Natural Isotope Abundances of Carbon and Nitrogen in Tissue Proteins and Amino Acids as Biomarkers of the Decreased Carbohydrate Oxidation and Increased Amino Acid Oxidation Induced by Caloric Restriction under a Maintained Protein Intake in Obese Rats

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet and Animals
2.2. Sample Preparation
2.3. Elemental and Isotope Determinations
2.4. Gene Expression
2.5. Statistical Analysis
3. Results
3.1. Body Weight and Composition
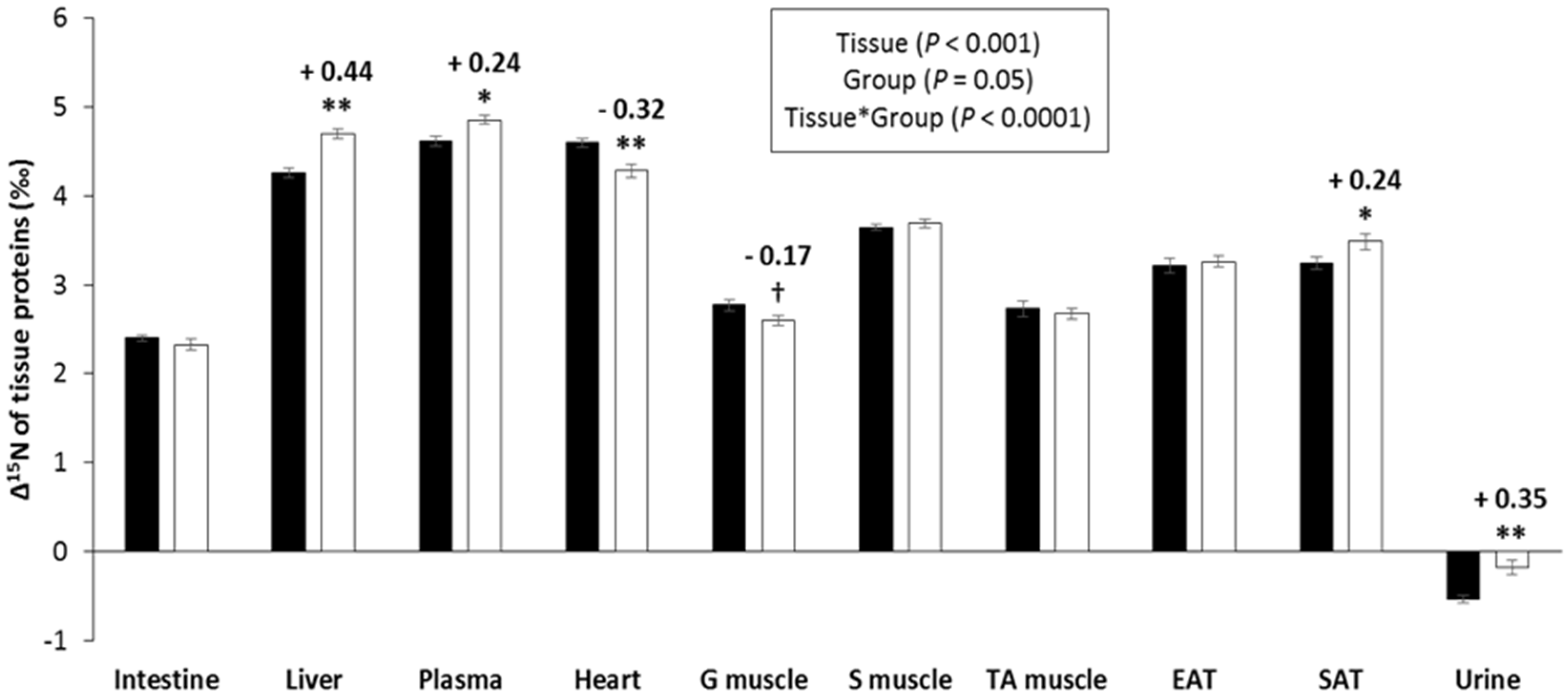
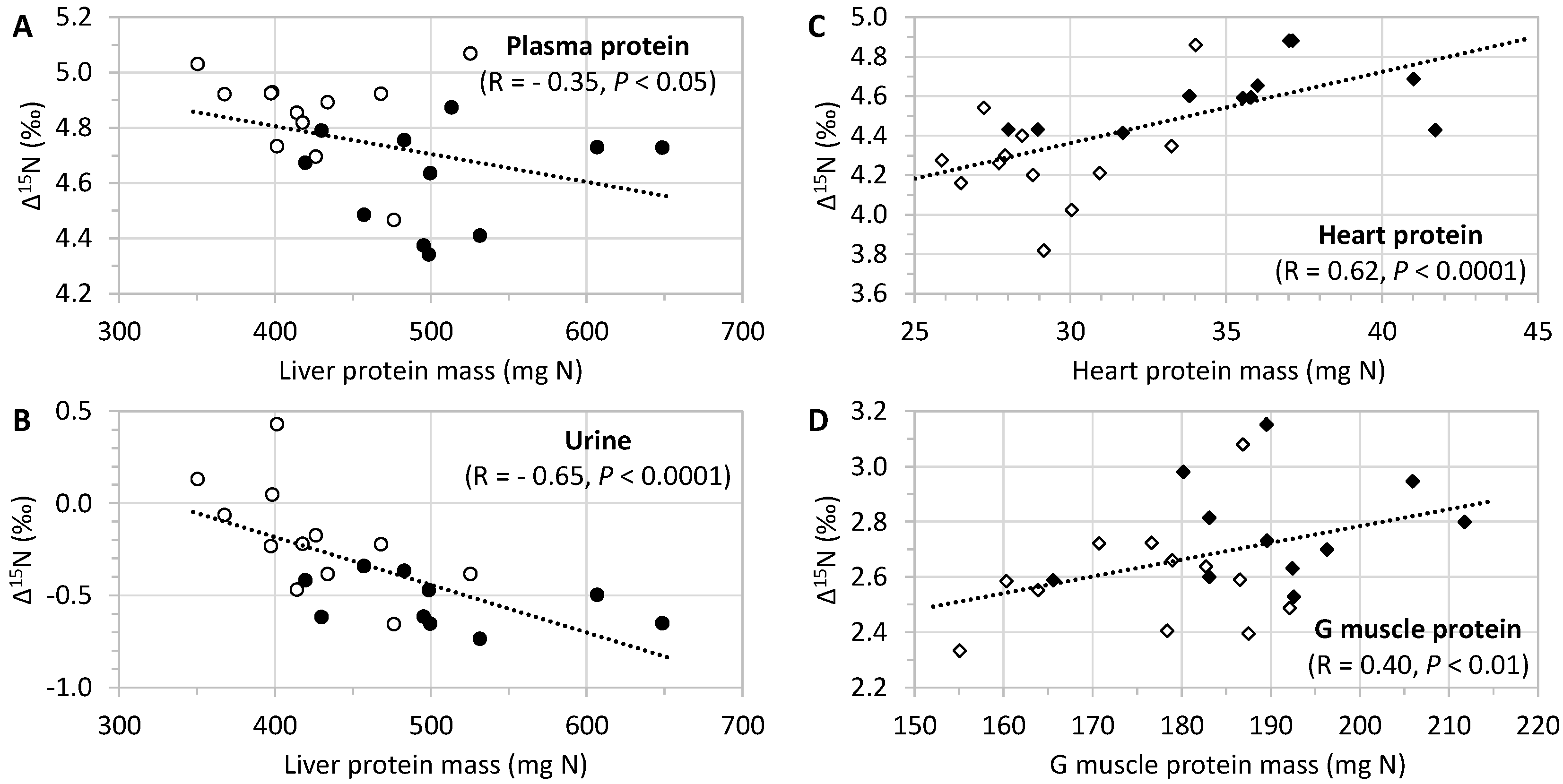
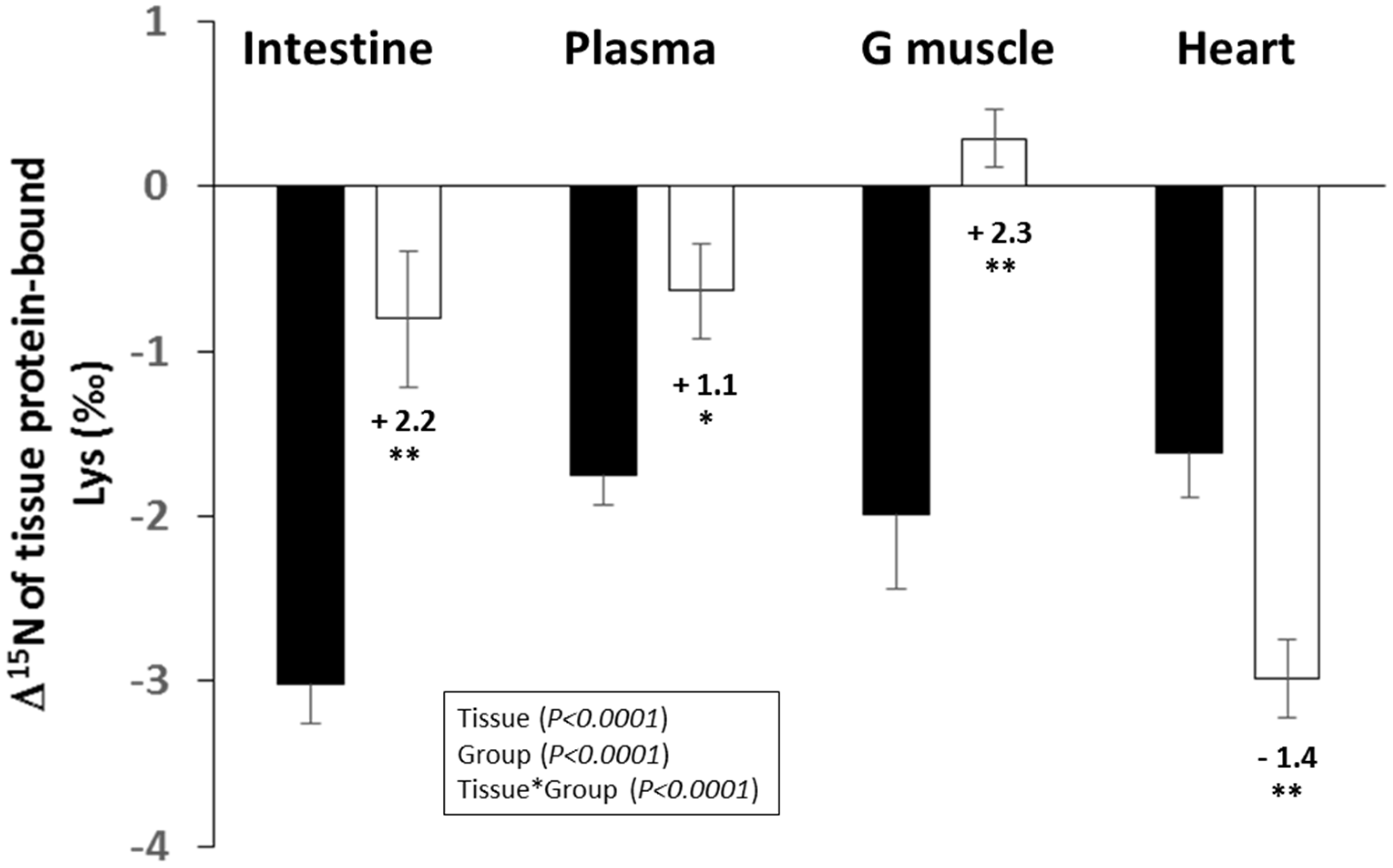
3.2. Δ15N Values in Tissue Protein and AA
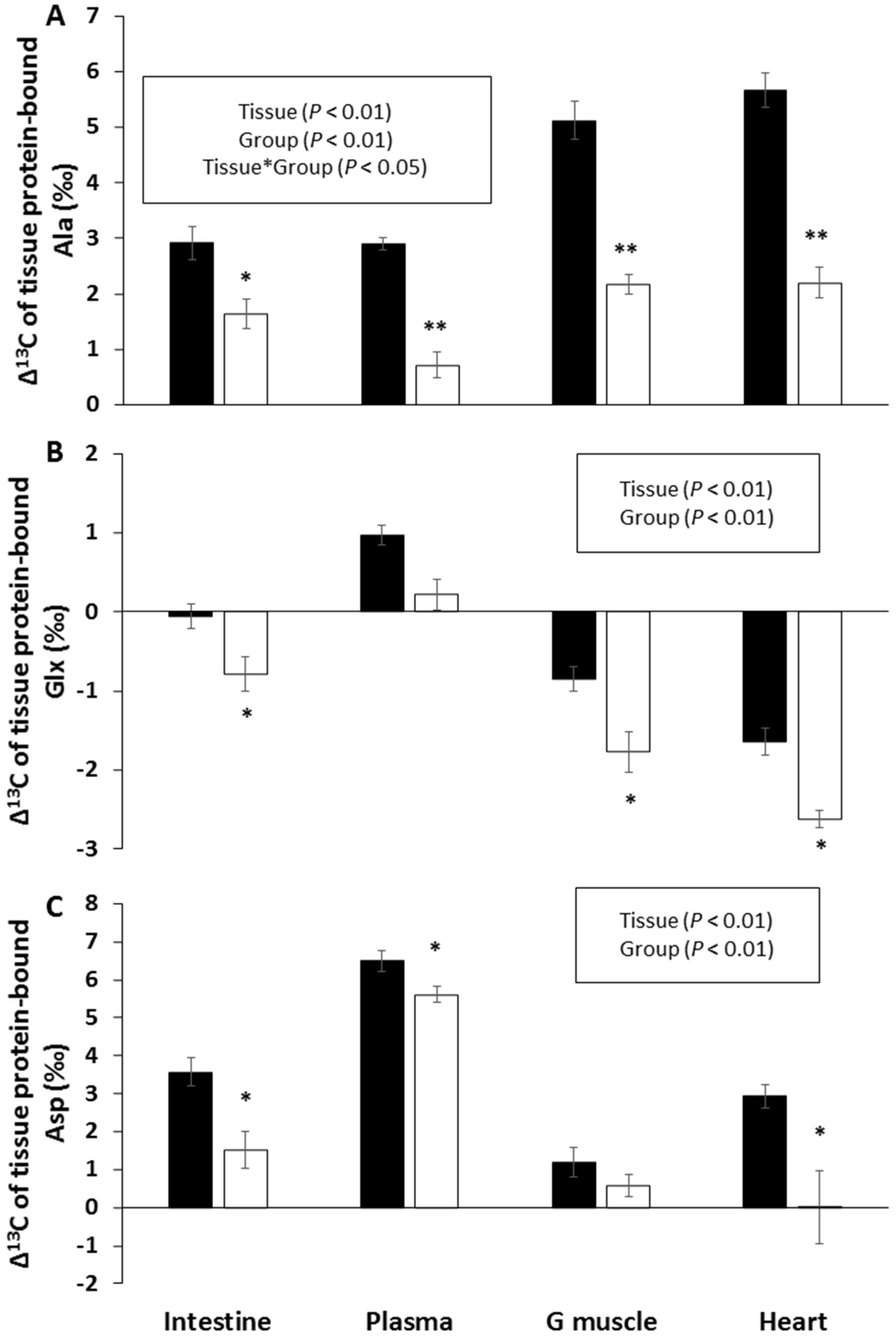
3.3. Δ13C Values in Tissue Protein and AA
3.4. Expression of Enzymes Involved in Liver AA Metabolism
4. Discussion
4.1. CR Effects on Body Composition and AA Oxidation
4.2. Tissue Δ15N Are Fingerprints of CR-Induced Effects on Tissue AA Allocation to Oxidation
4.3. Effects of CR on Dietary CHO Routing to Tissue Proteins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Brien, D.M. Stable isotope ratios as biomarkers of diet for health research. Annu. Rev. Nutr. 2015, 35, 565–594. [Google Scholar] [CrossRef]
- Nash, S.H.; Kristal, A.R.; Hopkins, S.E.; Boyer, B.B.; O’Brien, D.M. Stable isotope models of sugar intake using hair, red blood cells, and plasma, but not fasting plasma glucose, predict sugar intake in a yup’ik study population. J. Nutr. 2014, 144, 75–80. [Google Scholar] [CrossRef]
- Choy, K.; Nash, S.H.; Kristal, A.R.; Hopkins, S.; Boyer, B.B.; O’Brien, D.M. The carbon isotope ratio of alanine in red blood cells is a new candidate biomarker of sugar-sweetened beverage intake. J. Nutr. 2013, 143, 878–884. [Google Scholar] [CrossRef]
- Patel, P.S.; Cooper, A.J.M.; O’Connell, T.C.; Kuhnle, G.G.C.; Kneale, C.K.; Mulligan, A.M.; Luben, R.N.; Brage, S.; Khaw, K.T.; Wareham, N.J.; et al. Serum carbon and nitrogen stable isotopes as potential biomarkers of dietary intake and their relation with incident type 2 diabetes: The epic-norfolk study. Am. J. Clin. Nutr. 2014, 100, 708–718. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, D.M.; Kristal, A.R.; Nash, S.H.; Hopkins, S.E.; Luick, B.R.; Stanhope, K.L.; Havel, P.J.; Boyer, B.B. A stable isotope biomarker of marine food intake captures associations between n-3 fatty acid intake and chronic disease risk in a yup’ik study population, and detects new associations with blood pressure and adiponectin. J. Nutr. 2014, 144, 706–713. [Google Scholar] [CrossRef]
- Fakhouri, T.H.I.; Jahren, A.H.; Appel, L.J.; Chen, L.W.; Alavi, R.; Anderson, C.A.M. Serum carbon isotope values change in adults in response to changes in sugar-sweetened beverage intake. J. Nutr. 2014, 144, 902–905. [Google Scholar] [CrossRef][Green Version]
- McCutchan, J.H.; Lewis, W.M.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Paneth, P.; Oleary, M.H. Carbon isotope effect on dehydration of bicarbonate ion catalyzed by carbonic-anhydrase. Biochemistry 1985, 24, 5143–5147. [Google Scholar] [CrossRef]
- Macko, S.A.; Estep, M.L.F.; Engel, M.H.; Hare, P.E. Kinetic fractionation of stable nitrogen isotopes during amino-acid transamination. Geochim. Cosmochim. Acta 1986, 50, 2143–2146. [Google Scholar] [CrossRef]
- Poupin, N.; Mariotti, F.; Huneau, J.F.; Hermier, D.; Fouillet, H. Natural isotopic signatures of variations in body nitrogen fluxes: A compartmental model analysis. PLoS Comput. Biol. 2014, 10, e1003865. [Google Scholar] [CrossRef] [PubMed]
- Martinez del Rio, C.; Wolf, B.O. Mass-balance models for animal isotopic ecology. In Physiological and Ecological Adaptations to Feeding in Vertebrates; Starck, J.M., Wang, T., Eds.; Science Publishers, Inc.: Enfield, NH, USA, 2005; pp. 141–174. [Google Scholar]
- Fernandes, R.; Nadeau, M.J.; Grootes, P.M. Macronutrient-based model for dietary carbon routing in bone collagen and bioapatite. Archaeol. Anthropol. Sci. 2012, 4, 291–301. [Google Scholar] [CrossRef]
- Kurle, C.M.; Koch, P.L.; Tershy, B.R.; Croll, D.A. The effects of sex, tissue type, and dietary components on stable isotope discrimination factors (delta c-13 and delta n-15) in mammalian omnivores. Isotopes. Environ. Health Stud. 2014, 50, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Reitsema, L.J. Beyond diet reconstruction: Stable isotope applications to human physiology, health, and nutrition. Am. J. Hum. Biol. 2013, 25, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Petzke, K.J.; Fuller, B.T.; Metges, C.C. Advances in natural stable isotope ratio analysis of human hair to determine nutritional and metabolic status. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Hobson, K.A.; Alisauskas, R.T.; Clark, R.G. Stable-nitrogen isotope enrichment in avian-tissues due to fasting and nutritional stress—Implications for isotopic analyses of diet. Condor 1993, 95, 388–394. [Google Scholar] [CrossRef]
- Barboza, P.S.; Parker, K.L. Body protein stores and isotopic indicators of n balance in female reindeer (rangifer tarandus) during winter. Physiol. Biochem. Zool. 2006, 79, 628–644. [Google Scholar] [CrossRef]
- Deschner, T.; Fuller, B.T.; Oelze, V.M.; Boesch, C.; Hublin, J.J.; Mundry, R.; Richards, M.P.; Ortmann, S.; Hohmann, G. Identification of energy consumption and nutritional stress by isotopic and elemental analysis of urine in bonobos (pan paniscus). Rapid Commun. Mass Spectrom. 2012, 26, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.N.; Buck, C.L.; Barnes, B.M.; O’Brien, D.M. A test of alternative models for increased tissue nitrogen isotope ratios during fasting in hibernating arctic ground squirrels. J. Exp. Biol. 2012, 215, 3354–3361. [Google Scholar] [CrossRef] [PubMed]
- Fuller, B.T.; Fuller, J.L.; Sage, N.E.; Harris, D.A.; O’Connell, T.C.; Hedges, R.E.M. Nitrogen balance and delta n-15: Why you’re not what you eat during pregnancy. Rapid Commun. Mass Spectrom. 2004, 18, 2889–2896. [Google Scholar] [CrossRef]
- Hatch, K.A.; Crawford, M.A.; Kunz, A.W.; Thomsen, S.R.; Eggett, D.L.; Nelson, S.T.; Roeder, B.L. An objective means of diagnosing anorexia nervosa and bulimia nervosa using n-15/n-14 and c-13/c-12 ratios in hair. Rapid Commun. Mass Spectrom. 2006, 20, 3367–3373. [Google Scholar] [CrossRef]
- Mekota, A.M.; Grupe, G.; Ufer, S.; Cuntz, U. Serial analysis of stable nitrogen and carbon isotopes in hair: Monitoring starvation and recovery phases of patients suffering from anorexia nervosa. Rapid Commun. Mass Spectrom. 2006, 20, 1604–1610. [Google Scholar] [CrossRef]
- Butz, D.E.; Cook, M.E.; Eghbalnia, H.R.; Assadi-Porter, F.; Porter, W.P. Changes in the natural abundance of (co2)-c-13/(co2)-c-12 in breath due to lipopolysacchride-induced acute phase response. Rapid Commun. Mass Spectrom. 2009, 23, 3729–3735. [Google Scholar] [CrossRef]
- Whigham, L.D.; Butz, D.E.; Johnson, L.K.; Schoeller, D.A.; Abbott, D.H.; Porter, W.P.; Cook, M.E. Breath carbon stable isotope ratios identify changes in energy balance and substrate utilization in humans. Int. J. Obes. 2014, 38, 1248–1250. [Google Scholar] [CrossRef]
- Gannes, L.Z.; del Rio, C.M.; Koch, P. Natural abundance variations in stable isotopes and their potential uses in animal physiological ecology. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1998, 119, 725–737. [Google Scholar] [CrossRef]
- Neuberger, F.M.; Jopp, E.; Graw, M.; Puschel, K.; Grupe, G. Signs of malnutrition and starvation-reconstruction of nutritional life histories by serial isotopic analyses of hair. Forensic Sci. Int. 2013, 226, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Mantha, O.L.; Polakof, S.; Huneau, J.F.; Mariotti, F.; Poupin, N.; Zalko, D.; Fouillet, H. Early changes in tissue amino acid metabolism and nutrient routing in rats fed a high-fat diet: Evidence from natural isotope abundances of nitrogen and carbon in tissue proteins. Br. J. Nutr. 2018, 119, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Galmiche, G.; Huneau, J.F.; Mathe, V.; Mourot, J.; Simon, N.; Le Guillou, C.; Hermier, D. N-3 fatty acids preserve muscle mass and insulin sensitivity in a rat model of energy restriction. Br. J. Nutr. 2016, 116, 1141–1152. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Walsh, R.G.; He, S.N.; Yarnes, C.T. Compound-specific delta c-13 and delta n-15 analysis of amino acids: A rapid, chloroformate-based method for ecological studies. Rapid Commun. Mass Spectrom. 2014, 28, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Carbone, J.W.; McClung, J.P.; Pasiakos, S.M. Skeletal muscle responses to negative energy balance: Effects of dietary protein. Adv. Nutr. 2012, 3, 119–126. [Google Scholar] [CrossRef]
- Johnson, J.A.; Yu, C.H.; Yang, M.U.; Pi-Sunyer, F.X. Effect of age on protein conservation during very-low-energy diet in obese sprague-dawley rats. Obes. Res. 1998, 6, 448–457. [Google Scholar] [CrossRef]
- Chevalier, L.; Bos, C.; Azzout-Marniche, D.; Fromentin, G.; Mosoni, L.; Hafnaoui, N.; Piedcoq, J.; Tome, D.; Gaudichon, C. Energy restriction only slightly influences protein metabolism in obese rats, whatever the level of protein and its source in the diet. Int. J. Obes. 2013, 37, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.F.; Robinson, M.M.; Reuland, D.J.; Drake, J.C.; Peelor, F.F.; Bruss, M.D.; Hellerstein, M.K.; Hamilton, K.L. Calorie restriction does not increase short-term or long-term protein synthesis. J. Gerontol. A-Biol. 2013, 68, 530–538. [Google Scholar] [CrossRef]
- Zangarelli, A.; Chanseaume, E.; Morio, B.; Brugere, C.; Mosoni, L.; Rousset, P.; Giraudet, C.; Patrac, V.; Gachon, P.; Boirie, Y.; et al. Synergistic effects of caloric restriction with maintained protein intake on skeletal muscle performance in 21-month-old rats: A mitochondria-mediated pathway. FASEB J. 2006, 20, 2439–2450. [Google Scholar] [CrossRef]
- Faure, M.; Glomot, F.; Papet, I. Branched-chain amino acid aminotransferase activity decreases during development in skeletal muscles of sheep. J. Nutr. 2001, 131, 1528–1534. [Google Scholar] [CrossRef]
- Yang, Q.; Birkhahn, R.H. Branched-chain transaminase and keto acid dehydrogenase activities in burned rats: Evidence for a differential adaptation according to sex. Nutrition 1997, 13, 640–645. [Google Scholar] [CrossRef]
- Hagopian, K.; Ramsey, J.J.; Weindruch, R. Caloric restriction increases gluconeogenic and transaminase enzyme activities in mouse liver. Exp. Gerontol. 2003, 38, 267–278. [Google Scholar] [CrossRef]
- Owen, O.E.; Smalley, K.J.; D’Alessio, D.A.; Mozzoli, M.A.; Dawson, E.K. Protein, fat, and carbohydrate requirements during starvation: Anaplerosis and cataplerosis. Am. J. Clin. Nutr. 1998, 68, 12–34. [Google Scholar] [CrossRef] [PubMed]
- Jousse, C.; Averous, J.; Bruhat, A.; Carraro, V.; Mordier, S.; Fafournoux, P. Amino acids as regulators of gene expression: Molecular mechanisms. Biochem. Biophys. Res. Commun. 2004, 313, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra-Hijar, G.; Ortigues-Marty, I.; Sepchat, B.; Agabriel, J.; Huneau, J.F.; Fouillet, H. Diet-animal fractionation of nitrogen stable isotopes reflects the efficiency of nitrogen assimilation in ruminants. Br. J. Nutr. 2015, 113, 1158–1169. [Google Scholar] [CrossRef]
- Handley, L.L.; Raven, J.A. The use of natural abundance of nitrogen isotopes in plant physiology and ecology. Plant Cell Environ. 1992, 15, 965–985. [Google Scholar] [CrossRef]
- Benevenga, N.J.; Blemings, K.P. Unique aspects of lysine nutrition and metabolism. J. Nutr. 2007, 137, 1610s–1615s. [Google Scholar] [CrossRef]
- Webb, J.G.; Kiess, M.C.; Chanyan, C.C. Malnutrition and the heart. Can. Med. Assoc. J. 1986, 135, 753–758. [Google Scholar]
- Zheng, Q.J.; Zhao, K.; Han, X.F.; Huff, A.F.; Cui, Q.; Babcock, S.A.; Yu, S.Q.; Zhang, Y.M. Inhibition of ampk accentuates prolonged caloric restriction-induced change in cardiac contractile function through disruption of compensatory autophagy. BBA-Mol. Basis Dis. 2015, 1852, 332–342. [Google Scholar] [CrossRef] [PubMed]
- MacAvoy, S.E.; Arneson, L.S.; Bassett, E. Correlation of metabolism with tissue carbon and nitrogen turnover rate in small mammals. Oecologia 2006, 150, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Wolf, N.; Newsome, S.D.; Peters, J.; Fogel, M.L. Variability in the routing of dietary proteins and lipids to consumer tissues influences tissue-specific isotopic discrimination. Rapid Commun. Mass Spectrom. 2015, 29, 1448–1456. [Google Scholar] [CrossRef]
- Arneson, L.S.; MacAvoy, S.E. Carbon, nitrogen, and sulfur diet–tissue discrimination in mouse tissues. Can. J. Zool. 2005, 83, 989–995. [Google Scholar] [CrossRef]
- Kraeer, K.; Arneson, L.S.; MacAvoy, S.E. The intraspecies relationship between tissue turnover and metabolic rate in rats. Ecol. Res. 2014, 29, 937–947. [Google Scholar] [CrossRef]
- Kurle, C.M. Interpreting temporal variation in omnivore foraging ecology via stable isotope modelling. Funct. Ecol. 2009, 23, 733–744. [Google Scholar] [CrossRef]
- MacAvoy, S.E.; Macko, S.A.; Arneson, L.S. Growth versus metabolic tissue replacement in mouse tissues determined by stable carbon and nitrogen isotope analysis. Can. J. Zool. 2005, 83, 631–641. [Google Scholar] [CrossRef]
- Palmer, T.N.; Caldecourt, M.A.; Snell, K.; Sugden, M.C. Alanine and inter-organ relationships in branched-chain amino and 2-oxo acid metabolism. Biosci. Rep. 1985, 5, 1015–1033. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Caloric Restriction Diets * | |||
|---|---|---|---|---|
| Ad libitum Diet | 75% Restriction | 60% Restriction | 50% Restriction | |
| Nutrients composition (% of diet weight) | ||||
| Casein HCl | 19.4 | 25.6 | 32.4 | 35.4 |
| L-Cystine | 0.3 | 0.3 | 0.4 | 0.4 |
| Corn starch | 25.5 | 20.1 | 14.1 | 11.6 |
| Sucrose | 14.4 | 11.4 | 8.0 | 6.5 |
| High-Oleic sunflower oil | 27.0 | 27.1 | 27.2 | 27.2 |
| Soybean oil | 2.0 | 2.1 | 2.2 | 2.3 |
| AIN 93N mineral mix | 4.8 | 6.3 | 8.0 | 8.7 |
| AIN 93VX vitamin mix | 1.4 | 1.8 | 2.3 | l2.5 |
| Alpha-cellulose | 5.0 | 5.0 | 5.0 | 5.0 |
| Choline bitartrate | 0.2 | 0.3 | 0.4 | 0.4 |
| Energy composition (% of diet energy) | ||||
| Protein | 16 | 21 | 27 | 30 |
| CHO | 32 | 26 | 18 | 15 |
| Lipids | 52 | 53 | 55 | 55 |
| Isotope composition (‰) ** | ||||
| δ15N | 5.8 | 5.8 | 5.8 | 5.8 |
| δ13C | −22.3 | −22.6 | −22.8 | −23.0 |
| AL (n = 11) | CR (n = 12) | CR vs. AL | |||
|---|---|---|---|---|---|
| Whole body weight (g) | 602 ± 20 | 482 ± 10 | −21% | ** | |
| Adiposity (%) | 29.0 ± 0.7 | 23.6 ± 0.7 | −19% | ** | |
| Epididymal adipose tissue | Tissue mass (g) | 26.7 ± 1.5 | 15.1 ± 1.1 | −43% | ** |
| Protein mass (mg N) | 15.3 ± 0.8 | 10.4 ± 1.0 | −32% | ** | |
| Subcutaneous adipose tissue | Tissue mass (g) | 148.1 ± 8.3 | 99.4 ± 4.3 | −33% | ** |
| Protein mass (mg N) | 71.0 ± 6.1 | 52.0 ± 4.6 | −27% | * | |
| Small Intestine | Tissue mass (g) | 6.43 ± 0.21 | 5.15 ± 0.10 | −20% | ** |
| Protein mass (mg N) | 173.7 ± 6.0 | 142.4 ± 4.9 | −18% | ** | |
| Liver | Tissue mass (g) | 12.71 ± 0.57 | 11.14 ± 0.29 | −12% | * |
| Protein mass (mg N) | 507.5 ± 20.8 | 422.9 ± 13.9 | −17% | ** | |
| Heart | Tissue mass (g) | 1.07 ± 0.03 | 0.92 ± 0.02 | −14% | ** |
| Protein mass (mg N) | 35.1 ± 1.3 | 29.1 ± 0.7 | −17% | ** | |
| Gastrocnemius muscle | Tissue mass (g) | 5.57 ± 0.08 | 5.31 ± 0.13 | ||
| Protein mass (mg N) | 190.0 ± 3.8 | 176.6 ± 3.4 | −7% | * | |
| Tibialis anterior muscle | Tissue mass (g) | 1.80 ± 0.04 | 1.71 ± 0.04 | ||
| Protein mass (mg N) | 60.4 ± 1.5 | 59.0 ± 1.6 | |||
| Soleus muscle | Tissue mass (g) | 0.39 ± 0.01 | 0.36 ± 0.01 | ||
| Protein mass (mg N) | 14.3 ± 0.5 | 12.8 ± 0.4 | −10% | * | |
| Total | Tissue mass (g) | 202.7 ± 9.9 | 139.1 ± 5.5 | −31% | ** |
| Protein mass (mg N) | 1067 ± 30 | 905 ± 19 | −15% | ** | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huneau, J.-F.; Mantha, O.L.; Hermier, D.; Mathé, V.; Galmiche, G.; Mariotti, F.; Fouillet, H. Natural Isotope Abundances of Carbon and Nitrogen in Tissue Proteins and Amino Acids as Biomarkers of the Decreased Carbohydrate Oxidation and Increased Amino Acid Oxidation Induced by Caloric Restriction under a Maintained Protein Intake in Obese Rats. Nutrients 2019, 11, 1087. https://doi.org/10.3390/nu11051087
Huneau J-F, Mantha OL, Hermier D, Mathé V, Galmiche G, Mariotti F, Fouillet H. Natural Isotope Abundances of Carbon and Nitrogen in Tissue Proteins and Amino Acids as Biomarkers of the Decreased Carbohydrate Oxidation and Increased Amino Acid Oxidation Induced by Caloric Restriction under a Maintained Protein Intake in Obese Rats. Nutrients. 2019; 11(5):1087. https://doi.org/10.3390/nu11051087
Chicago/Turabian StyleHuneau, Jean-François, Olivier L. Mantha, Dominique Hermier, Véronique Mathé, Guillaume Galmiche, François Mariotti, and Hélène Fouillet. 2019. "Natural Isotope Abundances of Carbon and Nitrogen in Tissue Proteins and Amino Acids as Biomarkers of the Decreased Carbohydrate Oxidation and Increased Amino Acid Oxidation Induced by Caloric Restriction under a Maintained Protein Intake in Obese Rats" Nutrients 11, no. 5: 1087. https://doi.org/10.3390/nu11051087
APA StyleHuneau, J.-F., Mantha, O. L., Hermier, D., Mathé, V., Galmiche, G., Mariotti, F., & Fouillet, H. (2019). Natural Isotope Abundances of Carbon and Nitrogen in Tissue Proteins and Amino Acids as Biomarkers of the Decreased Carbohydrate Oxidation and Increased Amino Acid Oxidation Induced by Caloric Restriction under a Maintained Protein Intake in Obese Rats. Nutrients, 11(5), 1087. https://doi.org/10.3390/nu11051087