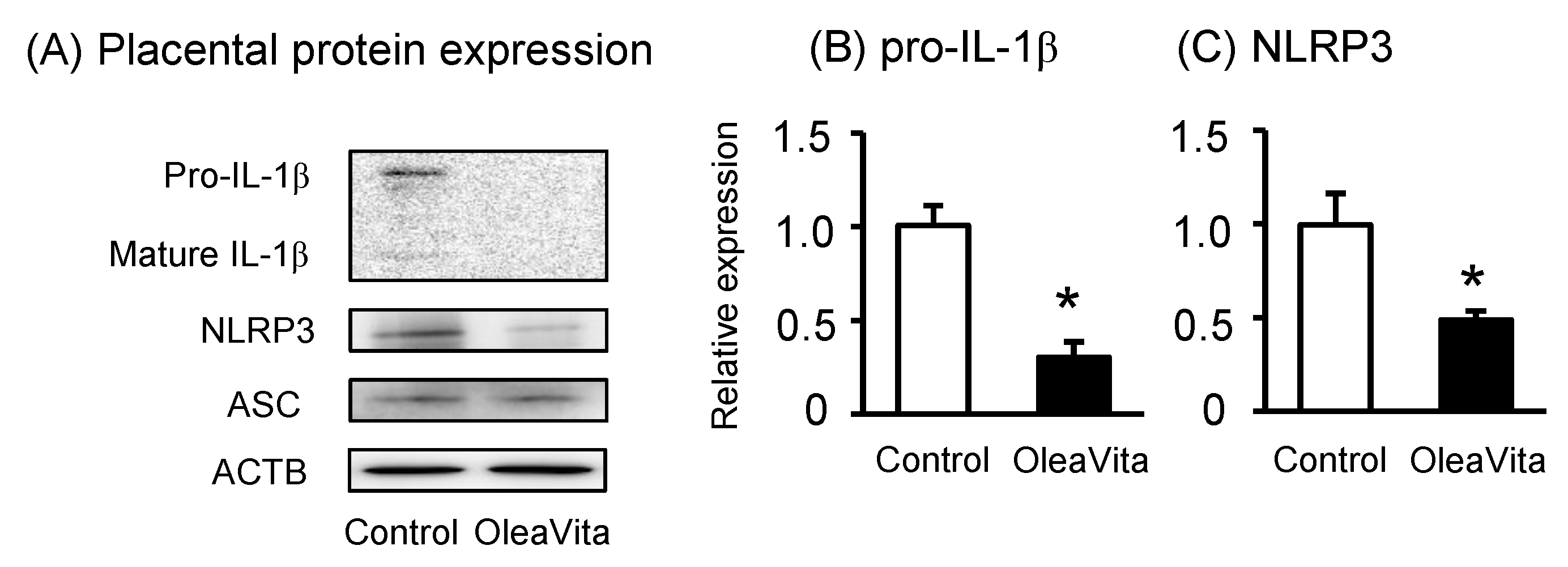
3.3. Effects of OleaVita on NLRP3 Inflammasomes in Human Placental Tissues
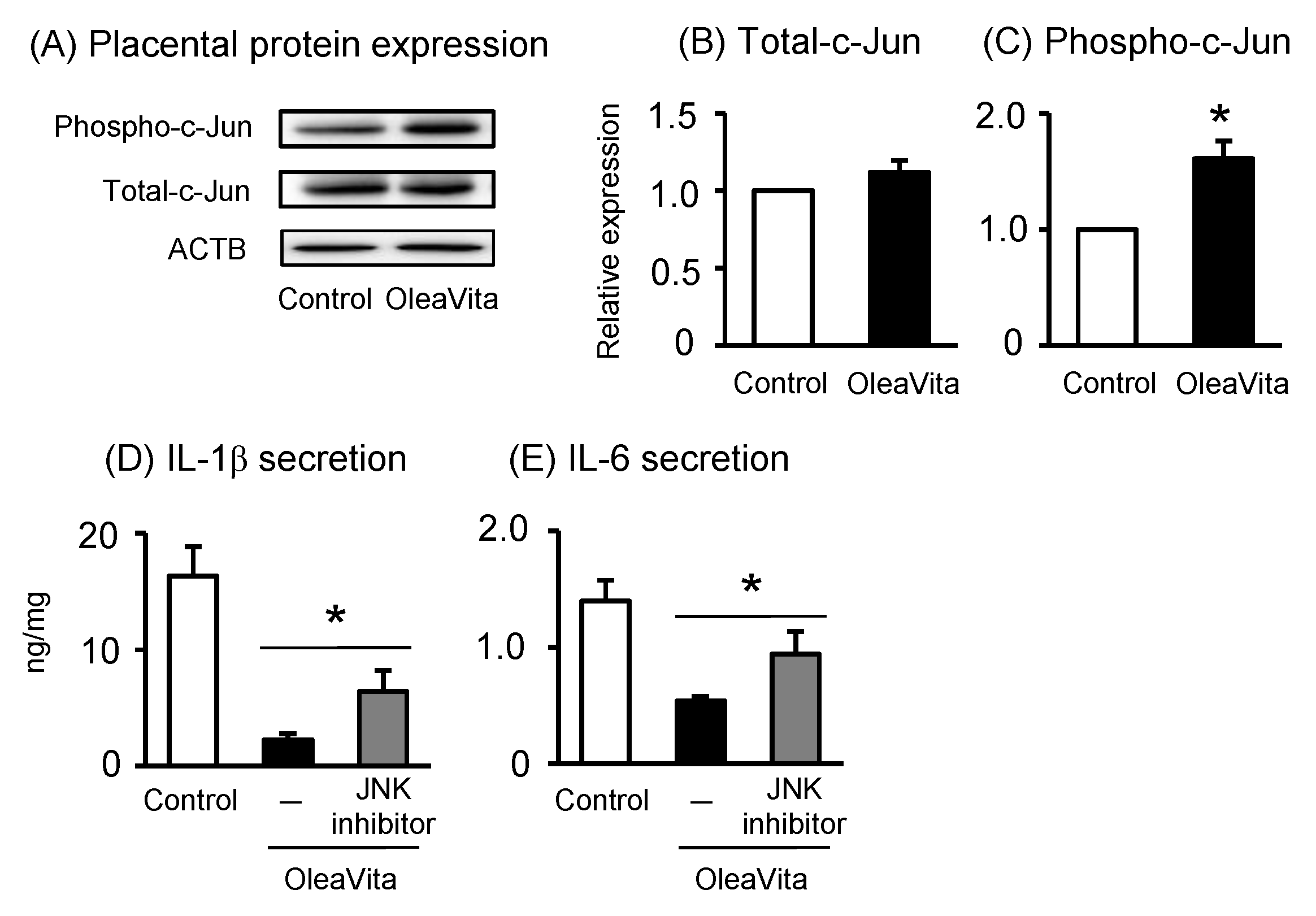
3.4. Effects of OleaVita on the JNK Pathway in Human Placental Tissues
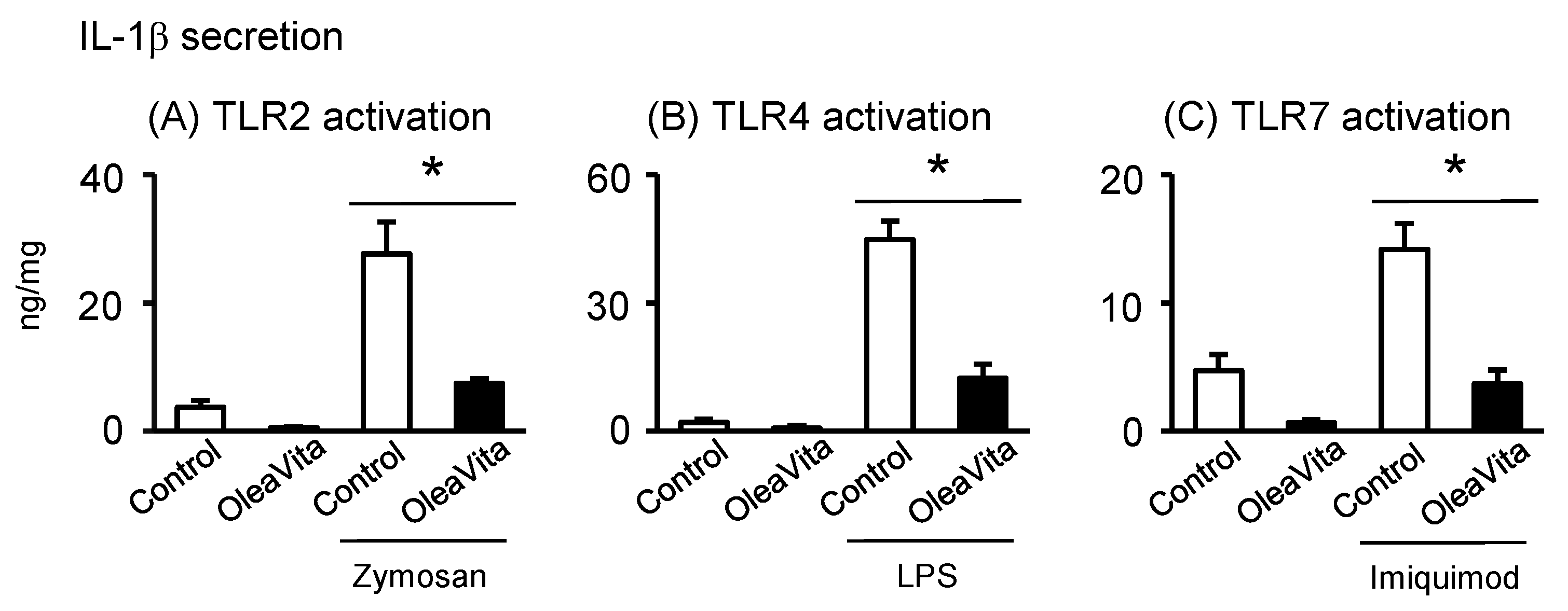
3.5. Effects of OleaVita on TLR Ligands-Induced IL-1β Secretion in Human Placental Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASC | apoptosis-associated speck-like protein containing a caspase recruitment domain |
| DRP1 | dynamin-related protein 1 |
| FCS | fetal calf serum |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| IL | interleukin |
| JNK | Jun N-terminal kinase |
| LDH | lactate dehydrogenase |
| LPS | lipopolysaccharide |
| MFN | mitofusion |
| NLR | nucleotide-binding oligomerization domain-like receptor |
| NLRP3 | NLR family pyrin domain-containing 3 |
| OPA1 | optic atrophy 1 |
| TGR5 | G protein-coupled bile acid receptor 1 |
| TLR | toll-like receptor ligand |
References
- Arck, P.C.; Hecher, K. Fetomaternal immune cross-talk and its consequences for maternal and offspring’s health. Nat. Med. 2013, 19, 548–556. [Google Scholar] [CrossRef]
- Aye, I.L.; Jansson, T.; Powell, T.L. Interleukin-1beta inhibits insulin signaling and prevents insulin-stimulated system A amino acid transport in primary human trophoblasts. Mol. Cell Endocrinol. 2013, 381, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Marchi, J.; Berg, M.; Dencker, A.; Olander, E.K.; Begley, C. Risks associated with obesity in pregnancy, for the mother and baby: A systematic review of reviews. Obes. Rev. 2015, 16, 621–638. [Google Scholar] [CrossRef]
- Catalano, P.M.; Ehrenberg, H.M. The short- and long-term implications of maternal obesity on the mother and her offspring. BJOG 2006, 113, 1126–1133. [Google Scholar] [CrossRef]
- Basu, S.; Haghiac, M.; Surace, P.; Challier, J.C.; Guerre-Millo, M.; Singh, K.; Waters, T.; Minium, J.; Presley, L.; Catalano, P.M.; et al. Pregravid obesity associates with increased maternal endotoxemia and metabolic inflammation. Obesity (Silver Spring) 2011, 19, 476–482. [Google Scholar] [CrossRef]
- Challier, J.C.; Basu, S.; Bintein, T.; Minium, J.; Hotmire, K.; Catalano, P.M.; Hauguel-de Mouzon, S. Obesity in pregnancy stimulates macrophage accumulation and inflammation in the placenta. Placenta 2008, 29, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.A.; Riley, S.C.; Reynolds, R.M.; Barr, S.; Evans, M.; Statham, A.; Hor, K.; Jabbour, H.N.; Norman, J.E.; Denison, F.C. Placental structure and inflammation in pregnancies associated with obesity. Placenta 2011, 32, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Laresgoiti-Servitje, E. A leading role for the immune system in the pathophysiology of preeclampsia. J. Leukoc. Biol. 2013, 94, 247–257. [Google Scholar] [CrossRef]
- Shirasuna, K.; Karasawa, T.; Usui, F.; Kobayashi, M.; Komada, T.; Kimura, H.; Kawashima, A.; Ohkuchi, A.; Taniguchi, S.; Takahashi, M. NLRP3 Deficiency Improves Angiotensin II-Induced Hypertension But Not Fetal Growth Restriction During Pregnancy. Endocrinology 2015, 156, 4281–4292. [Google Scholar] [CrossRef]
- Davis, B.K.; Wen, H.; Ting, J.P. The inflammasome NLRs in immunity, inflammation, and associated diseases. Annu. Rev. Immunol. 2011, 29, 707–735. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M. Role of the inflammasome in myocardial infarction. Trends Cardiovasc. Med. 2011, 21, 37–41. [Google Scholar] [CrossRef]
- Schroder, K.; Zhou, R.; Tschopp, J. The NLRP3 inflammasome: A sensor for metabolic danger? Science 2010, 327, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Nunez, G. Sterile inflammation: sensing and reacting to damage. Nat. Rev. Immunol. 2010, 10, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Weel, I.C.; Romao-Veiga, M.; Matias, M.L.; Fioratti, E.G.; Peracoli, J.C.; Borges, V.T.; Araujo, J.P., Jr.; Peracoli, M.T. Increased expression of NLRP3 inflammasome in placentas from pregnant women with severe preeclampsia. J. Reprod. Immunol. 2017, 123, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Shirasuna, K.; Usui, F.; Karasawa, T.; Kimura, H.; Kawashima, A.; Mizukami, H.; Ohkuchi, A.; Nishimura, S.; Sagara, J.; Noda, T.; et al. Nanosilica-induced placental inflammation and pregnancy complications: Different roles of the inflammasome components NLRP3 and ASC. Nanotoxicology 2015, 9, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Shirasuna, K.; Takano, H.; Seno, K.; Ohtsu, A.; Karasawa, T.; Takahashi, M.; Ohkuchi, A.; Suzuki, H.; Matsubara, S.; Iwata, H.; Kuwayama, T. Palmitic acid induces interleukin-1beta secretion via NLRP3 inflammasomes and inflammatory responses through ROS production in human placental cells. J. Reprod. Immunol. 2016, 116, 104–112. [Google Scholar] [CrossRef]
- Efentakis, P.; Iliodromitis, E.K.; Mikros, E.; Papachristodoulou, A.; Dagres, N.; Skaltsounis, A.L.; Andreadou, I. Effects of the olive tree leaf constituents on myocardial oxidative damage and atherosclerosis. Planta. Med. 2015, 81, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Elgebaly, H.A.; Mosa, N.M.; Allach, M.; El-Massry, K.F.; El-Ghorab, A.H.; Al Hroob, A.M.; Mahmoud, A.M. Olive oil and leaf extract prevent fluoxetine-induced hepatotoxicity by attenuating oxidative stress, inflammation and apoptosis. Biomed Pharmacother 2018, 98, 446–453. [Google Scholar] [CrossRef]
- Kim, M.S.; Han, J.Y.; Kim, S.H.; Jeon, D.; Kim, H.Y.; Lee, S.W.; Rho, M.C.; Lee, K. Oleanolic acid acetate attenuates polyhexamethylene guanidine phosphate-induced pulmonary inflammation and fibrosis in mice. Respir. Physiol. Neurobiol. 2018, 252, 1–9. [Google Scholar] [CrossRef]
- Djeziri, F.Z.; Belarbi, M.; Murtaza, B.; Hichami, A.; Benammar, C.; Khan, N.A. Oleanolic acid improves diet-induced obesity by modulating fat preference and inflammation in mice. Biochimie 2018, 152, 110–120. [Google Scholar] [CrossRef]
- Martin, R.; Cordova, C.; San Roman, J.A.; Gutierrez, B.; Cachofeiro, V.; Nieto, M.L. Oleanolic acid modulates the immune-inflammatory response in mice with experimental autoimmune myocarditis and protects from cardiac injury. Therapeutic implications for the human disease. J. Mol. Cell Cardiol. 2014, 72, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Yang, Z.; Sheng, J.; Liu, J.B.; Xie, Q.Y.; Zheng, W.; Chen, K. Oleanolic acid prevents cartilage degeneration in diabetic mice via PPARgamma associated mitochondrial stabilization. Biochem. Biophys. Res. Commun. 2017, 490, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, S.; Suzuki, H.; Yokozawa, M.; Uchiyama, K.; Ishikawa, M. The effect of the intake of ethanol/water extract of olive leaves (EEO) on body fat percentage and lean body mass percentage: A randomized, double-blind, placebo-controlled, parallel-group study of healthy japanese people. Pharmacometrics 2016, 91, 115–121. [Google Scholar]
- Seno, K.; Sase, S.; Ozeki, A.; Takahashi, H.; Ohkuchi, A.; Suzuki, H.; Matsubara, S.; Iwata, H.; Kuwayama, T.; Shirasuna, K. Advanced glycation end products regulate interleukin-1beta production in human placenta. J. Reprod. Dev. 2017, 63, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Brasier, A.R. The nuclear factor-kappaB-interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef]
- Kaneko, Y.; Tokyo University of Agriculture, Kanagawa, Japan. Unpublished work. 2019.
- Lou, G.; Ma, X.; Fu, X.; Meng, Z.; Zhang, W.; Wang, Y.D.; Van Ness, C.; Yu, D.; Xu, R.; Huang, W. GPBAR1/TGR5 mediates bile acid-induced cytokine expression in murine Kupffer cells. PLoS ONE 2014, 9, e93567. [Google Scholar] [CrossRef] [PubMed]
- Kawamata, Y.; Fujii, R.; Hosoya, M.; Harada, M.; Yoshida, H.; Miwa, M.; Fukusumi, S.; Habata, Y.; Itoh, T.; Shintani, Y.; et al. A G protein-coupled receptor responsive to bile acids. J. Biol. Chem. 2003, 278, 9435–9440. [Google Scholar] [CrossRef]
- Sato, H.; Genet, C.; Strehle, A.; Thomas, C.; Lobstein, A.; Wagner, A.; Mioskowski, C.; Auwerx, J.; Saladin, R. Anti-hyperglycemic activity of a TGR5 agonist isolated from Olea europaea. Biochem. Biophys. Res. Commun. 2007, 362, 793–798. [Google Scholar] [CrossRef]
- An, Q.; Hu, Q.; Wang, B.; Cui, W.; Wu, F.; Ding, Y. Oleanolic acid alleviates diabetic rat carotid artery injury through the inhibition of NLRP3 inflammasome signaling pathways. Mol. Med. Rep. 2017, 16, 8413–8419. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, J.; Song, L.; Liu, Z.; Han, G.; Yuan, D.; Wang, T.; Dun, Y.; Zhou, Z.; Liu, Z. Oleanolic acid rejuvenates testicular function through attenuating germ cell DNA damage and apoptosis via deactivation of NF-kappaB, p53 and p38 signalling pathways. J. Pharm. Pharmacol. 2017, 69, 295–304. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Normal Pregnancy (n = 12) |
|---|---|
| Age of mother | 35.4 (27–40) |
| Maximum systolic blood pressure (mmHg) | 112.3 (88–132) |
| Maximum diastolic blood pressure (mmHg) | 70.8 (48–96) |
| Delivery gestation (week) | 38.0 (37–39) |
| Placental weight (g) | 540.3 (415–627) |
| Birthweight (g) | 2712.7 (2066–3660) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaneko, Y.; Sano, M.; Seno, K.; Oogaki, Y.; Takahashi, H.; Ohkuchi, A.; Yokozawa, M.; Yamauchi, K.; Iwata, H.; Kuwayama, T.; et al. Olive Leaf Extract (OleaVita) Suppresses Inflammatory Cytokine Production and NLRP3 Inflammasomes in Human Placenta. Nutrients 2019, 11, 970. https://doi.org/10.3390/nu11050970
Kaneko Y, Sano M, Seno K, Oogaki Y, Takahashi H, Ohkuchi A, Yokozawa M, Yamauchi K, Iwata H, Kuwayama T, et al. Olive Leaf Extract (OleaVita) Suppresses Inflammatory Cytokine Production and NLRP3 Inflammasomes in Human Placenta. Nutrients. 2019; 11(5):970. https://doi.org/10.3390/nu11050970
Chicago/Turabian StyleKaneko, Yasuaki, Michiya Sano, Kotomi Seno, Yuka Oogaki, Hironori Takahashi, Akihide Ohkuchi, Miki Yokozawa, Ken Yamauchi, Hisataka Iwata, Takehito Kuwayama, and et al. 2019. "Olive Leaf Extract (OleaVita) Suppresses Inflammatory Cytokine Production and NLRP3 Inflammasomes in Human Placenta" Nutrients 11, no. 5: 970. https://doi.org/10.3390/nu11050970
APA StyleKaneko, Y., Sano, M., Seno, K., Oogaki, Y., Takahashi, H., Ohkuchi, A., Yokozawa, M., Yamauchi, K., Iwata, H., Kuwayama, T., & Shirasuna, K. (2019). Olive Leaf Extract (OleaVita) Suppresses Inflammatory Cytokine Production and NLRP3 Inflammasomes in Human Placenta. Nutrients, 11(5), 970. https://doi.org/10.3390/nu11050970