Blind Analysis of Food-Related IgG Identifies Five Possible Nutritional Clusters for the Italian Population: Future Implications for Pregnancy and Lactation


Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Statistical Methods
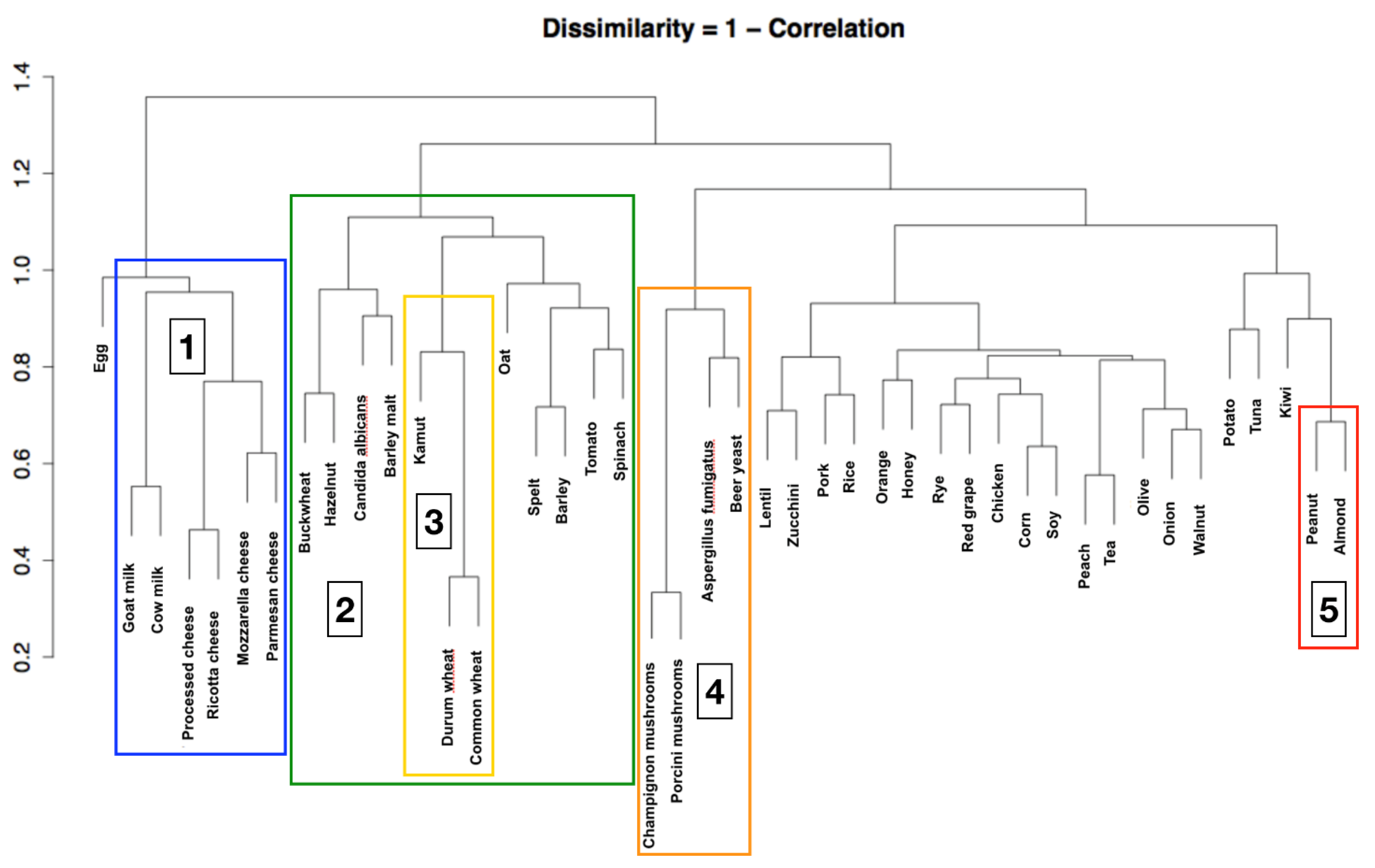
3. Results
4. Discussion
4.1. Main Findings and Interpretation
4.2. Speculations and Further Hypothesis for Future Implications in Pregnancy and Lactation
4.3. Strength and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ligaarden, S.C.; Lydersen, S.; Farup, P.G. IgG and IgG4 Antibodies in Subjects with Irritable Bowel Syndrome: A Case Control Study in the General Population. BMC Gastroenterol. 2012, 12, 166. [Google Scholar] [CrossRef]
- Speciani, A.F.; Piuri, G.; Ferrazzi, E. IgG Levels to Food Correlate with Nutritional Exposure to Food Antigens but a Methodological Weakness of this Research Prevents the Recognition of Yeast-Related Foods as a Possible Cause of Irritable Bowel Syndrome (IBS). Comment to IgG and IgG4 Antibodies in Subjects with Irritable Bowel Syndrome: A Case Control Study in the General Population Solveig C Ligaarden*, Stian Lydersen and Per G Farup BMC Gastroenterology 2012, 12:166. BMC Gastroenterol. 2012, 12, 166. [Google Scholar]
- Wisniewski, J.; Agrawal, R.; Woodfolk, J.A. Mechanisms of Tolerance Induction in Allergic Disease: Integrating Current and Emerging Concepts. Clin. Exp. Allergy 2013, 43, 164–176. [Google Scholar] [CrossRef]
- Kolfschoten, M.V.D.N.; Schuurman, J.; Losen, M.; Bleeker, W.K.; Martinez, P.M.; Vermeulen, E.; Bleker, T.H.D.; Wiegman, L.; Vink, T.; Aarden, L.A.; et al. Anti-Inflammatory Activity of Human IgG4 Antibodies by Dynamic Fab Arm Exchange. Science 2007, 317, 1554–1557. [Google Scholar] [CrossRef] [PubMed]
- Piuri, G. Individual Food Clusters Excess and Low-Grade Inflammation in Pregnancy. In Metabolic Syndrome and Complications of Pregnancy, 1st ed.; Ferrazzi, E., Sears, B., Eds.; Springer Science Publisher: New York, NY, USA; Heidelberg, Germany, 2015; pp. 23–33. [Google Scholar]
- Finkelman, F.D. Anaphylaxis: Lessons from Mouse Models. J. Allergy Clin. Immunol. 2007, 120, 506–515. [Google Scholar] [CrossRef]
- Khodoun, M.V.; Strait, R.; Armstrong, L.; Yanase, N.; Finkelman, F.D. Identification of Markers that Distinguish IgE-From IgG-Mediated Anaphylaxis. Proc. Natl. Acad. Sci. USA 2011, 108, 12413–12418. [Google Scholar] [CrossRef]
- Finkelman, F.D.; Khodoun, M.V.; Strait, R. Human IgE-Independent Systemic Anaphylaxis. J. Allergy Clin. Immunol. 2016, 137, 1674–1680. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Cano, R.; Picado, C.; Valero, A.; Bartra, J. Mechanisms of Anaphylaxis Beyond IgE. J. Investig. Allergol. Clin. Immunol. 2016, 26, 73–82. [Google Scholar] [CrossRef]
- Fritscher-Ravens, A.; Schuppan, D.; Ellrichmann, M.; Schoch, S.; Röcken, C.; Brasch, J.; Bethge, J.; Böttner, M.; Klose, J.; Milla, P.J. Confocal Endomicroscopy Shows Food-Associated Changes in the Intestinal Mucosa of Patients with Irritable Bowel Syndrome. Gastroenterology 2014, 147, 1012–1020.e4. [Google Scholar] [CrossRef]
- Bentz, S.; Hausmann, M.; Piberger, H.; Kellermeier, S.; Paul, S.; Held, L.; Falk, W.; Obermeier, F.; Fried, M.; Schölmerich, J.; et al. Clinical Relevance of IgG Antibodies against Food Antigens in Crohn’s Disease: A Double-Blind Cross-Over Diet Intervention Study. Digestion 2010, 81, 252–264. [Google Scholar] [CrossRef]
- Cai, C.; Shen, J.; Zhao, D.; Qiao, Y.; Xu, A.; Jin, S.; Ran, Z.; Zheng, Q. Serological Investigation of Food Specific Immunoglobulin G Antibodies in Patients with Inflammatory Bowel Diseases. PLoS ONE 2014, 9, e112154. [Google Scholar] [CrossRef] [PubMed]
- Alpay, K.; Ertaş, M.; Orhan, E.K.; Üstay, D.K.; Lieners, C.; Baykan, B. Diet Restriction in Migraine, Based on IgG against Foods: A Clinical Double-Blind, Randomised, Cross-Over Trial. Cephalalgia 2010, 30, 829–837. [Google Scholar] [CrossRef]
- Biesiekierski, J.R.; Newnham, E.D.; Irving, P.M.; Barrett, J.S.; Haines, M.; Doecke, J.D.; Shepherd, S.J.; Muir, J.G.; Gibson, P.R. Gluten Causes Gastrointestinal Symptoms in Subjects without Celiac Disease: A Double-Blind Randomized Placebo-Controlled Trial. Am. J. Gastroenterol. 2011, 106, 508–514. [Google Scholar] [CrossRef]
- Kim-Lee, C.; Suresh, L.; Ambrus, J.L. Gastrointestinal Disease in Sjogren’s Syndrome: Related to Food Hypersensitivities. Springerplus 2015, 4, 766. [Google Scholar] [CrossRef]
- Shulman, R.J.; Eakin, M.N.; Czyzewski, D.I.; Jarrett, M.; Ou, C.-N. Increased Gastrointestinal Permeability and Gut Inflammation in Children with Functional Abdominal Pain and Irritable Bowel Syndrome. J. Pediatr. 2008, 153, 646–650. [Google Scholar] [CrossRef]
- Clayton, F.; Fang, J.C.; Gleich, G.J.; Lucendo, A.J.; Olalla, J.M.; Vinson, L.A.; Lowichik, A.; Chen, X.; Emerson, L.; Cox, K.; et al. Eosinophilic Esophagitis in Adults is Associated with IgG4 and not Mediated by IgE. Gastroenterology 2014, 147, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A.N.; Macia, L.; Mackay, C.R. Diet, Metabolites, and “Western-Lifestyle” Inflammatory Diseases. Immunity 2014, 40, 833–842. [Google Scholar] [CrossRef]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster Analysis and Display of Genome-Wide Expression Patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed]
- Fatati, G. [Italian eating behavior: Survey 2011]. Recenti Prog. Med. 2012, 103, 225–233. [Google Scholar]
- Pala, V.; Sieri, S.; Palli, D.; Salvini, S.; Berrino, F.; Bellegotti, M.; Frasca, G.; Tumino, R.; Sacerdote, C.; Fiorini, L.; et al. Diet in the Italian EPIC Cohorts: Presentation of Data and Methodological Issues. Tumori J. 2003, 89, 594–607. [Google Scholar] [CrossRef]
- Riccioni, G.; Menna, V.; Di Ilio, C.; D’Orazio, N. Food-Intake and Nutrients Pattern in Italian Adult Male Subjects. Clin. Ther. 2004, 155, 283–286. [Google Scholar]
- Hjartåker, A.; Lagiou, A.; Slimani, N.; Lund, E.; Chirlaque, M.D.; Vasilopoulou, E.; Zavitsanos, X.; Berrino, F.; Sacerdote, C.; Ocké, M.C.; et al. Consumption of Dairy Products in the European Prospective Investigation into Cancer and Nutrition (EPIC) Cohort: Data from 35 955 24-h Dietary Recalls in 10 European countries. Public Health Nutr. 2002, 5, 1259–1271. [Google Scholar] [CrossRef] [PubMed]
- Sherman, P.W.; Flaxman, S.M. Nausea and Vomiting of Pregnancy in an Evolutionary Perspective. Am. J. Obstet. Gynecol. 2002, 186, S190–S197. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
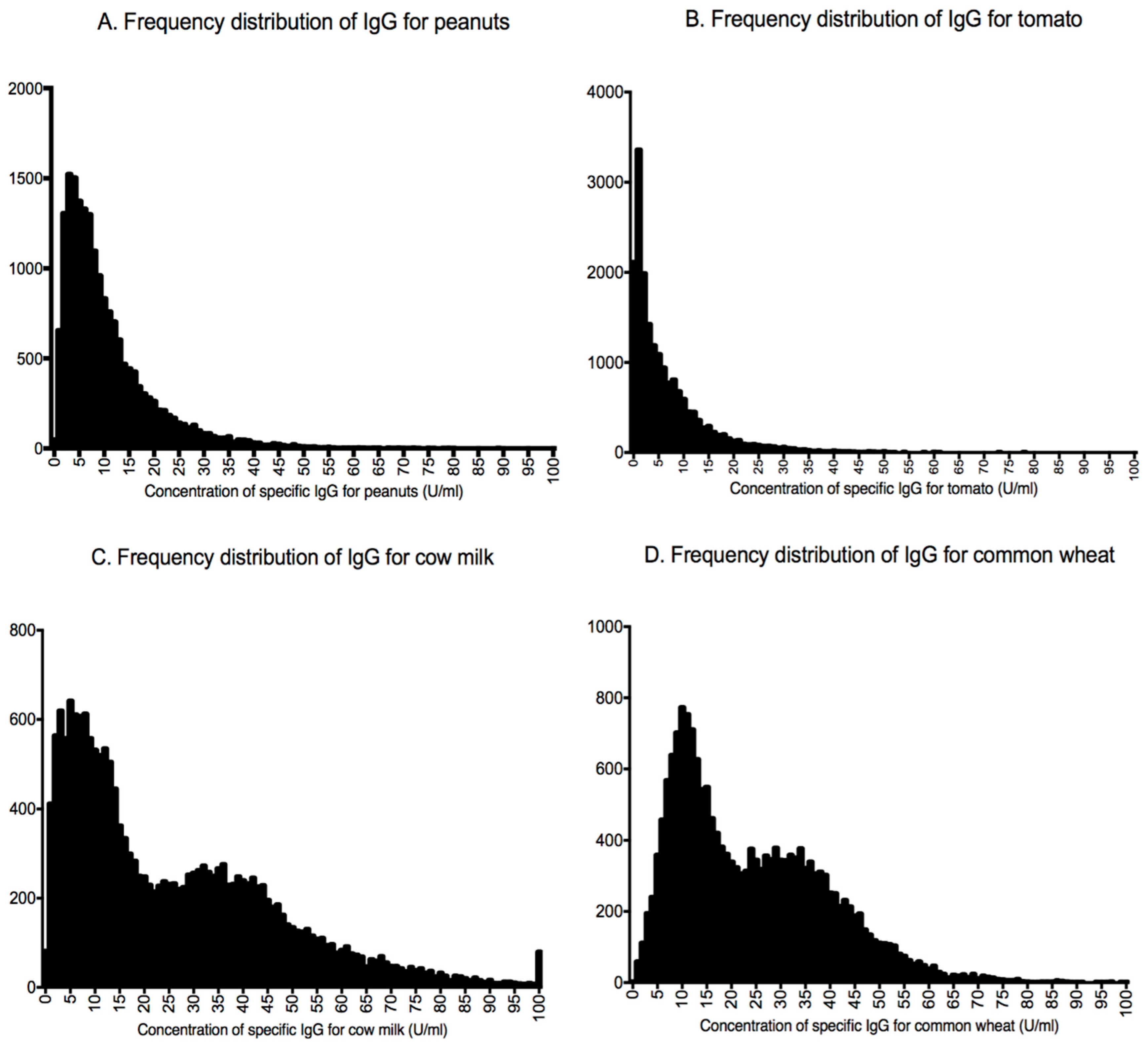
| Food–Antigen | IgG Levels U/mL | Distribution |
|---|---|---|
| Almonds | 8.35 (4.55–14.25) | Asymmetrical |
| Aspergillus fumigatus | 3.95 (1.63–10.75) | Asymmetrical |
| Barley | 9.16 (5.06–15.36) | Asymmetrical |
| Barley malt | 1.17 (0.55–2.54) | Asymmetrical |
| Buckwheat | 6.99 (3.43–12.49) | Asymmetrical |
| Candida albicans | 2.48 (0.97–7.00) | Asymmetrical |
| Canned tuna | 3.24 (1.42–6.21) | Asymmetrical |
| Champignon mushrooms | 4.05 (1.38–9.60) | Asymmetrical |
| Chicken | 0.78 (0.29–1.60) | Asymmetrical |
| Corn | 1.32 (0.67–2.51) | Asymmetrical |
| Eggs | 8.63 (3.01–21.58) | Asymmetrical |
| Goat milk | 8.18 (3.92–17.39) | Asymmetrical |
| Hazelnuts | 5.19 (2.46–9.5) | Asymmetrical |
| Honey | 0.07 (0.00–0.41) | Asymmetrical |
| Kamut | 6.47 (3.87–11.27) | Asymmetrical |
| Kiwis | 3.16 (1.55–6.72) | Asymmetrical |
| Lentils | 0.65 (0.25–1.51) | Asymmetrical |
| Mozzarella cheese | 8.71 (4.7–15.56) | Asymmetrical |
| Oats | 1.97 (0.99–4.28) | Asymmetrical |
| Olives | 0.01 (0.00–0.16) | Asymmetrical |
| Onion | 0.71 (0.32–1.40) | Asymmetrical |
| Oranges | 0.19 (0.02–0.79) | Asymmetrical |
| Parmesan cheese | 8.77 (3.45–18.72) | Asymmetrical |
| Peaches | 0.01 (0.00–0.16) | Asymmetrical |
| Peanuts | 7.83 (4.28–13.71) | Asymmetrical |
| Porcini mushrooms | 2.83 (1.1–7.08) | Asymmetrical |
| Potato | 3.34 (1.56–6.30) | Asymmetrical |
| Red grapes | 0.08 (0.00–0.50) | Asymmetrical |
| Rice | 0.52 (0.10–1.48) | Asymmetrical |
| Ricotta cheese | 13.71 (7.32–28.15) | Asymmetrical |
| Rye | 1.28 (0.61–2.65) | Asymmetrical |
| Soy | 0.97 (0.45–2.05) | Asymmetrical |
| Spinach | 0.20 (0.01–0.97) | Asymmetrical |
| Tea | 0.02 (0.00–0.19) | Asymmetrical |
| Tomato | 3.94 (1.20–9.10) | Asymmetrical |
| Walnuts | 0.41 (0.07–1.19) | Asymmetrical |
| Zucchini | 0.08 (0.00–0.60) | Asymmetrical |
| Beer yeast | 3.80 (1.87–6.81) 17.92 (9.01–30.36) | Bimodal |
| Common wheat | 11.07 (7.95–14.61) 32.53 (24.83–41.21) | Bimodal |
| Cow milk | 8.54 (4.68–13.10) 37.55 (27.60–49.70) | Bimodal |
| Durum wheat | 12.04 (7.86–16.47) 34.64 (28.39–42.74) | Bimodal |
| Pork | 1.17 (0.53–2.75) 10.74 (6.05–18.16) | Bimodal |
| Processed cheese | 14.72 (8.86–21.90) 47.19 (38.76–58.96) | Bimodal |
| Spelt | 6.42 (3.71–9.89) 23.46 (18.01–30.26) | Bimodal |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piuri, G.; Ferrazzi, E.; Speciani, A.F. Blind Analysis of Food-Related IgG Identifies Five Possible Nutritional Clusters for the Italian Population: Future Implications for Pregnancy and Lactation. Nutrients 2019, 11, 1096. https://doi.org/10.3390/nu11051096
Piuri G, Ferrazzi E, Speciani AF. Blind Analysis of Food-Related IgG Identifies Five Possible Nutritional Clusters for the Italian Population: Future Implications for Pregnancy and Lactation. Nutrients. 2019; 11(5):1096. https://doi.org/10.3390/nu11051096
Chicago/Turabian StylePiuri, Gabriele, Enrico Ferrazzi, and Attilio Francesco Speciani. 2019. "Blind Analysis of Food-Related IgG Identifies Five Possible Nutritional Clusters for the Italian Population: Future Implications for Pregnancy and Lactation" Nutrients 11, no. 5: 1096. https://doi.org/10.3390/nu11051096
APA StylePiuri, G., Ferrazzi, E., & Speciani, A. F. (2019). Blind Analysis of Food-Related IgG Identifies Five Possible Nutritional Clusters for the Italian Population: Future Implications for Pregnancy and Lactation. Nutrients, 11(5), 1096. https://doi.org/10.3390/nu11051096