1. Introduction
Obesity is a global epidemic, with the number of obese adults currently exceeding 1.9 billion worldwide, with recent estimates suggesting that, for example, by 2030, 42% of Americans will be obese [
1]. Not only is obesity associated with serious cardiovascular and metabolic risk factors and enormous healthcare costs, it has also been linked to an elevated risk for age-related cognitive impairment and the development of Alzheimer’s disease (AD), which is a significantly increasing public health concern [
2,
3,
4,
5].
Accordingly, research on causal and maintenance mechanisms for obesity has become increasingly important, especially as decreasing obesity has the potential to aid in the intervention and prevention of AD and normal age-related cognitive declines. One potential focus in this respect has been the physiological and neural systems involved with appetite and eating behaviors, which are thought to play a key role in the development and maintenance of obesity [
6,
7,
8].
Olfactory processing is a critical component within these systems. For example, olfaction is known to act as a powerful driver of appetite and nutrient consumption, and olfactory input is also known to be critical in determining the palatability of food in both animals and humans. Olfactory processing can also be influenced by appetite and nutrient consumption. For example, behavioral studies have shown that olfactory processing varies as a function of caloric state (e.g., hunger state, and satiety state, i.e., after a preload). In animal models, rodents display improved odor detection and greater food-odor exploration under fasting conditions, however, odor detection and food-odor exploration decline under the condition of satiety [
9,
10,
11,
12]. Research in humans has supported this finding, with studies showing that olfactory perception increases during hunger states and decreases during states of satiety [
12,
13,
14].
At the neural level, primary olfactory processing occurs at the level of the olfactory bulb and the piriform cortex, and then proceeds to higher order cognitive processing and reward regions such as the entorhinal cortex, hippocampus, and striatum [
15]. These regions integrate executive, memory-based and reward information in order to determine knowledge of, decisions about, and behavioral responses to odors experienced in the environment, particularly in the context of food.
Importantly, research suggests that many of these regions are negatively affected in obesity. In animals, electrophysiological studies have demonstrated that rodents fed a high fat diet show a marked loss of olfactory sensory neurons, reduced projections from olfactory sensory neurons to the glomeruli in the olfactory bulb, and poorer olfactory discrimination and odor learning [
16]. In humans, differences in brain function associated with obesity have been associated with poorer performance on olfactory tasks. For example, research using event-related potentials revealed a positive linear relationship between measures of adiposity (including body mass index (BMI) and waist circumference) and latency of the brain response to odors in older adults performing an odor identification task, suggesting brain response during odor identification was significantly slower with increasing adiposity [
17]. Odor identification involves not only the detection of odor, but also the memory for odor and odor names, suggesting the importance of investigating the processing of odor in both sensory and memory areas.
Further research on how differences in olfactory processing at the neural level are related to processing of food-related stimuli within the context of different caloric states (e.g., hunger and satiety) may help elucidate the role of olfactory processing as a potential maintenance mechanism for obesity.
Thus, the main aim of the current study was to gain insight into the relationships between BMI and central olfactory response using event-related functional magnetic resonance imaging (fMRI) to examine brain activation during the hedonic evaluation of a pure odor stimulus in two opposing caloric states: Hunger and satiety. Focusing on activation during a hedonic evaluation task allowed for further investigation of the relationships between BMI and activation in olfactory and higher order memory and reward processing regions, and whether these differ as a function of caloric state.
We hypothesized that activation would vary as a function of BMI such that greater BMI would be associated with differences in activation within olfactory and higher order processing regions important for reward, executive functioning, and memory. Specifically, based on the literature cited above, we hypothesized that activation in reward areas would decrease with increasing BMI, whereas activation in primary olfactory and memory regions would increase with increasing BMI. Further, it was hypothesized that the relationship between activation and BMI would vary as a function of caloric state, driven by greater activation in the hunger state.
6. Discussion
The present study investigated the relationship between BMI and brain activation during the pleasantness rating of a pure odor stimulus within two caloric states: Hunger and satiety. We hypothesized that fMRI activation in the olfactory and higher-order processing regions would vary as a function of caloric condition and BMI.
We found a dissociation between the relationship between BMI and activation in reward areas and in olfactory and odor memory areas, such that activation in reward areas decreased as BMI increased, whereas activation in primary olfactory and memory regions increased as BMI increased. Lower activation in the reward and frontal regions, in association with higher BMI, supports previous findings showing a blunted reward response in obesity that has been linked to changes in decision making, response inhibition, and reward-based behaviors, which may serve as causal and maintenance factors in obesity. In contrast, increasing BMI associated with increased activation in the primary olfactory and memory areas was observed during hunger.
In order to investigate fMRI activation in response to odor under conditions that mimic natural flavor perception and ingestion, the present study used a methodology described by Cerf-Ducastel and Murphy [
19], in which the odorant is delivered orally via an aqueous solution. Importantly, significant activation was seen in the piriform (primary olfactory cortex), an area not reported to be active in previous studies from our group using gustatory stimuli in this paradigm [
36,
37,
38,
39,
40,
41]. These results support the utility of the delivery of odor stimuli via aqueous solutions into the oral cavity, suggesting that brain activation in response to an odor stimulus acting through the retronasal pathway is similar to the brain activation previously shown to occur in response to gustatory stimuli.
8. BMI and Reward Activation
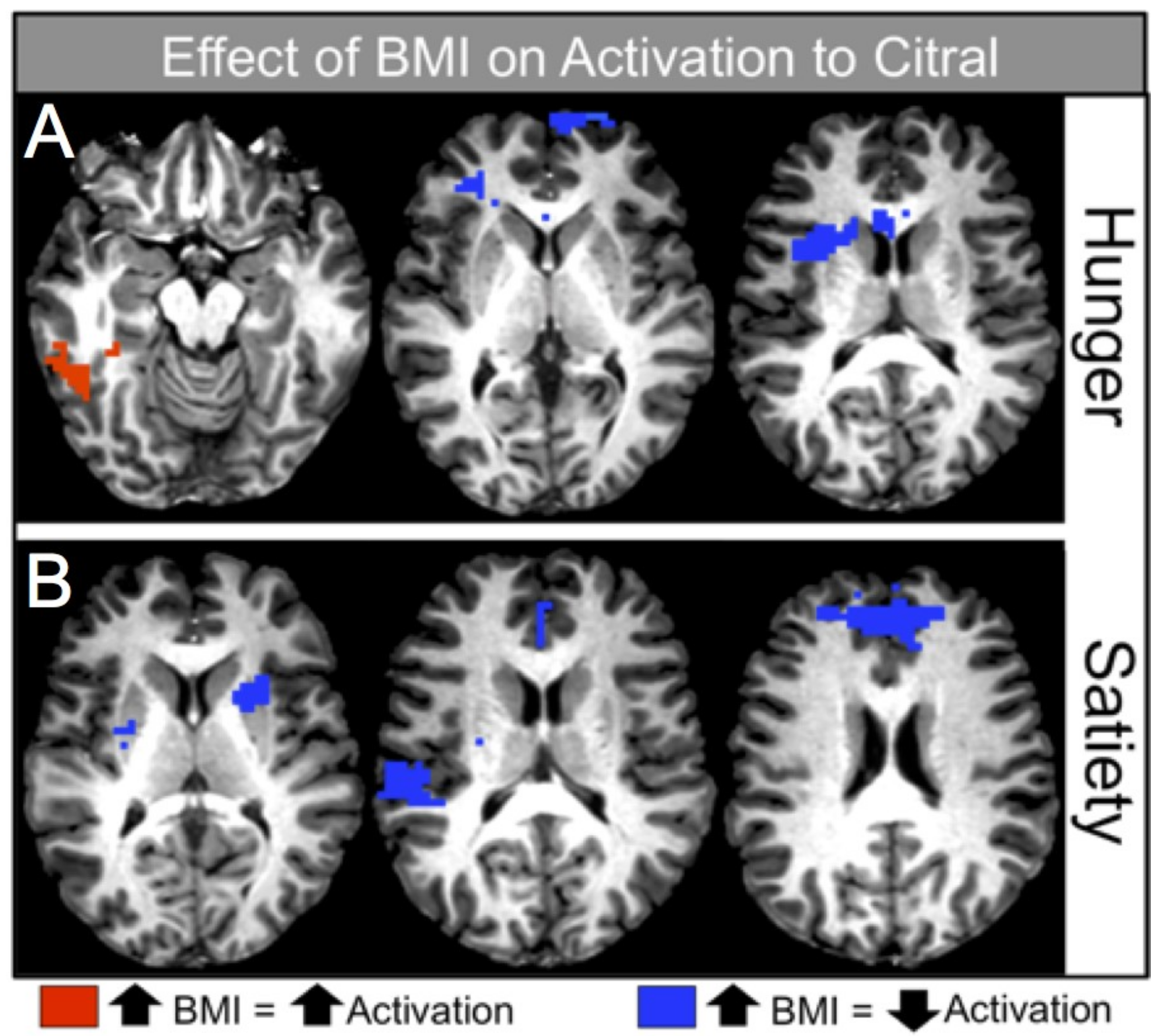
Within both caloric conditions, decreased activation was seen with increasing BMI in brain regions associated with reward processing. For example, increased BMI was associated with decreased activation of the right caudate nucleus in the hunger condition (
Table 6,
Figure 3A, right), and decreased activation of the bilateral caudate and putamen in the satiety condition (
Table 7,
Figure 3B, left).
Negative associations between obesity and reward response to nutritive stimuli have also been reported in overweight adolescent females. Studies have shown increased activation in the gustatory and reward circuitry in response to the anticipation of a chocolate milkshake and decreased gustatory and reward circuitry activation in response to the intake of a chocolate milkshake [
46,
47]. The present findings suggest that decreased reward activation occurs not only in response to complex food stimuli, but also to a simple food odor stimulus.
The relationship between BMI and the caudate nucleus was found in both hunger and satiety conditions. Recent research in humans has linked increased BMI with a decreased caudate response to sucrose, a pure taste stimulus [
43], and to a milkshake, which stimulates both olfaction and taste [
46,
48]. The present results support a decreased caudate response associated with a pure olfactory stimulus. Importantly, the caudate and its connections to other brain regions within the basal ganglia, limbic structures, and the prefrontal cortex are highly associated with reward-related dopaminergic activity. While dopamine levels were not measured directly in the present study, the BOLD response measured during fMRI is directly coupled to dopamine activity [
49,
50].
Thus, these findings lend support to theories suggesting that increasing BMI may be associated with changes in dopaminergic function. In particular, obesity has been associated with decreased striatal D
2 receptor availability, and this abnormality in dopamine function has been linked to reward deficiency syndrome [
51,
52,
53]. Reward deficiency syndrome refers to the predisposition to seek to engage in additional rewarding behaviors to compensate for inefficient dopamine functioning [
52]. In the case of overweight and obese individuals, the rewarding behavior of choice may be over-eating, or eating foods high in sugar or fat content.
9. BMI and Frontal Activation
The present results also indicated a negative relationship between BMI and regions critical for decision-making about rewarding stimuli, such as the frontopolar prefrontal cortex (BA 10;
Table 6 and
Table 7). BA 10 is hypothesized to play a role in binary choices about, and the valuation of, olfactory and reward stimuli, as well as with the ease of and confidence in decision making during olfactory-based choices [
54,
55]. Activation in BA 10 has also been shown to correlate with goal-oriented decisions about food cues and food stimuli [
56,
57].
We might speculate that decreased response in BA 10 would be associated with poor food choice in obesity, especially when coupled with decreases in reward that increase risk-taking and reinforcement-seeking behavior. Notably, dysfunction in the ventro-lateral regions of the prefrontal cortex, such as the inferior frontal gyrus, has also been associated with impulsivity and behavioral factors such as increased risk taking in decision making [
58,
59]. Studies of obesity have linked impulsivity and risky decision making to weight gain and dysfunctional eating behaviors, such as eating in the absence of hunger, binge eating, and emotionally-motivated eating [
47,
60]. Thus, the present findings suggesting decreased activation in both reward and prefrontal brain regions with increasing BMI highlight a potential neuro-behavioral mechanism for obesity.
10. Increased Activation in Olfactory Sensory and Memory Regions during Hunger
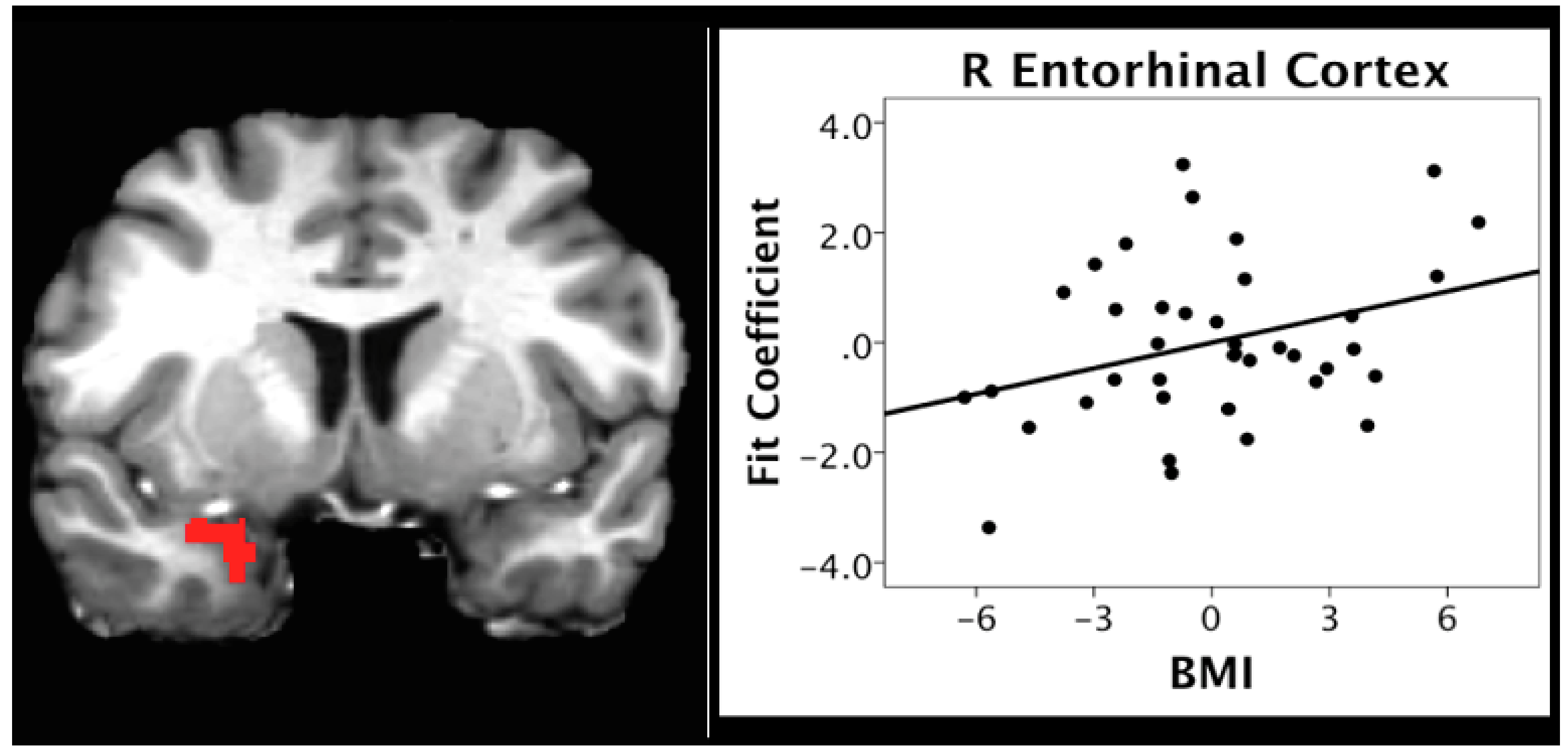
In contrast to the negative relationship between BMI and activation within the reward and frontal regions across caloric conditions, the present study found a positive relationship between BMI and activation within regions important for olfactory and higher-order processing, such as odor memory, during the hunger condition (
Table 6). ROI analyses supported the whole brain analyses, indicating a positive relationship between BMI and activation in regions critical to memory and olfactory processing, specifically the entorhinal and piriform cortices (see
Figure 4 and
Figure 5). Similar findings have been reported by studies in animals showing the hyper-responsivity of sensory processing brain systems in obesity, which has been associated with behaviors such as over-eating [
12].
Hyperactivity associated with increasing obesity may have important implications, as it is theorized that long-term neuronal hyperactivity within a brain region may result in excitotoxic effects that have been implicated as a potential mechanism for the cognitive declines reported in obesity and obesity-related risk factors such as insulin resistance and high-fat diets [
4,
5,
61,
62]. Obesity-related excitotoxicity may also serve as a mechanism for age-related cognitive declines [
63] and neurodegenerative conditions such as AD [
64,
65,
66,
67,
68].
For example, animal models have found that hyperactivity in the hippocampus and the lateral entorhinal cortex has been associated with the production and secretion of amyloid β (Aβ) and Aβ precursor protein (AβPP) metabolites [
66,
69], important in the development of AD. In humans, hyperactivation in the hippocampus, which maintains direct connections to the entorhinal cortex, has been previously reported in cases of very early and moderate mild cognitive impairment (MCI) [
70,
71]. Although the role and the trajectory of hyperactivity in AD is uncertain [
66,
72], it has been reported to be associated with atrophy and clinical progression in amyloid positive patients with MCI, a percentage of whom will go on to develop AD [
73].
The present findings of an association between increasing BMI or waist circumference and hyperactivity in regions critical for olfactory processing are particularly noteworthy, given that neuropathology in the olfactory bulbs and entorhinal cortex manifests very early in AD [
74,
75,
76,
77] and olfactory function has been shown to decline in the preclinical stages of the disease [
67,
68,
78,
79].
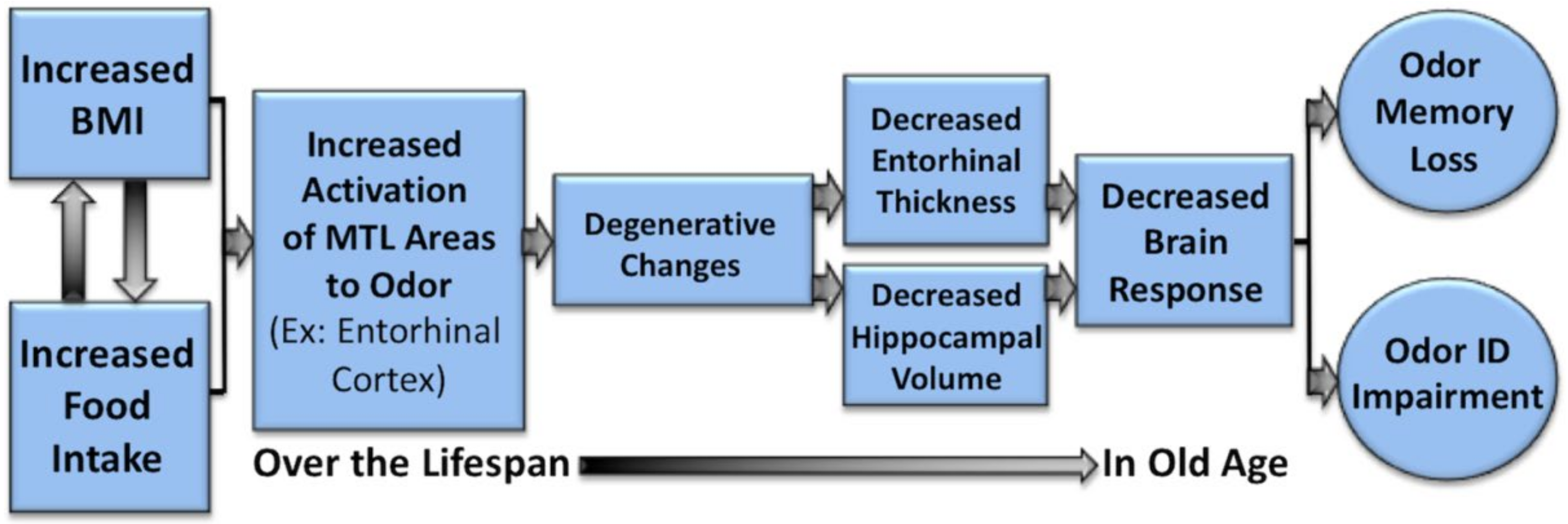
Thus, these data would support a speculative hypothesis that the hyperactivation of olfactory and odor memory processing regions, including the piriform and entorhinal cortices, may be one underlying mechanism for the effects of high BMI on olfactory processing, and on the degeneration of the entorhinal cortex over time in individuals with a high BMI [
67]. If this is the case, it would represent one potential mechanism whereby the BMI may contribute to deterioration of areas critical to olfaction and memory function with aging and neurodegenerative disorders, particularly AD (see
Figure 4,
Figure 5 and
Figure 6). Future research is warranted to systematically examine this hypothesis in longitudinal cohorts.
In summary, we present novel findings of a dissociation between the relationship between BMI and functional brain activation in reward regions, and activation in primary olfactory and memory regions during hedonic evaluation of an odor stimulus: Activation in reward areas decreased as BMI increased in both the hunger and satiety conditions, whereas activation in primary olfactory and higher order processing and memory regions increased as BMI increased during the hunger condition. These findings expand upon previous reports of the effects of overweight and obesity-related variables like adiposity and BMI on brain activation in reward areas in response to gustatory stimuli [
43] and visual food-related stimuli [
42], suggesting that these patterns of activation may be associated with food-related stimuli in general, and are not specific to a sensory modality. A blunted reward and frontal response during the hedonic evaluation of a food-related odor has important potential implications for decision making, response inhibition, and reward-based behaviors such as binge-eating and eating-when-full, which may play key roles as causal and maintenance factors for obesity. Further research investigating how the functional response to olfactory stimuli differs in overweight and normal weight individuals, as well as the mechanisms underlying these changes, is warranted.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}