Tartary Buckwheat Extract Attenuated the Obesity-Induced Inflammation and Increased Muscle PGC-1a/SIRT1 Expression in High Fat Diet-Induced Obese Rats

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Tartary Buckwheat Extracts
2.2. Total Phenolic, Total Flavonoid, and Rutin Determination
2.3. Animals and Experimental Design
2.4. Serum Chemical Measurement
2.5. Histological Analysis of Adipose Tissue
2.6. Immunohistochemistry of Adipose Tissue
2.7. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.8. Serum TNF-α Measurement
2.9. Nitric Oxide (NO) Production Measurement
2.10. Glycerol-3-Phosphate Dehydrogenase (GPDH) Activity Analysis
2.11. Sirtulin (SIRT) and AMP-Activated Protein Kinase (AMPK) Activitivess
2.12. Statistical Analysis
3. Results
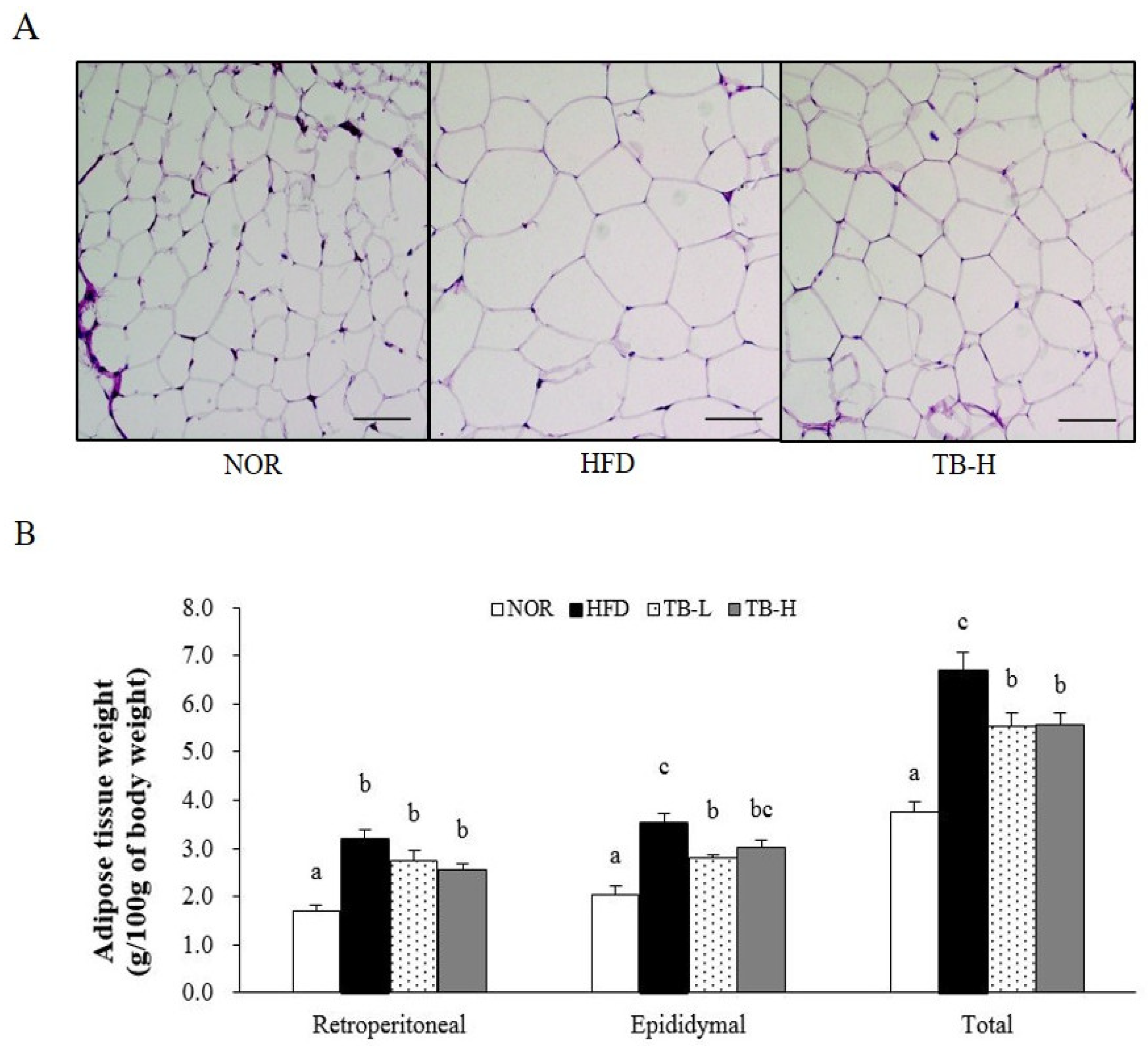
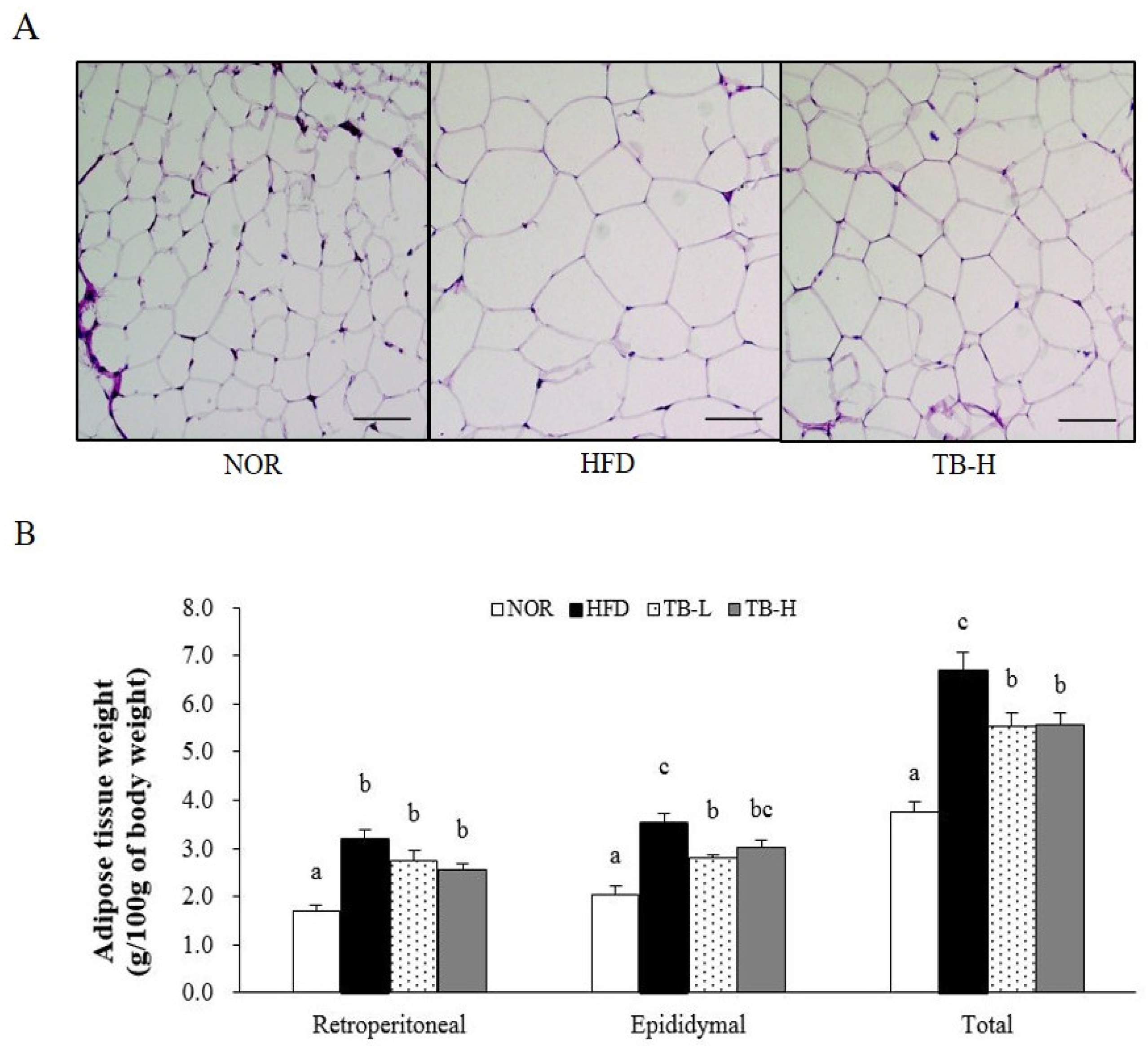
3.1. Effect of Tartary Buckwheat Extract on Body Fat Mass
3.2. Effect of Tartary Buckwheat Extract on Body Fat Mass
3.3. Effect of Tartary Buckwheat Extract on Serum Lipid Profiles
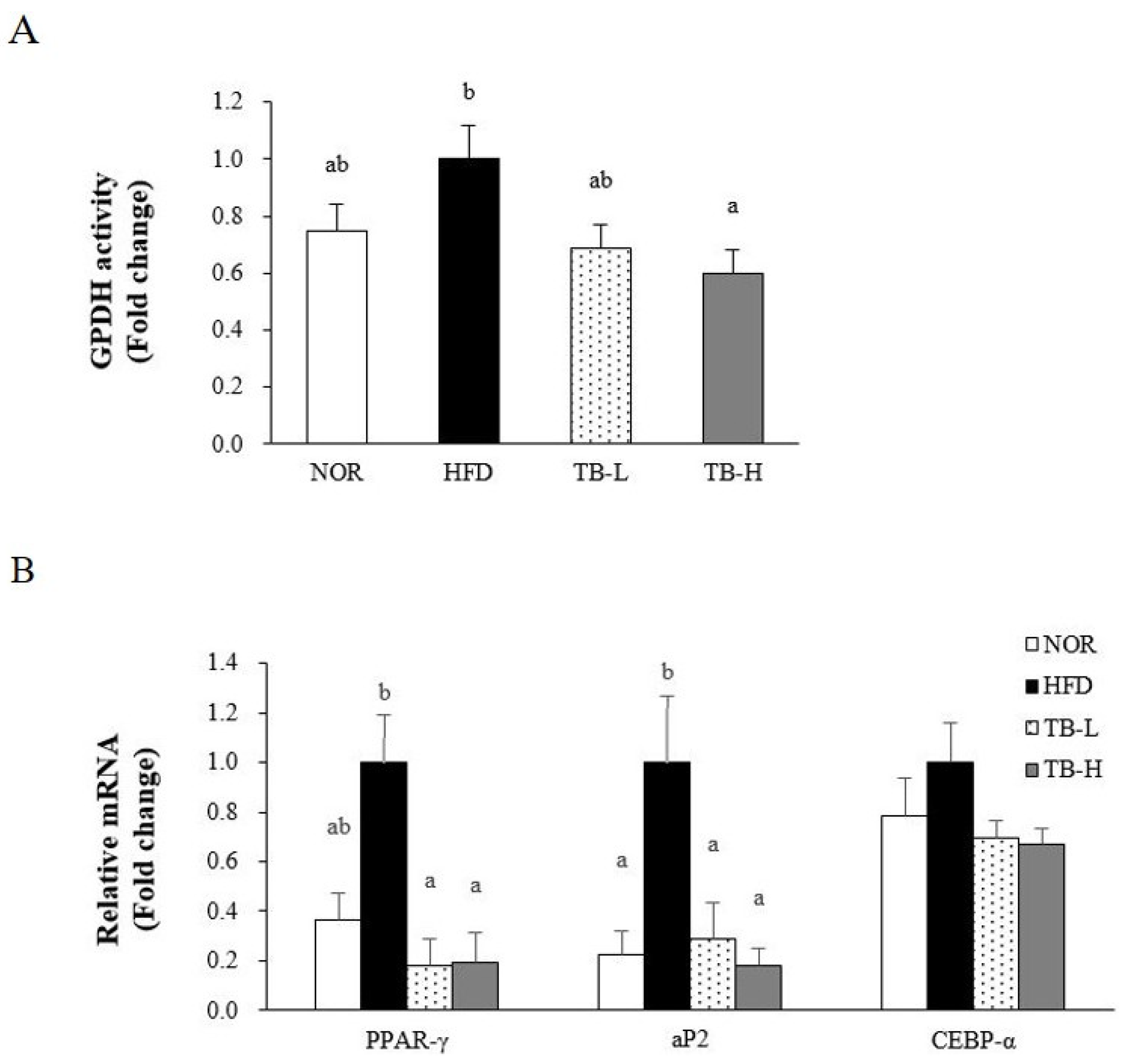
3.4. Effect of Tartary Buckwheat Extract on Lipid Accumulation
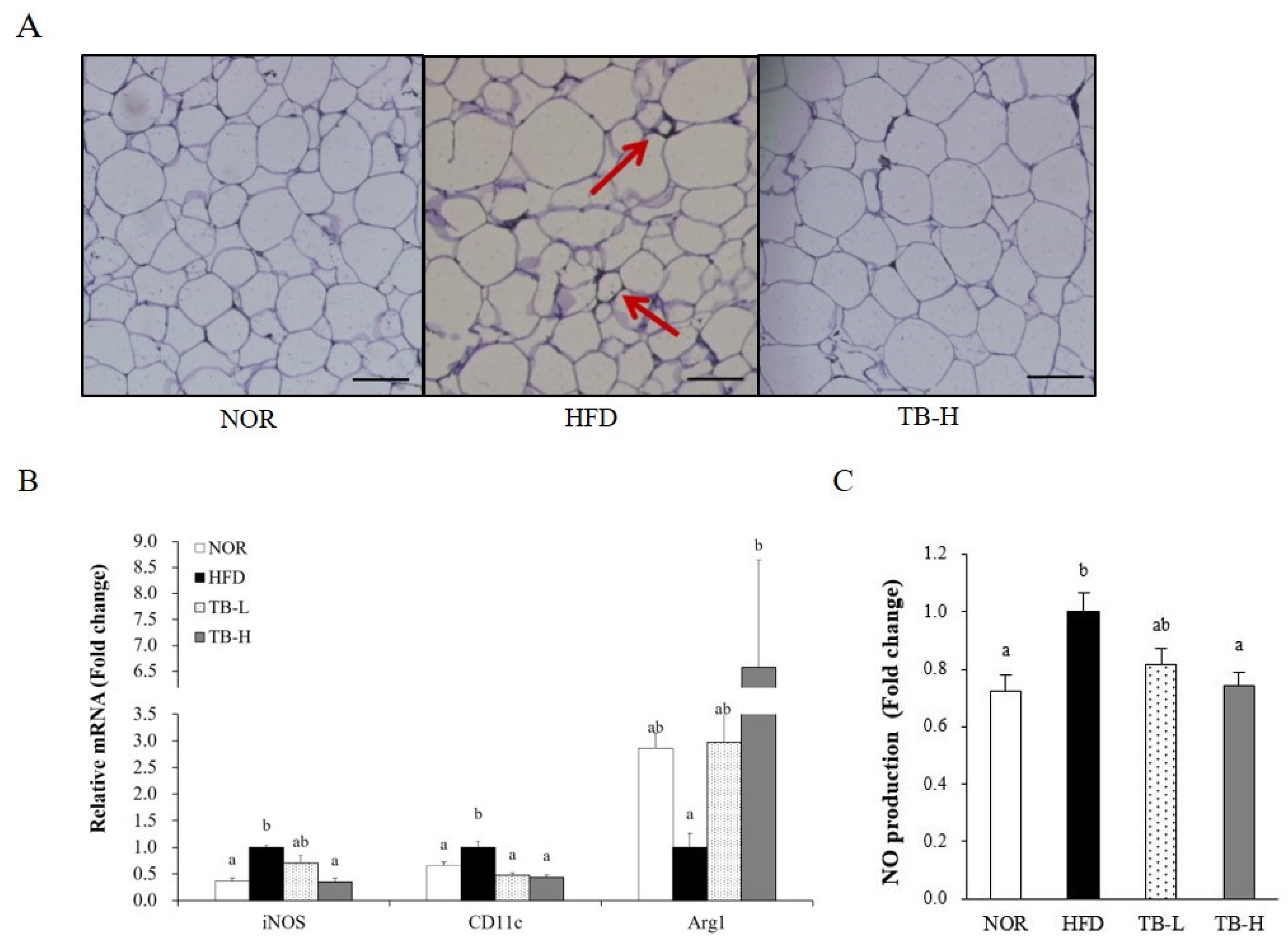
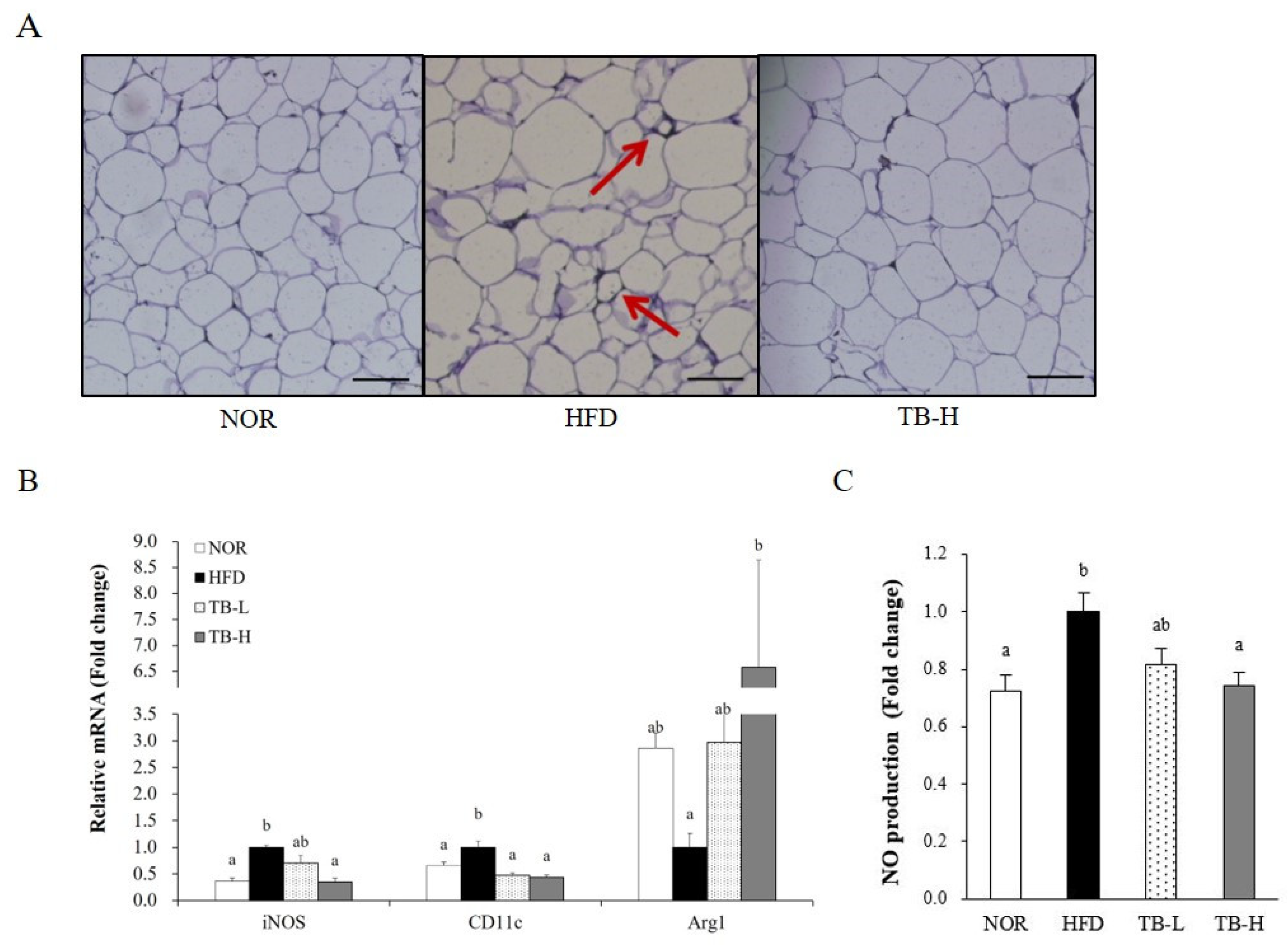
3.5. Effect of Tartary Buckwheat Extract on Macrophage Polarization in Adipose Tissue
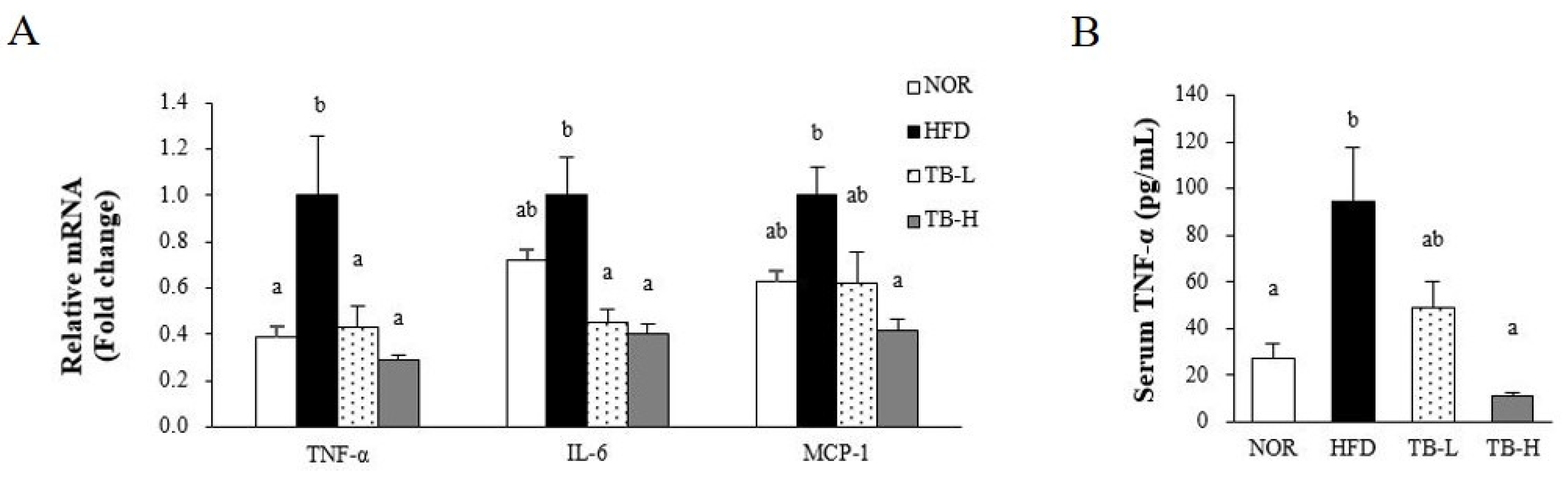
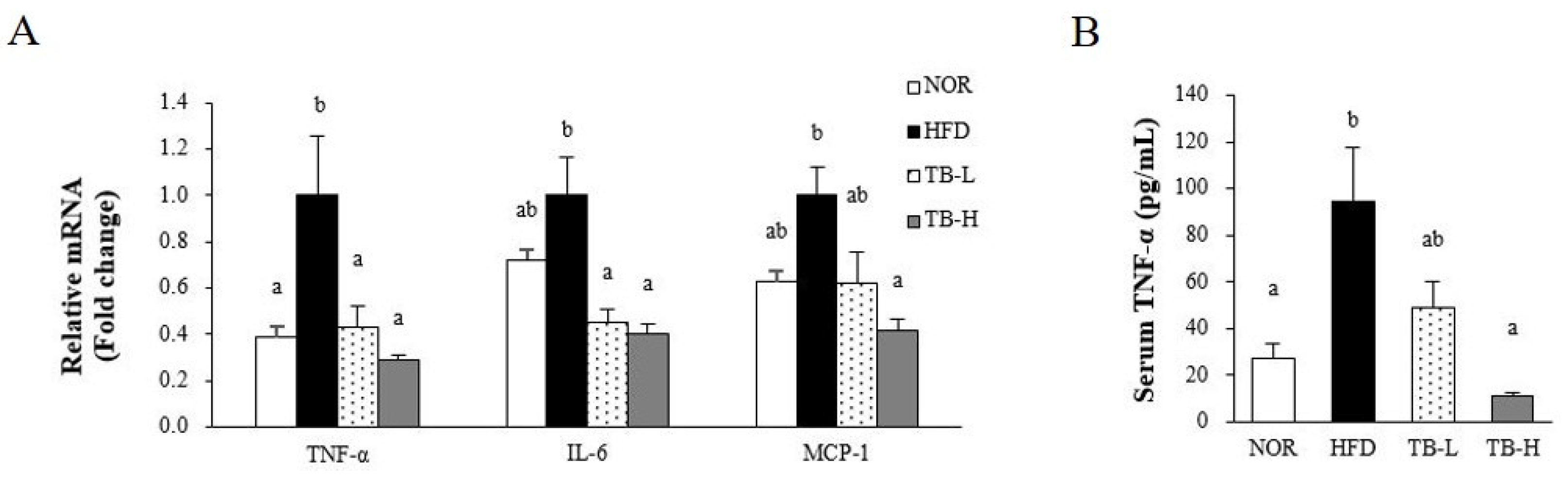
3.6. Effect of Tartary Buckwheat Extract on Inflammatory Response
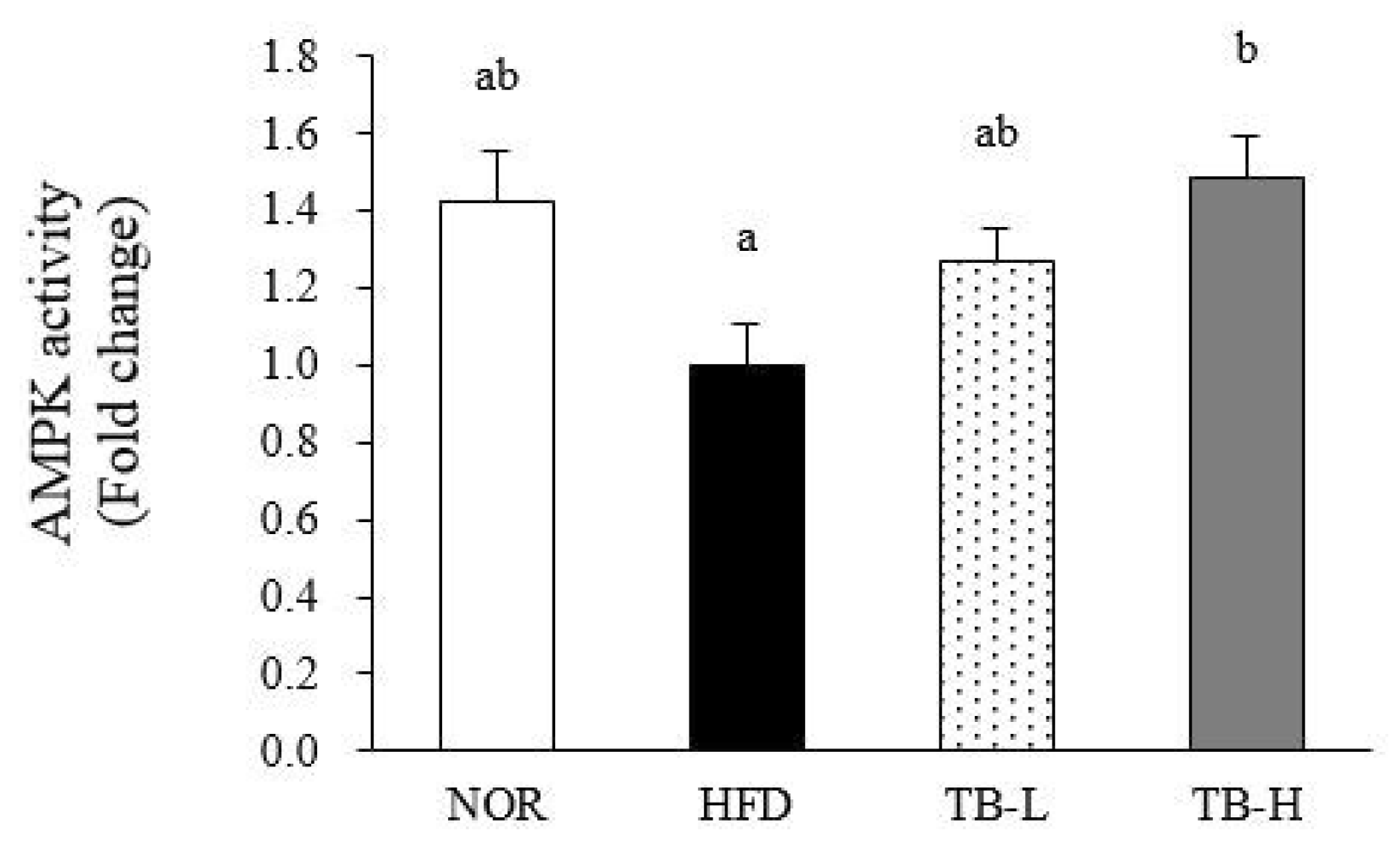
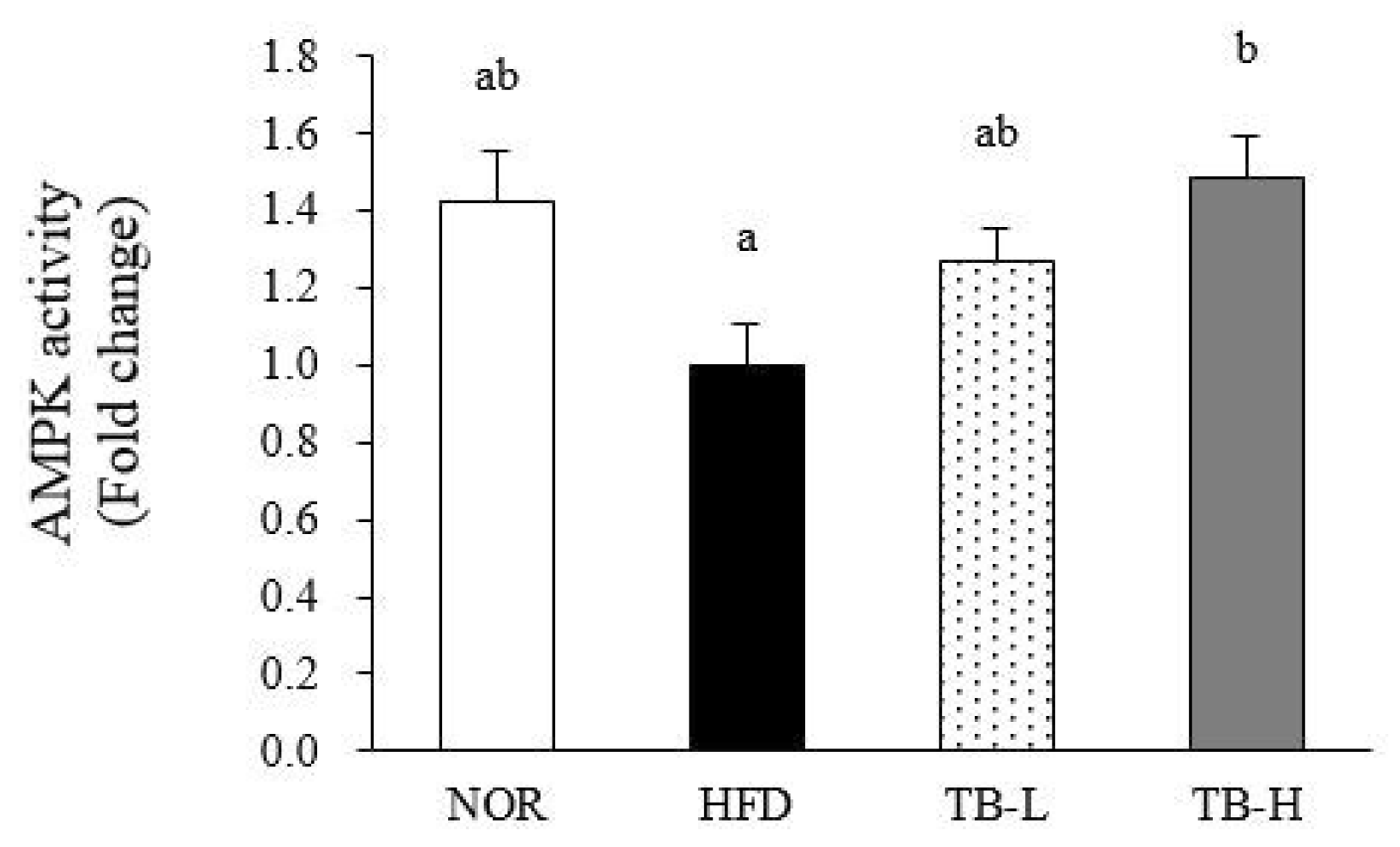
3.7. Effect of Tartary Buckwheat Extract on AMPK Activity in Adipose Tissue
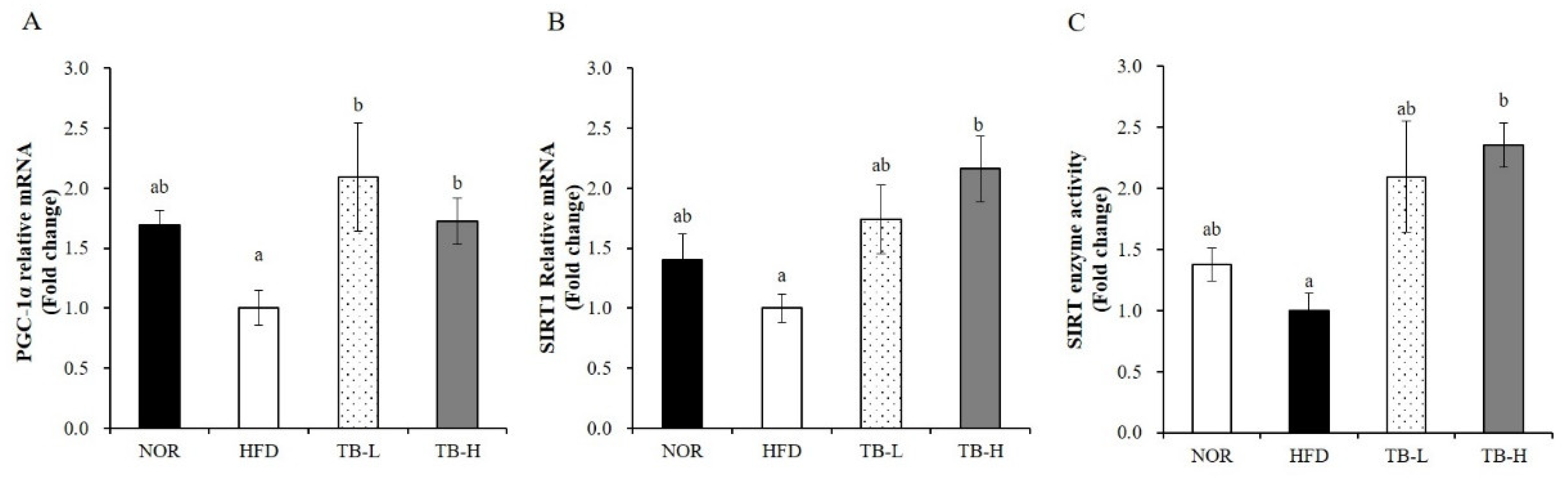
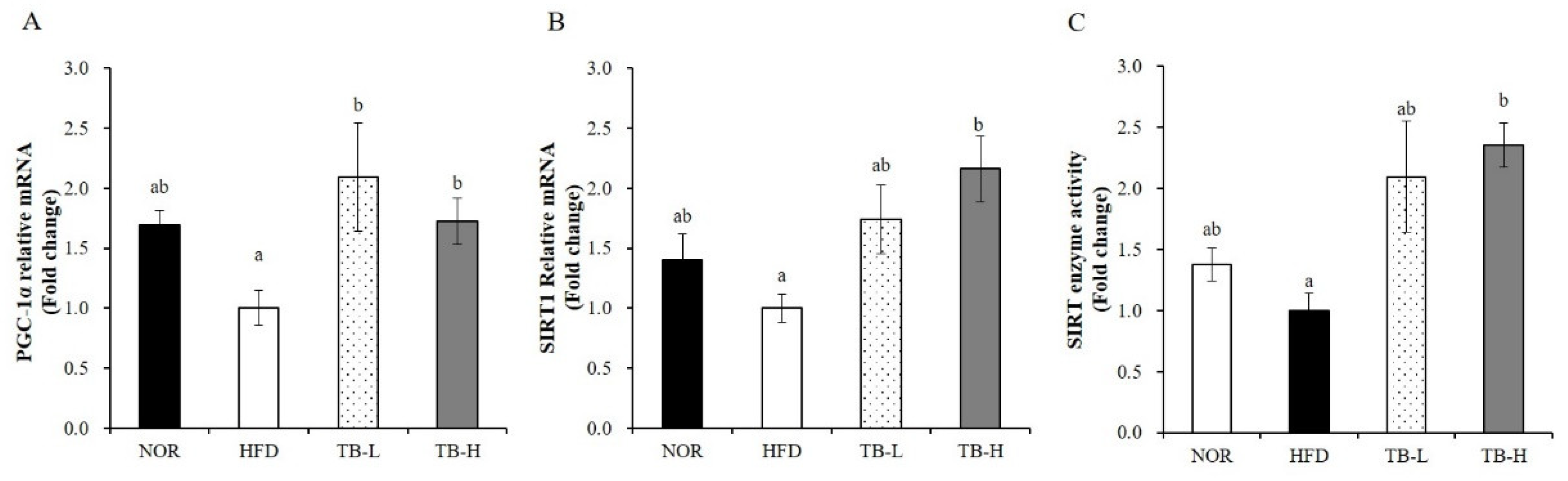
3.8. Effect of Tartary Buckwheat Extract on Mitochondrial Gene Expression and SIRT Activity in Skeletal Muscle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Puhl, R.M.; Heuer, C.A. The stigma of obesity: A review and update. Obesity 2009, 17, 941–964. [Google Scholar] [CrossRef]
- de Ferranti, S.; Mozaffarian, D. The perfect storm: Obesity, adipocyte dysfunction, and metabolic consequences. Clin. Chem. 2008, 54, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 2010, 289645. [Google Scholar] [CrossRef] [PubMed]
- Lafontan, M. Adipose tissue and adipocyte dysregulation. Diabetes Metab. 2014, 40, 16–28. [Google Scholar] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef]
- Bonnard, C.; Durand, A.; Peyrol, S.; Chanseaume, E.; Chauvin, M.A.; Morio, B.; Vidal, H.; Rieusset, J. Mitochondrial dysfunction results from oxidative stress in the skeletal muscle of diet-induced insulin-resistant mice. J. Clin. Investig. 2008, 118, 789–800. [Google Scholar] [CrossRef]
- Koves, T.R.; Ussher, J.R.; Noland, R.C.; Slentz, D.; Mosedale, M.; Ilkayeva, O.; Bain, J.; Stevens, R.; Dyck, J.R.; Newgard, C.B.; et al. Mitochondrial overload and incomplete fatty acid oxidation contribute to skeletal muscle insulin resistance. Cell Metab. 2008, 7, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Hirabara, S.M.; Curi, R.; Maechler, P. Saturated fatty acid-induced insulin resistance is associated with mitochondrial dysfunction in skeletal muscle cells. J. Cell Physiol. 2010, 222, 187–194. [Google Scholar] [CrossRef]
- Kelley, D.E.; He, J.; Menshikova, E.V.; Ritov, V.B. Dysfunction of mitochondria in human skeletal muscle in type 2 diabetes. Diabetes 2002, 51, 2944–2950. [Google Scholar] [CrossRef]
- Kim, J.Y.; Hickner, R.C.; Cortright, R.L.; Dohm, G.L.; Houmard, J.A. Lipid oxidation is reduced in obese human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E1039–E1044. [Google Scholar] [CrossRef] [PubMed]
- Befroy, D.E.; Petersen, K.F.; Dufour, S.; Mason, G.F.; de Graaf, R.A.; Rothman, D.L.; Shulman, G.I. Impaired mitochondrial substrate oxidation in muscle of insulin-resistant offspring of type 2 diabetic patients. Diabetes 2007, 56, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Murai, M.; Ohnishi, O. Diffusion routes of buckwheat cultivation in Asia revealed by RAPD markers. Curr. Adv. Buckwheat Res 1995, 163–174. [Google Scholar]
- Jiang, P.; Burczynski, F.; Campbell, C.; Pierce, G.; Austria, J.; Briggs, C. Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum, F. tataricum, and F. homotropicum and their protective effects against lipid peroxidation. Food Res. Int. 2007, 40, 356–364. [Google Scholar] [CrossRef]
- Karki, R.; Park, C.H.; Kim, D.W. Extract of buckwheat sprouts scavenges oxidation and inhibits pro-inflammatory mediators in lipopolysaccharide-stimulated macrophages (RAW264.7). J. Integr. Med. 2013, 11, 246–252. [Google Scholar] [CrossRef]
- Lee, M.S.; Shin, Y.; Jung, S.; Kim, S.Y.; Jo, Y.H.; Kim, C.T.; Yun, M.K.; Lee, S.J.; Sohn, J.; Yu, H.J.; et al. The inhibitory effect of tartary buckwheat extracts on adipogenesis and inflammatory response. Molecules 2017, 22, 1160. [Google Scholar] [CrossRef]
- Hainal, A.-R.; Ignat, I.; Volf, I.; Popa, V.I. Transformation of polyphenols from biomass by some yeast species. Cell. Chem. Technol. 2011, 45, 211. [Google Scholar]
- Zdravković, J.M.; Aćamović-Đoković, G.S.; Mladenović, J.D.; Pavlović, R.M.; Zdravković, M.S. Antioxidant capacity and contents of phenols, ascorbic acid, β-carotene and lycopene in lettuce. HEM. IND. 2014, 68, 193–198. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Divella, R.; De Luca, R.; Abbate, I.; Naglieri, E.; Daniele, A. Obesity and cancer: The role of adipose tissue and adipo-cytokines-induced chronic inflammation. J. Cancer 2016, 7, 2346–2359. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.; Bruce, C.R.; Beale, S.M.; Hoehn, K.L.; So, T.; Rolph, M.S.; Cooney, G.J. Excess lipid availability increases mitochondrial fatty acid oxidative capacity in muscle: Evidence against a role for reduced fatty acid oxidation in lipid-induced insulin resistance in rodents. Diabetes 2007, 56, 2085–2092. [Google Scholar] [CrossRef]
- Wang, M.; Liu, J.R.; Gao, J.M.; Parry, J.W.; Wei, Y.M. Antioxidant activity of tartary buckwheat bran extract and its effect on the lipid profile of hyperlipidemic rats. J. Agric. Food Chem. 2009, 57, 5106–5112. [Google Scholar] [CrossRef]
- Jin, H.M.; Wei, P. Anti-fatigue properties of tartary buckwheat extracts in mice. Int. J. Mol. Sci. 2011, 12, 4770–4780. [Google Scholar] [CrossRef]
- Panchal, S.K.; Poudyal, H.; Arumugam, T.V.; Brown, L. Rutin attenuates metabolic changes, nonalcoholic steatohepatitis, and cardiovascular remodeling in high-carbohydrate, high-fat diet-fed rats. J. Nutr. 2011, 141, 1062–1069. [Google Scholar] [CrossRef]
- Al-Rejaie, S.S.; Aleisa, A.M.; Sayed-Ahmed, M.M.; Al-Shabanah, O.A.; Abuohashish, H.M.; Ahmed, M.M.; Al-Hosaini, K.A.; Hafez, M.M. Protective effect of rutin on the antioxidant genes expression in hypercholestrolemic male Westar rat. BMC Complement. Altern. Med. 2013, 13, 136. [Google Scholar] [CrossRef]
- Seo, S.; Lee, M.S.; Chang, E.; Shin, Y.; Oh, S.; Kim, I.H.; Kim, Y. Rutin increases muscle mitochondrial biogenesis with ampk activation in high-fat diet-induced obese rats. Nutrients 2015, 7, 8152–8169. [Google Scholar] [CrossRef]
- Samuelsson, L.; Stromberg, K.; Vikman, K.; Bjursell, G.; Enerback, S. The CCAAT/enhancer binding protein and its role in adipocyte differentiation: Evidence for direct involvement in terminal adipocyte development. EMBO J. 1991, 10, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Cowherd, R.M.; Lyle, R.E.; McGehee, R.E., Jr. Molecular regulation of adipocyte differentiation. Semin. Cell Dev. Biol. 1999, 10, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Jung, S.; Oh, S.; Shin, Y.; Kim, C.T.; Kim, I.H.; Kim, Y. Effect of high hydrostatic pressure extract of fresh ginseng on adipogenesis in 3T3-L1 adipocytes. J. Sci. Food Agric. 2015, 95, 2409–2415. [Google Scholar] [CrossRef]
- Hosogai, N.; Fukuhara, A.; Oshima, K.; Miyata, Y.; Tanaka, S.; Segawa, K.; Furukawa, S.; Tochino, Y.; Komuro, R.; Matsuda, M.; et al. Adipose tissue hypoxia in obesity and its impact on adipocytokine dysregulation. Diabetes 2007, 56, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Gao, Z.; Yin, J.; He, Q. Hypoxia is a potential risk factor for chronic inflammation and adiponectin reduction in adipose tissue of ob/ob and dietary obese mice. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1118–E1128. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Cook, H.T.; Cattell, V. Role of nitric oxide in immune-mediated diseases. Clin. Sci. (London) 1996, 91, 375–384. [Google Scholar] [CrossRef]
- Liu, S.F.; Adcock, I.M.; Old, R.W.; Barnes, P.J.; Evans, T.W. Differential regulation of the constitutive and inducible nitric oxide synthase mRNA by lipopolysaccharide treatment in vivo in the rat. Crit. Care Med. 1996, 24, 1219–1225. [Google Scholar] [CrossRef]
- Soeters, P.B.; Hallemeesch, M.M.; Bruins, M.J.; van Eijk, H.M.; Deutz, N.E. Quantitative in vivo assessment of arginine utilization and nitric oxide production in endotoxemia. Am. J. Surg. 2002, 183, 480–488. [Google Scholar] [CrossRef]
- Gao, M.; Ma, Y.; Liu, D. Rutin suppresses palmitic acids-triggered inflammation in macrophages and blocks high fat diet-induced obesity and fatty liver in mice. Pharm. Res. 2013, 30, 2940–2950. [Google Scholar] [CrossRef]
- Christiansen, T.; Richelsen, B.; Bruun, J.M. Monocyte chemoattractant protein-1 is produced in isolated adipocytes, associated with adiposity and reduced after weight loss in morbid obese subjects. Int. J. Obes. (Lond) 2005, 29, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Rossmeisl, M.; Flachs, P.; Brauner, P.; Sponarova, J.; Matejkova, O.; Prazak, T.; Ruzickova, J.; Bardova, K.; Kuda, O.; Kopecky, J. Role of energy charge and AMP-activated protein kinase in adipocytes in the control of body fat stores. Int. J. Obes. Relat. Metab. Disord. 2004, 28 (Suppl. 4), S38–S44. [Google Scholar] [CrossRef]
- Gaidhu, M.P.; Bikopoulos, G.; Ceddia, R.B. Chronic AICAR-induced AMP-kinase activation regulates adipocyte lipolysis in a time-dependent and fat depot-specific manner in rats. Am. J. Physiol. Cell Physiol. 2012, 303, C1192–C1197. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Schertzer, J.D. AMPK promotes macrophage fatty acid oxidative metabolism to mitigate inflammation: Implications for diabetes and cardiovascular disease. Immunol. Cell Biol. 2014, 92, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Sag, D.; Carling, D.; Stout, R.D.; Suttles, J. Adenosine 5’-monophosphate-activated protein kinase promotes macrophage polarization to an anti-inflammatory functional phenotype. J. Immunol. 2008, 181, 8633–8641. [Google Scholar] [PubMed]
- Galic, S.; Fullerton, M.D.; Schertzer, J.D.; Sikkema, S.; Marcinko, K.; Walkley, C.R.; Izon, D.; Honeyman, J.; Chen, Z.P.; van Denderen, B.J.; et al. Hematopoietic AMPK β1 reduces mouse adipose tissue macrophage inflammation and insulin resistance in obesity. J. Clin. Investig. 2011, 121, 4903–4915. [Google Scholar] [CrossRef] [PubMed]
- Heilbronn, L.K.; Gan, S.K.; Turner, N.; Campbell, L.V.; Chisholm, D.J. Markers of mitochondrial biogenesis and metabolism are lower in overweight and obese insulin-resistant subjects. J. Clin. Endocrinol. Metab. 2007, 92, 1467–1473. [Google Scholar] [CrossRef]
- Mensink, M.; Hesselink, M.K.; Russell, A.P.; Schaart, G.; Sels, J.P.; Schrauwen, P. Improved skeletal muscle oxidative enzyme activity and restoration of PGC-1 alpha and PPAR beta/delta gene expression upon rosiglitazone treatment in obese patients with type 2 diabetes mellitus. Int. J. Obes. (Lond) 2007, 31, 1302–1310. [Google Scholar] [CrossRef]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef]
- Huang, T.Y.; Zheng, D.; Houmard, J.A.; Brault, J.J.; Hickner, R.C.; Cortright, R.N. Overexpression of PGC-1α increases peroxisomal activity and mitochondrial fatty acid oxidation in human primary myotubes. Am. J. Physiol. Endocrinol. Metab. 22017, 312, E253–E263. [Google Scholar] [CrossRef]
- Gerhart-Hines, Z.; Rodgers, J.T.; Bare, O.; Lerin, C.; Kim, S.H.; Mostoslavsky, R.; Alt, F.W.; Wu, Z.; Puigserver, P. Metabolic control of muscle mitochondrial function and fatty acid oxidation through SIRT1/PGC-1α. EMBO J. 2007, 26, 1913–1923. [Google Scholar] [CrossRef]
- Nemoto, S.; Fergusson, M.M.; Finkel, T. SIRT1 functionally interacts with the metabolic regulator and transcriptional coactivator pgc-1α. J. Biol. Chem. 2005, 280, 16456–16460. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | HFD | TB-L | TB-H |
|---|---|---|---|
| Casein | 238.79 | 238.79 | 238.79 |
| Corn starch | 185.11 | 180.11 | 175.11 |
| Dextrose | 157.60 | 157.60 | 157.60 |
| Sucrose | 59.70 | 59.70 | 59.70 |
| Cellulose | 59.70 | 59.70 | 59.70 |
| Soybean oil | 29.85 | 29.85 | 29.85 |
| Lard | 208.94 | 208.94 | 208.94 |
| tert-Butylhydroquinone (TBHQ) | 0.02 | 0.02 | 0.02 |
| Mineral mix 1 | 41.79 | 41.79 | 41.79 |
| Vitamin mix 2 | 11.94 | 11.94 | 11.94 |
| L-cystine | 3.58 | 3.58 | 3.58 |
| Choline bitartrate | 2.98 | 2.98 | 2.98 |
| TB 3 | - | 5.00 | 10.00 |
| Total | 1000 | 1000 | 1000 |
| Energy density (kcal/g) | 4.8 | 4.8 | 4.8 |
| Carb % (calories) | 35 | 35 | 35 |
| Fat % (calories) | 45 | 45 | 45 |
| Protein % (calories) | 20 | 20 | 20 |
| Contents | Tartary Buckwheat Extract |
|---|---|
| Total phenolics (mg GAE/g dried sample) | 101.11 ± 5.5 |
| Total flavonoid (mg QE/g dried sample) | 95.05 ± 3.6 |
| Rutin (mg/g dried sample) | 106.02 ± 1.3 |
| Variables | NOR | HFD | TB-L | TB-H |
|---|---|---|---|---|
| Initial body weight (g) | 67.24 ± 2.16 a | 66.16 ± 2.23 a | 66.42 ± 2.31 a | 66.90 ± 2.17 a |
| Final body weight (g) | 482.46 ± 16.86 a | 554.49 ± 12.52 b | 556.47± 19.08 b | 555.42 ± 13.26 b |
| Body weight gain (g/13 weeks) | 415.22 ± 17.49 a | 488.33 ± 11.32 b | 490.05 ± 19.87 b | 488.52 ± 12.25 b |
| Food intake (g/day) | 25.59 ± 1.62 a | 21.02 ± 0.54 a | 20.28 ± 0.53 a | 20.17 ± 0.50 a |
| Energy intake (kcal/day) | 79.33 ± 5.01 b | 100.49 ± 2.58 a | 96.95 ± 2.53 a | 96.41 ± 2.40 a |
| Energy efficiency ratio (EER) 1 | 0.058 ± 0.004 a | 0.053 ± 0.000 a | 0.055 ± 0.001 a | 0.056 ± 0.000 a |
| Variables | NOR | HFD | TB-L | TB-H |
|---|---|---|---|---|
| Serum lipids (mmol/L) | ||||
| Triglyceride | 0.58 ± 0.04 ab | 0.73 ± 0.07 b | 0.43 ± 0.03 a | 0.48 ± 0.07 a |
| Total cholesterol | 2.79 ± 0.17 bc | 2.98 ± 0.16 c | 2.24± 0.13 ab | 2.03 ± 0.13 a |
| HDL cholesterol | 1.59 ± 0.08 b | 1.23 ± 0.07 a | 1.63 ± 0.07 b | 1.46 ± 0.02 ab |
| LDL cholesterol | 1.06 ± 0.12 b | 1.50 ± 0.11 b | 0.52 ± 0.10 a | 0.56 ± 0.12 a |
| Liver toxicity (IU/L) | ||||
| AST | 38.96 ± 2.17 | 40.10 ± 3.61 | 37.91 ± 3.38 | 37.99 ± 2.64 |
| ALT | 8.23 ± 0.52 | 8.06 ± 0.64 | 6.86 ± 0.80 | 7.11 ± 0.32 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-Y.; Lee, M.-S.; Chang, E.; Jung, S.; Ko, H.; Lee, E.; Lee, S.; Kim, C.-T.; Kim, I.-H.; Kim, Y. Tartary Buckwheat Extract Attenuated the Obesity-Induced Inflammation and Increased Muscle PGC-1a/SIRT1 Expression in High Fat Diet-Induced Obese Rats. Nutrients 2019, 11, 654. https://doi.org/10.3390/nu11030654
Kim S-Y, Lee M-S, Chang E, Jung S, Ko H, Lee E, Lee S, Kim C-T, Kim I-H, Kim Y. Tartary Buckwheat Extract Attenuated the Obesity-Induced Inflammation and Increased Muscle PGC-1a/SIRT1 Expression in High Fat Diet-Induced Obese Rats. Nutrients. 2019; 11(3):654. https://doi.org/10.3390/nu11030654
Chicago/Turabian StyleKim, Seog-Young, Mak-Soon Lee, Eugene Chang, Sunyoon Jung, Hyunmi Ko, Eunyoung Lee, Soojin Lee, Chong-Tai Kim, In-Hwan Kim, and Yangha Kim. 2019. "Tartary Buckwheat Extract Attenuated the Obesity-Induced Inflammation and Increased Muscle PGC-1a/SIRT1 Expression in High Fat Diet-Induced Obese Rats" Nutrients 11, no. 3: 654. https://doi.org/10.3390/nu11030654
APA StyleKim, S.-Y., Lee, M.-S., Chang, E., Jung, S., Ko, H., Lee, E., Lee, S., Kim, C.-T., Kim, I.-H., & Kim, Y. (2019). Tartary Buckwheat Extract Attenuated the Obesity-Induced Inflammation and Increased Muscle PGC-1a/SIRT1 Expression in High Fat Diet-Induced Obese Rats. Nutrients, 11(3), 654. https://doi.org/10.3390/nu11030654