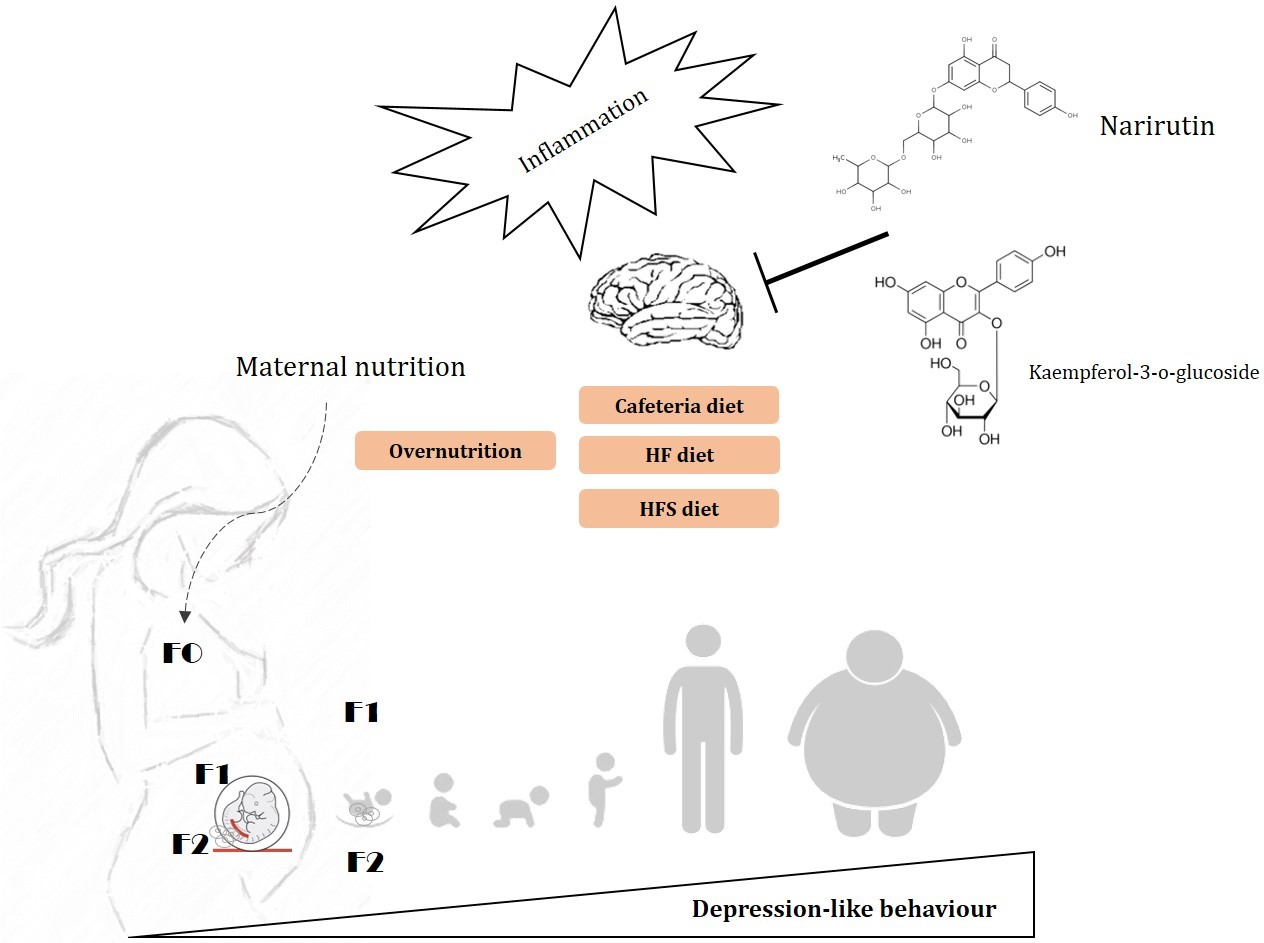
Maternal Flavonoids Intake Reverts Depression-Like Behaviour in Rat Female Offspring

 ,
,  , , and
, , and
Abstract

1. Introduction
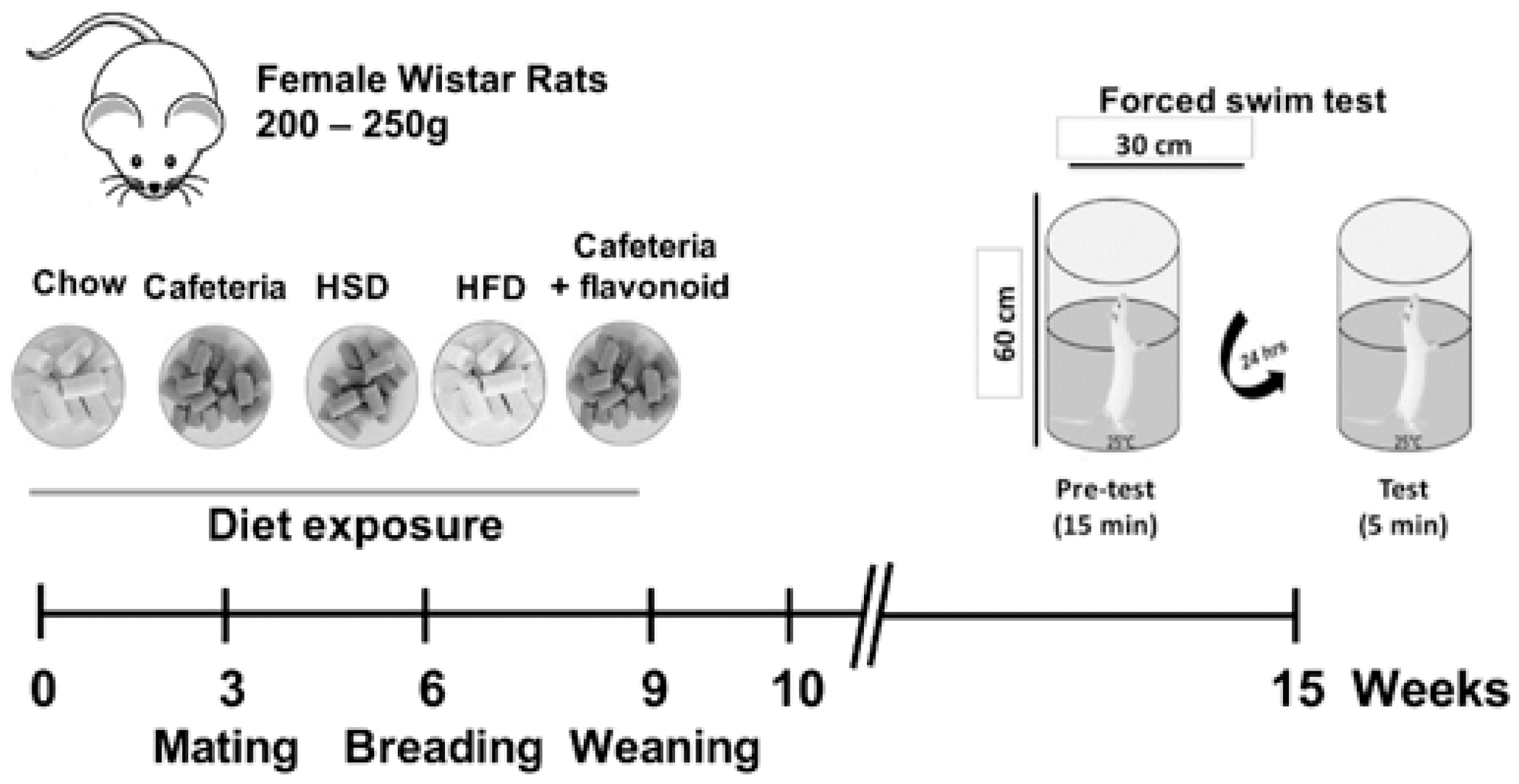
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Animals and Housing
2.3. Diets
2.4. Nutritional Programming by HSD, HFD, Cafeteria, and Cafeteria Diet Supplemented with Kaempferol-3-O-Glucoside and Narirutin
2.5. Forced Swim Test
2.6. Prefrontal Cortex (PFC) and Hippocampus Isolation
2.7. Western Blot
2.8. Statistical Analysis
3. Results
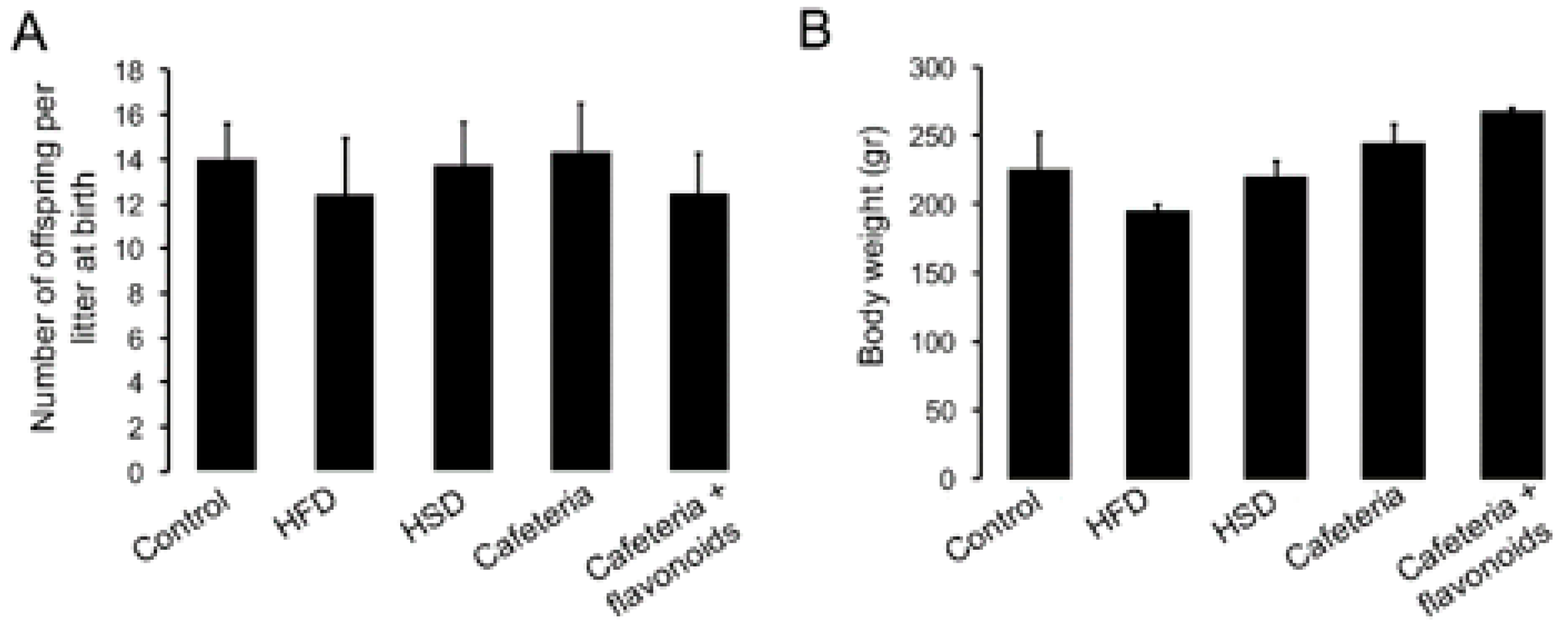
3.1. Nutritional Programming by Diet Exposure Modulates Conception and Survival of Male Offspring
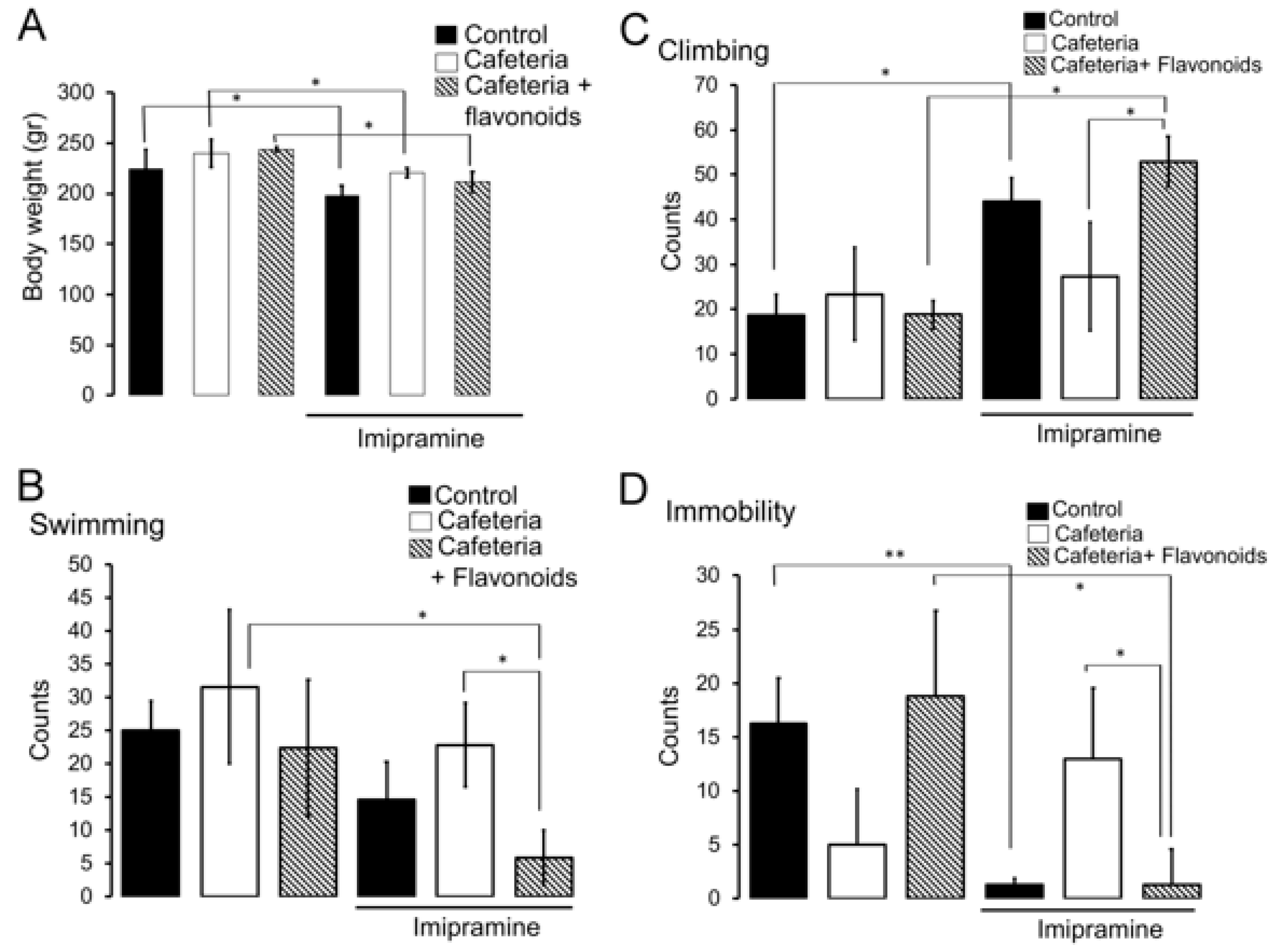
3.2. HSD Exposure during Pregnancy and Lactation Promotes Insensitivity to the Anti-Depressant Imipramine
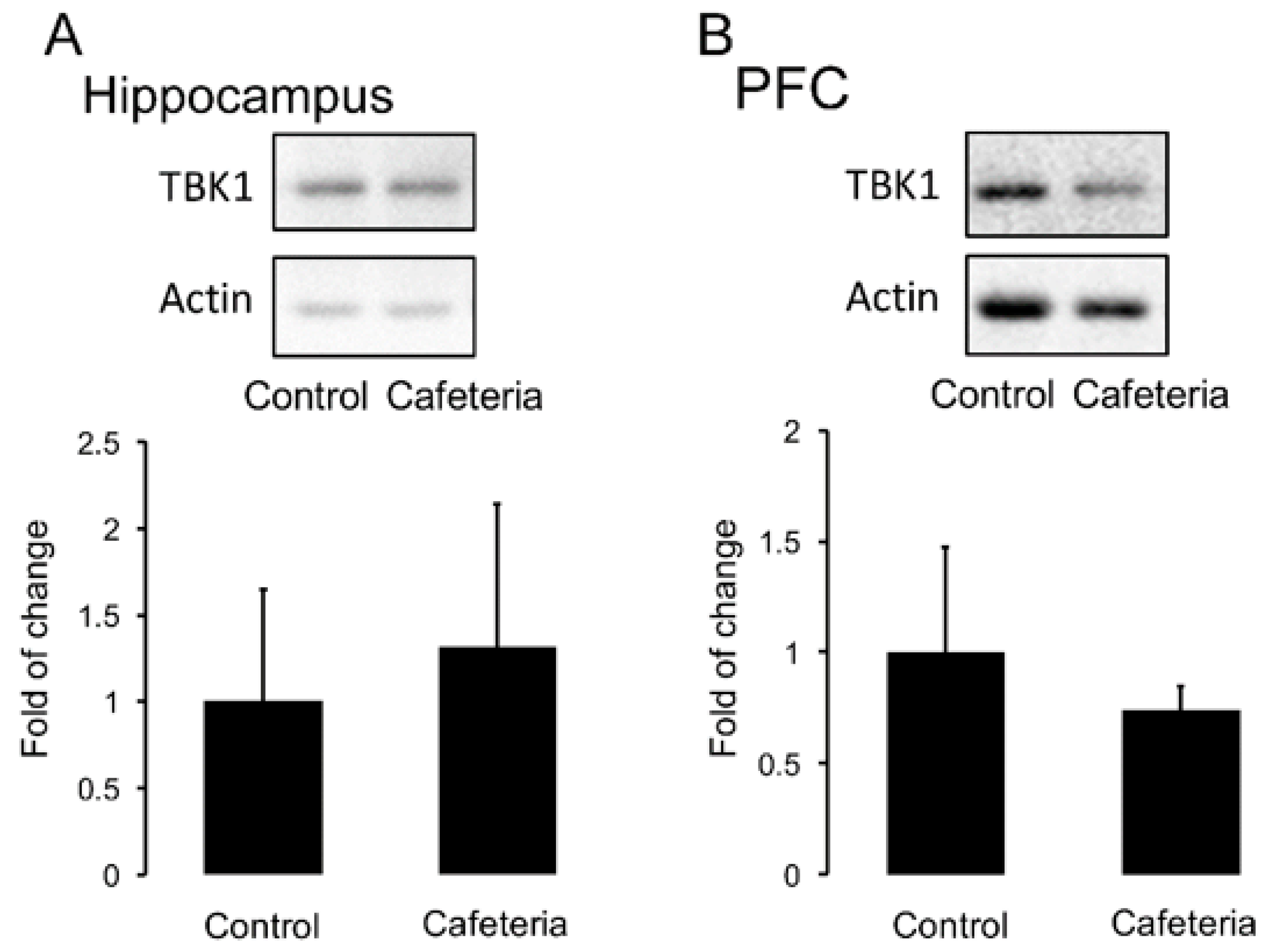
3.3. Flavonoids Revert the Insensitivity to the Anti-Depressant Imipramine Induced by Nutritional Programming by Cafeteria Diet
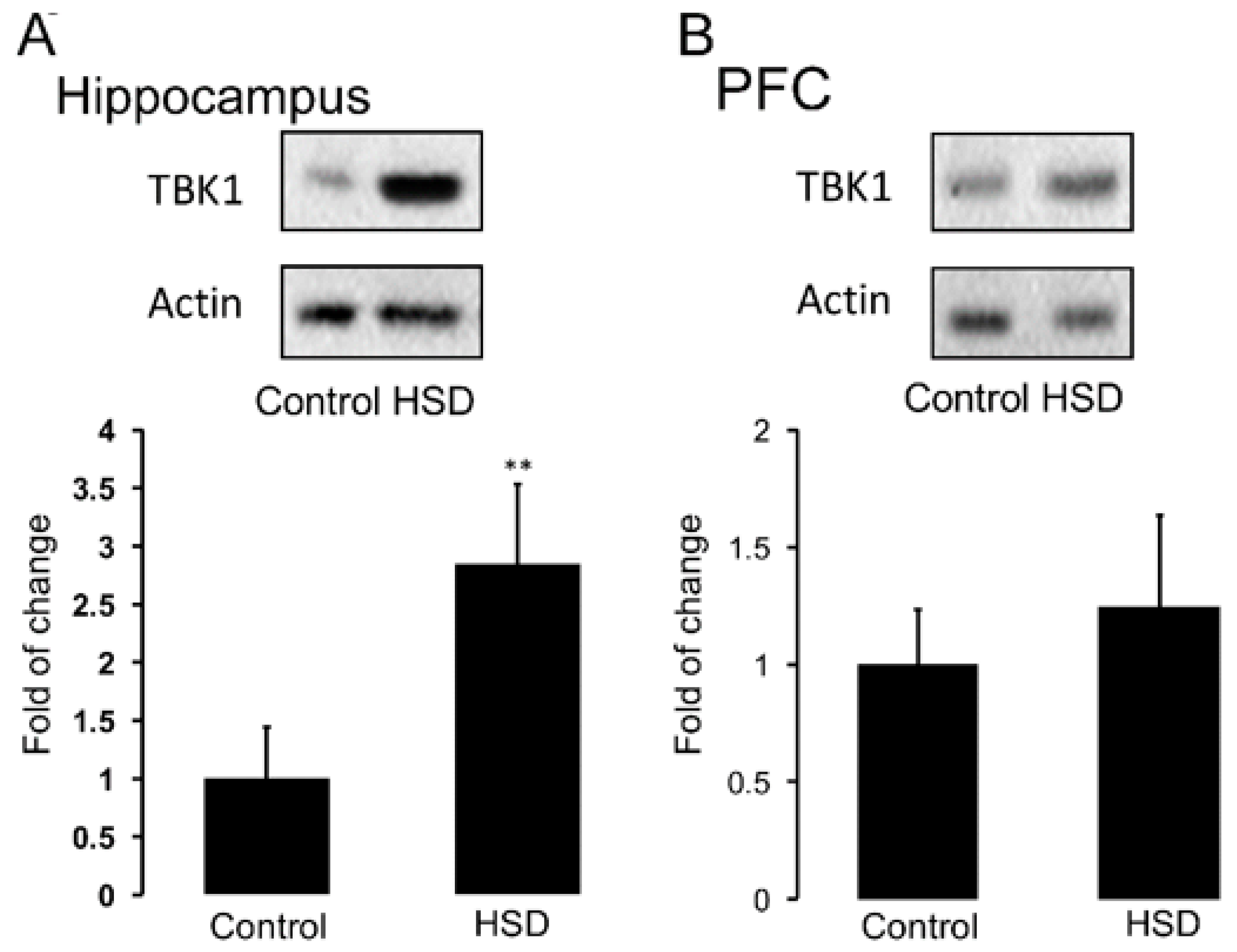
3.4. Nutritional Programming by HSD Promotes an Increase in Hippocampal TBK1 Expression of Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HFD | High fat diet |
| HSD | High sugar diet |
| TBK1 | TANK binding kinase 1 |
| IL | Interleukin |
| ANOVA | Analysis of variance |
References
- Goran, M.I.; Dumke, K.; Bouret, S.G.; Kayser, B.; Walker, R.W.; Blumberg, B. The obesogenic effect of high fructose exposure during early development. Nat. Rev. Endocrinol. 2013, 9, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.N.; Wlodek, M.E.; Moritz, K.M.; Cuffe, J.S.M. Programming of maternal and offspring disease: Impact of growth restriction, fetal sex and transmission across generations. J. Physiol. 2016, 594, 4727–4740. [Google Scholar] [CrossRef] [PubMed]
- Neri, C.; Edlow, A.G. Effects of maternal obesity on fetal programming: Molecular approaches. Cold Spring Harb. Perspect. Med. 2016, 6, a026591. [Google Scholar] [CrossRef] [PubMed]
- Montalvo-Martínez, L.; Maldonado-Ruiz, R.; Cárdenas-Tueme, M.; Reséndez-Pérez, D.; Camacho, A. Maternal Overnutrition Programs Central Inflammation and Addiction-Like Behavior in Offspring. BioMed Res. Int. 2018, 2018, 8061389. [Google Scholar] [CrossRef] [PubMed]
- Atladóttir, H.Ó.; Thorsen, P.; Østergaard, L.; Schendel, D.E.; Lemcke, S.; Abdallah, M.; Parner, E.T. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J. Autism Dev. Disord. 2010, 40, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Vucetic, Z.; Kimmel, J.; Totoki, K.; Hollenbeck, E.; Reyes, T.M. Maternal high-fat diet alters methylation and gene expression of dopamine and opioid-related genes. Endocrinology 2010, 151, 4756–4764. [Google Scholar] [CrossRef] [PubMed]
- Grissom, N.M.; Lyde, R.; Christ, L.; Sasson, I.E.; Carlin, J.; Vitins, A.P.; Simmons, R.A.; Reyes, T.M. Obesity at Conception Programs the Opioid System in the Offspring Brain. Neuropsychopharmacology 2014, 39, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Bellisario, V.; Panetta, P.; Balsevich, G.; Baumann, V.; Noble, J.; Raggi, C.; Nathan, O.; Berry, A.; Seckl, J.; Schmidt, M.; et al. Maternal high-fat diet acts as a stressor increasing maternal glucocorticoids’ signaling to the fetus and disrupting maternal behavior and brain activation in C57BL/6J mice. Psychoneuroendocrinology 2015, 60, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, E.; Reyes-Castro, L.A.; Nathanielsz, P.W. Aging, glucocorticoids and developmental programming. Age 2015, 37, 52. [Google Scholar] [CrossRef] [PubMed]
- Hodes, G.E.; Kana, V.; Menard, C.; Merad, M.; Russo, S.J. Neuroimmune mechanisms of depression. Nat. Neurosci. 2015, 18, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sheng, H.; Bao, Q.; Wang, Y.; Lu, J.; Ni, X. NLRP3 inflammasome activation mediates estrogen deficiency-induced depression- and anxiety-like behavior and hippocampal inflammation in mice. Brain Behav. Immun. 2016, 56, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Krebs, N.F.; Lozoff, B.; Georgieff, M.K. Neurodevelopment: The Impact of Nutrition and Inflammation During Infancy in Low-Resource Settings. Pediatrics 2017, 139, S38–S49. [Google Scholar] [CrossRef] [PubMed]
- Posillico, C.K.; Schwarz, J.M. An investigation into the effects of antenatal stressors on the postpartum neuroimmune profile and depressive-like behaviors. Behav. Brain Res. 2016, 298, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.M.; Li, M.; Gray, C.; Vickers, M.H. Pre-Weaning Growth Hormone Treatment Ameliorates Bone Marrow Macrophage Inflammation in Adult Male Rat Offspring following Maternal Undernutrition. PLoS ONE 2013, 8, e68262. [Google Scholar] [CrossRef] [PubMed]
- Köhler, C.A.; Freitas, T.H.; Maes, M.; de Andrade, N.Q.; Liu, C.S.; Fernandes, B.S.; Stubbs, B.; Solmi, M.; Veronese, N.; Herrmann, N.; et al. Peripheral cytokine and chemokine alterations in depression: A meta-analysis of 82 studies. Acta Psychiatr. Scand. 2017, 135, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.M.; Garakani, A.; Yehuda, R.; Gorman, J.M. Proinflammatory and “resiliency” proteins in the CSF of patients with major depression. Depress. Anxiety 2012, 29, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.S. Regulation of cell death and autophagy by IKK and NF-κB: Critical mechanisms in immune function and cancer. Immunol. Rev. 2012, 246, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Ruiz, R.; Montalvo-Martínez, L.; Fuentes-Mera, L.; Camacho, A. Microglia activation due to obesity programs metabolic failure leading to type two diabetes. Nutr. Diabetes 2017, 7, e254. [Google Scholar] [CrossRef] [PubMed]
- Delint-Ramirez, I.; Maldonado Ruiz, R.; Torre-Villalvazo, I.; Fuentes-Mera, L.; Garza Ocañas, L.; Tovar, A.; Camacho, A. Genetic obesity alters recruitment of TANK-binding kinase 1 and AKT into hypothalamic lipid rafts domains. Neurochem. Int. 2015, 80, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and inflammation: From chemistry to medicine. Pharmacol. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef] [PubMed]
- De la Garza, A.L. The role of flavonoids in the effort to prevent obesity: Nutrition 4.0. Curr. Res. Nutr. Food Sci. J. 2018, 6, 573–575. [Google Scholar] [CrossRef]
- Park, S.E.; Sapkota, K.; Kim, S.; Kim, H.; Kim, S.J. Kaempferol acts through mitogen-activated protein kinases and protein kinase B/AKT to elicit protection in a model of neuroinflammation in BV2 microglial cells. Br. J. Pharmacol. 2011, 164, 1008–1025. [Google Scholar] [CrossRef] [PubMed]
- De la Garza, A.L.; Etxeberria, U.; Palacios-Ortega, S.; Haslberger, A.G.; Aumueller, E.; Milagro, F.I.; Martínez, J.A. Modulation of hyperglycemia and TNFα-mediated inflammation by helichrysum and grapefruit extracts in diabetic db/db mice. Food Funct. 2014, 5, 2120–2128. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Nakashima, T.; Nakamura, K.; Shinoka, W.; Kotani, M.; Sawamoto, A.; Nakajima, M.; Furukawa, Y. Inhibitory effects of auraptene and naringin on astroglial activation, tau hyperphosphorylation, and suppression of neurogenesis in the hippocampus of streptozotocin-induced hyperglycemic mice. Antioxidants 2018, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.D.; Jeong, K.H.; Jung, U.J.; Kim, S.R. Naringin treatment induces neuroprotective effects in a mouse model of Parkinson’s disease in vivo, but not enough to restore the lesioned dopaminergic system. J. Nutr. Biochem. 2016, 28, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Pérez-Gregorio, R.; Soares, S.; Mateus, N.; De Freitas, V. Wine flavonoids in health and disease prevention. Molecules 2017, 22, 292. [Google Scholar] [CrossRef] [PubMed]
- Ruchat, S.M.; Hivert, M.F.; Bouchard, L. Epigenetic programming of obesity and diabetes by in utero exposure to gestational diabetes mellitus. Nutr. Rev. 2013, 71, S88–S94. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, E.L.; Nousen, E.K.; Chamlou, K.A. Maternal high fat diet consumption during the perinatal period programs offspring behavior. Physiol. Behav. 2014, 123, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Yessoufou, A.; Moutairou, K. Maternal diabetes in pregnancy: Early and long-term outcomes on the offspring and the concept of “metabolic memory”. Exp. Diabetes Res. 2011, 2011, 218598. [Google Scholar] [CrossRef] [PubMed]
- Rando, O.J.; Simmons, R.A. I’m eating for two: Parental dietary effects on offspring metabolism. Cell 2015, 161, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Peleg-Raibstein, D.; Sarker, G.; Litwan, K.; Krämer, S.D.; Ametamey, S.M.; Schibli, R.; Wolfrum, C. Enhanced sensitivity to drugs of abuse and palatable foods following maternal overnutrition. Transl. Psychiatry 2016, 6, e911. [Google Scholar] [CrossRef] [PubMed]
- Lemos, C.; Rial, D.; Gonçalves, F.Q.; Pires, J.; Silva, H.B.; Matheus, F.C.; Da Silva, A.C.; Marques, J.M.; Rodrigues, R.J.; Jarak, I.; et al. High sucrose consumption induces memory impairment in rats associated with electrophysiological modifications but not with metabolic changes in the hippocampus. Neuroscience 2016, 315, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.S.; Burgado, J.; Kelly, S.D.; Johnson, Z.P.; Neigh, G.N. High-fructose diet during periadolescent development increases depressive-like behavior and remodels the hypothalamic transcriptome in male rats. Psychoneuroendocrinology 2015, 62, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Clouard, C.; Gerrits, W.J.J.; Kemp, B.; Val-Laillet, D.; Bolhuis, J.E. Perinatal exposure to a diet high in saturated fat, refined sugar and cholesterol affects behaviour, growth, and feed intake in weaned piglets. PLoS ONE 2016, 11, e0154698. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.D.; Reichelt, A.C. Impaired fear extinction retention and increased anxiety-like behaviours induced by limited daily access to a high-fat/high-sugar diet in male rats: Implications for diet-induced prefrontal cortex dysregulation. Neurobiol. Learn. Mem. 2016, 136, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Estrela, D.; da Silva, W.A.M.; Guimaraes, A.T.B.; de Oliveira Mendes, B.; da Silva Castro, A.L.; da Silva Torres, I.L.; Malafaia, G. Predictive behaviors for anxiety and depression in female Wistar rats subjected to cafeteria diet and stress. Physiol. Behav. 2015, 151, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Tonkiss, J.; Shultz, P.; Galler, J.R. Long-evans and sprague-dawley rats differ in their spatial navigation performance during ontogeny and at maturity. Dev. Psychobiol. 1992, 25, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Alkhalidy, H.; Wang, Y.; Liu, D. Dietary flavonoids in the prevention of T2D: An overview. Nutrients 2018, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Meireles, M.; Fernandes, I.; Santos-Buelga, C.; Gonzalez-Manzano, S.; Dueñas, M.; de Freitas, V.; Mateus, N.; Calhau, C. Flavonoid metabolites transport across a human BBB model. Food Chem. 2014, 149, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.S.; Khan, M.M.; Ahmad, A.; Ashafaq, M.; Islam, F.; Wagner, A.P.; Safhi, M.M. Neuroprotective effect of naringenin is mediated through suppression of NF-κB signaling pathway in experimental stroke. Neuroscience 2013, 230, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Ferro Cavalcante, T.C.; Marcelino Da Silva, A.A.; Alheiros Lira, M.C.; Do Amaral Almeida, L.C.; Jaques Marques, A.P.; Do Nascimento, E. Early exposure of dams to a westernized diet has long-term consequences on food intake and physiometabolic homeostasis of the rat offspring. Int. J. Food Sci. Nutr. 2014, 65, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Sim, Y.-B.; Han, P.-L.; Lee, J.-K.; Suh, H.-W. Antidepressant-like Effect of Kaempferol and Quercitirin, Isolated from Opuntia ficus-indica var. saboten. Exp. Neurobiol. 2010, 19, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Winther, G.; Elfving, B.; Müller, H.K.; Lund, S.; Wegener, G. Maternal High-fat Diet Programs Offspring Emotional Behavior in Adulthood. Neuroscience 2018, 388, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Watanabe, W.; Oomagari, H.; Tsuruta, E.; Shida, M.; Kurokawa, M. Citrus flavonoid naringenin inhibits TLR2 expression in adipocytes. J. Nutr. Biochem. 2013, 24, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, D.R.; Rapaport, M.H.; Miller, B.J. A meta-analysis of blood cytokine network alterations in psychiatric patients: Comparisons between schizophrenia, bipolar disorder and depression. Mol. Psychiatry 2016, 21, 1696–1709. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, D.; Janelidze, S.; Hagell, P.; Erhardt, S.; Samuelsson, M.; Minthon, L.; Hansson, O.; Björkqvist, M.; Träskman-Bendz, L.; Brundin, L. Interleukin-6 Is Elevated in the Cerebrospinal Fluid of Suicide Attempters and Related to Symptom Severity. Biol. Psychiatry 2009, 66, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Bonne, O.; Gill, J.M.; Luckenbaugh, D.A.; Collins, C.; Owens, M.J.; Alesci, S.; Neumeister, A.; Yuan, P.; Kinkead, B.; Manji, H.K.; et al. Corticotropin-releasing factor, interleukin-6, brain-derived neurotrophic factor, insulin-like growth factor-1, and substance P in the cerebrospinal fluid of civilians with posttraumatic stress disorder before and after treatment with paroxetine. J. Clin. Psychiatry 2011, 72, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Chau, T.L.; Gioia, R.; Gatot, J.S.; Patrascu, F.; Carpentier, I.; Chapelle, J.P.; O’Neill, L.; Beyaert, R.; Piette, J.; Chariot, A. Are the IKKs and IKK-related kinases TBK1 and IKK-epsilon similarly activated? Trends Biochem. Sci. 2008, 33, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Cirulli, E.T.; Lasseigne, B.N.; Petrovski, S.; Sapp, P.C.; Dion, P.A.; Leblond, C.S.; Couthouis, J.; Lu, Y.F.; Wang, Q.; Krueger, B.J.; et al. Exome sequencing in amyotrophic lateral sclerosis identifies risk genes and pathways. Science 2015, 347, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Freischmidt, A.; Wieland, T.; Richter, B.; Ruf, W.; Schaeffer, V.; Müller, K.; Marroquin, N.; Nordin, F.; Hübers, A.; Weydt, P.; et al. Haploinsufficiency of TBK1 causes familial ALS and fronto-temporal dementia. Nat. Neurosci. 2015, 18, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, K.; Kumari, A.; Radha, V.; Swarup, G. A Glaucoma-Associated variant of optineurin, M98K, activates Tbk1 to enhance autophagosome formation and retinal cell death dependent on Ser177 phosphorylation of optineurin. PLoS ONE 2015, 10, e0138289. [Google Scholar] [CrossRef] [PubMed]
- Dunn, A.J.; Swiergiel, A.H. The role of cytokines in infection-related behavior. Ann. N. Y. Acad. Sci. 1998, 840, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Teeling, J.L.; Felton, L.M.; Deacon, R.M.J.; Cunningham, C.; Rawlins, J.N.P.; Perry, V.H. Sub-pyrogenic systemic inflammation impacts on brain and behavior, independent of cytokines. Brain Behav. Immun. 2007, 21, 836–850. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, P.; Karthikeyan, A.; Dheen, S.T. Role of dietary phenols in mitigating microglia-mediated neuroinflammation. Neuromol. Med. 2016, 18, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Wang, W.; Peng, Y.; Deng, Z. Antidepressive effects of kaempferol mediated by reduction of oxidative stress, proinflammatory cytokines and up-regulation of AKT/β-catenin cascade. Metab. Brain Dis. 2019, 34, 1–10. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | Total Female Before Mating | Pregnant Females | Mean Litter Size After Birth | Males at 21 Days of Age | Females at 21 Days of Age | Ratio Male/Female | Total of f1 Females (Behaviour Test) |
|---|---|---|---|---|---|---|---|
| control | 4 | 4 | 13.75 | 28 | 27 | 1.04 | 20 |
| hfd | 4 | 3 | 14 | 5 | 12 | 0.42 * | 12 |
| cafeteria | 6 | 3 | 13.67 | 12 | 20 | 0.60 | 20 |
| hsd | 4 | 4 | 13.75 | 29 | 26 | 1.12 | 26 |
| cafeteria + flavonoids | 2 | 2 | 12.5 | 9 | 16 | 0.56 | 16 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Garza, A.L.; Garza-Cuellar, M.A.; Silva-Hernandez, I.A.; Cardenas-Perez, R.E.; Reyes-Castro, L.A.; Zambrano, E.; Gonzalez-Hernandez, B.; Garza-Ocañas, L.; Fuentes-Mera, L.; Camacho, A. Maternal Flavonoids Intake Reverts Depression-Like Behaviour in Rat Female Offspring. Nutrients 2019, 11, 572. https://doi.org/10.3390/nu11030572
de la Garza AL, Garza-Cuellar MA, Silva-Hernandez IA, Cardenas-Perez RE, Reyes-Castro LA, Zambrano E, Gonzalez-Hernandez B, Garza-Ocañas L, Fuentes-Mera L, Camacho A. Maternal Flavonoids Intake Reverts Depression-Like Behaviour in Rat Female Offspring. Nutrients. 2019; 11(3):572. https://doi.org/10.3390/nu11030572
Chicago/Turabian Stylede la Garza, Ana Laura, Miguel A. Garza-Cuellar, Ivan A. Silva-Hernandez, Robbi E. Cardenas-Perez, Luis A. Reyes-Castro, Elena Zambrano, Brenda Gonzalez-Hernandez, Lourdes Garza-Ocañas, Lizeth Fuentes-Mera, and Alberto Camacho. 2019. "Maternal Flavonoids Intake Reverts Depression-Like Behaviour in Rat Female Offspring" Nutrients 11, no. 3: 572. https://doi.org/10.3390/nu11030572
APA Stylede la Garza, A. L., Garza-Cuellar, M. A., Silva-Hernandez, I. A., Cardenas-Perez, R. E., Reyes-Castro, L. A., Zambrano, E., Gonzalez-Hernandez, B., Garza-Ocañas, L., Fuentes-Mera, L., & Camacho, A. (2019). Maternal Flavonoids Intake Reverts Depression-Like Behaviour in Rat Female Offspring. Nutrients, 11(3), 572. https://doi.org/10.3390/nu11030572