Improvement in Diabetic Retinopathy through Protection against Retinal Apoptosis in Spontaneously Diabetic Torii Rats Mediated by Ethanol Extract of Osteomeles schwerinae C.K. Schneid

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. OSSCE Preparation
2.2. High-Performance Liquid Chromatography (HPLC) Chromatogram of OSSCE
2.3. Inhibitory Activity on Nonenzymatic AGE Formation
2.4. Inhibitory Activity on AGEs Formation Expression of RAGE in HRMECs
2.5. Animal Experimental Design
2.6. Western Blot Analysis
2.7. Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling (TUNEL) Staining
2.8. IκB Kinase (IKK) Complex Assay
2.9. Morphological Observation of Mitochondria
2.10. Mitochondrial Membrane Potential (ΔΨm) Analysis
2.11. Intracellular ROS Measurement
2.12. Intracellular 8-OHdG Measurements
2.13. Immunostaining
2.14. Measuring Nuclear Factor-κB (NF-κB) Activity
2.15. Measurement of NADPH Oxidase Activity
2.16. Statistical Analysis
3. Results
3.1. HPLC Chromatogram of OSSCE
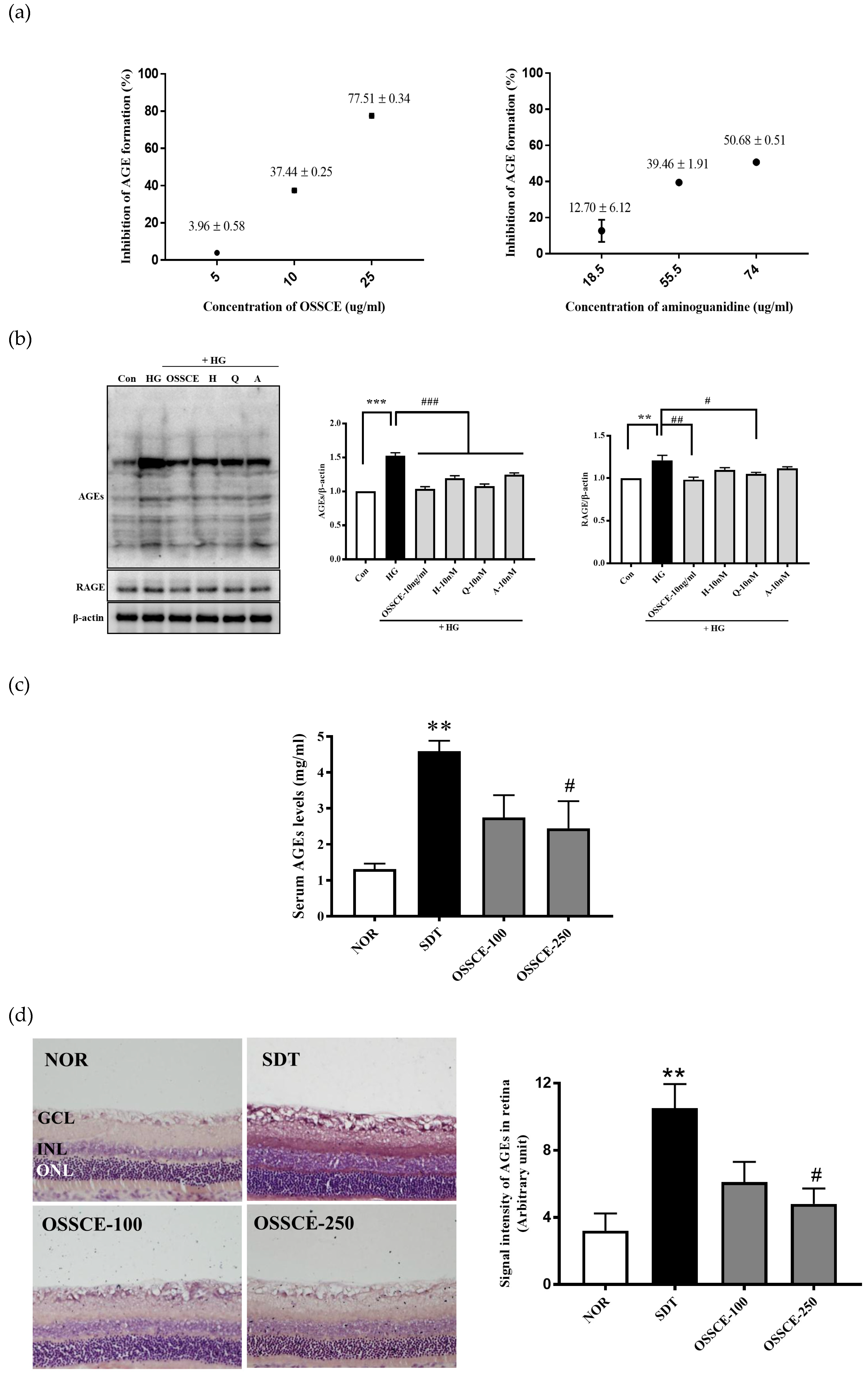
3.2. OSSCE Inhibits Nonenzymatic AGE Formation In Vitro, Expressions of Ages and RAGE in 25 mm Glucose-Treated HRMECs, and AGE Level in Serum and Whole Retina of SDT Rats
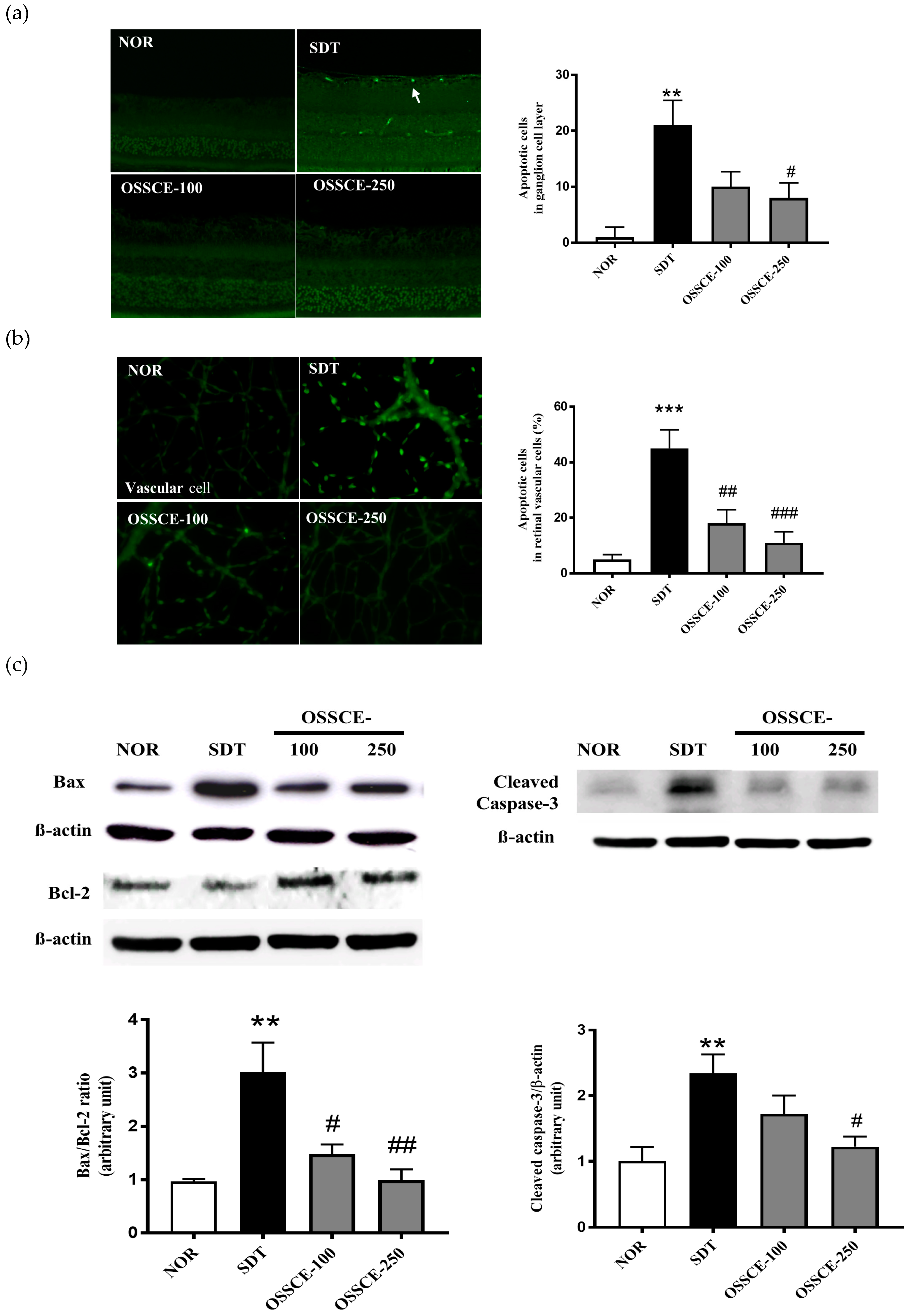
3.3. OSSCE Inhibits Apoptosis of the Retinal Ganglion Cell Layer and Whole Retinal Vessels in SDT Rats
3.4. OSSCE and MCs Inhibit HG-induced Intracellular ROS Generation and 8-OHdG Expression in HRMECs
3.5. Protective Effects of OSSCE and MCs on HG-Induced Mitochondrial Morphology and Mitochondrial Membrane Potential (ΔΨM) in HRMECs
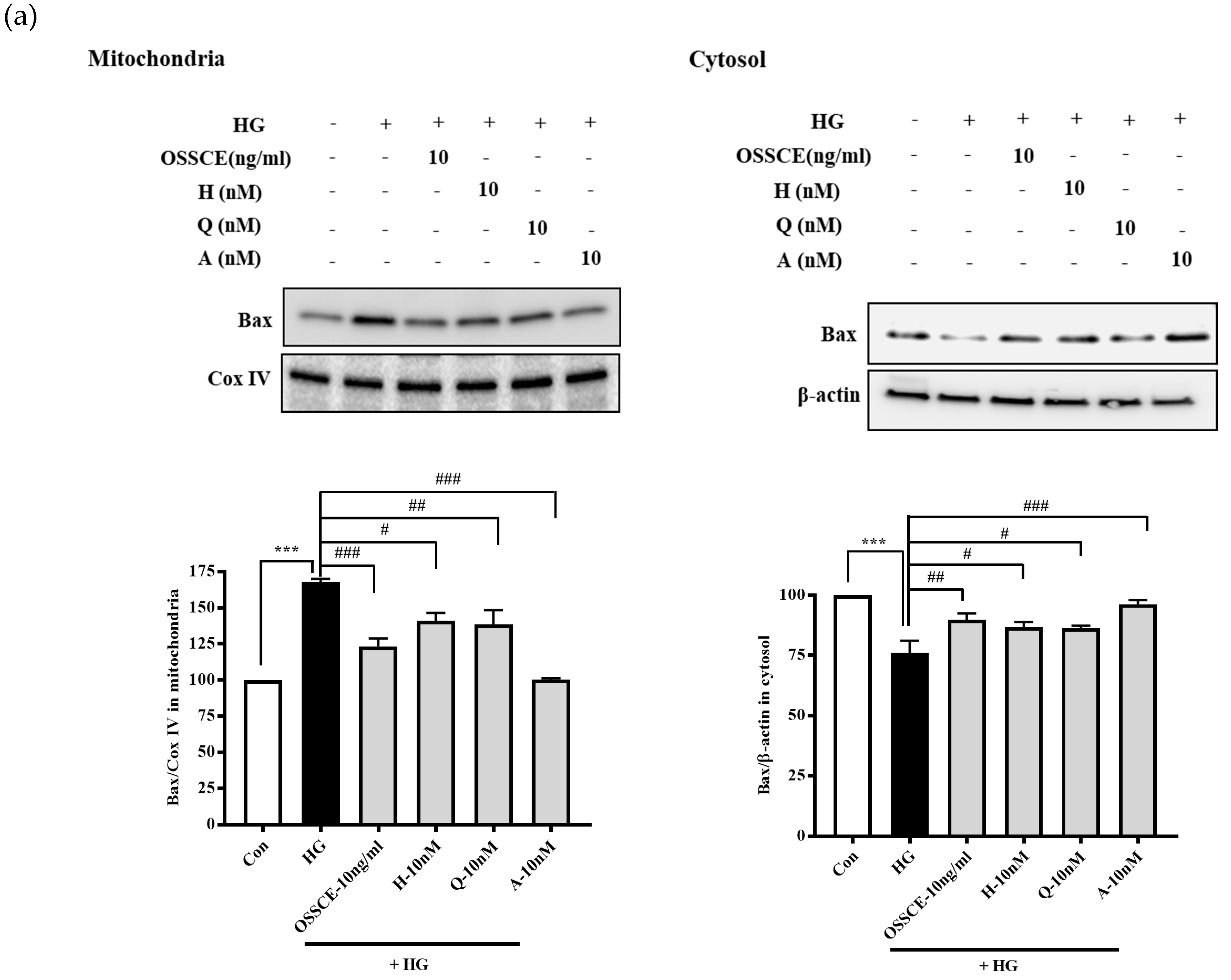
3.6. Effects of OSSCE and MCs on Mitochondria-Dependent Apoptotic Pathways in HG-Treated Hrmecs
3.7. OSSCE Inhibits the Activation of Nuclear Factor Κ-B (NF-Κb) in SDT Rat Retina and HG-Treated Hrmecs
3.8. Effects of OSSCE and MCs on NADPH Oxidase Activity and the Related Signalling Pathways in HG-Treated HRMECs
3.9. Levels of Haemoglobin A1c (Hba1c) and Blood Glucose in SDT Rats
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Conflicts of Interest
Data Availability
References
- Fong, D.S.; Aiello, L.; Gardner, T.W.; King, G.L.; Blankenship, G.; Cavallerano, J.D.; Ferris, F.L.; Klein, R. Diabetic retinopathy. Diabetes Care 2003, 26, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Fraser-Bell, S.; Symes, R.; Vaze, A. Hypertensive eye disease: A review. Clin. Exp. Ophthalmol. 2017, 45, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Barber, A.J. A new view of diabetic retinopathy: A neurodegenerative disease of the eye. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 283–290. [Google Scholar] [CrossRef]
- Barber, A.J.; Lieth, E.; Khin, S.A.; Antonetti, D.A.; Buchanan, A.G.; Gardner, T.W. Neural apoptosis in the retina during experimental and human diabetes. Early onset and effect of insulin. J. Clin. Invest. 1998, 102, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Girach, A.; Manner, D.; Porta, M. Diabetic microvascular complications: Can patients at risk be identified? A review. Int. J. Clin. Pract. 2006, 60, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Vasan, S.; Foiles, P.; Founds, H. Therapeutic potential of breakers of advanced glycation end product-protein crosslinks. Arch. Biochem. Biophys 2003, 419, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. The pathological implications of protein glycation. Clin. Investig. Med. 1995, 18, 275–281. [Google Scholar] [PubMed]
- Koga, K.; Yamagishi, S.; Okamoto, T.; Inagaki, Y.; Amano, S.; Takeuchi, M.; Makita, Z. Serum levels of glucose-derived advanced glycation end products are associated with the severity of diabetic retinopathy in type 2 diabetic patients without renal dysfunction. Int. J. Clin. Pharmacol. Res. 2002, 22, 13–17. [Google Scholar] [PubMed]
- Miura, J.; Yamagishi, S.; Uchigata, Y.; Takeuchi, M.; Yamamoto, H.; Makita, Z.; Iwamoto, Y. Serum levels of non-carboxymethyllysine advanced glycation endproducts are correlated to severity of microvascular complications in patients with Type 1 diabetes. J. Diabetes Complications 2003, 17, 16–21. [Google Scholar] [CrossRef]
- Sharma, I.; Tupe, R.S.; Wallner, A.K.; Kanwar, Y.S. Contribution of myo-inositol oxygenase in AGE: RAGE-mediated renal tubulointerstitial injury in the context of diabetic nephropathy. Am. J. Physiol. Renal. Physiol. 2018, 314, F107–F121. [Google Scholar] [CrossRef] [PubMed]
- Hammes, H.P.; Alt, A.; Niwa, T.; Clausen, J.T.; Bretzel, R.G.; Brownlee, M.; Schleicher, E.D. Differential accumulation of advanced glycation end products in the course of diabetic retinopathy. Diabetologia 1999, 42, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.; Nakamura, K.; Matsui, T.; Inagaki, Y.; Takenaka, K.; Jinnouchi, Y.; Yoshida, Y.; Matsuura, T.; Narama, I.; Motomiya, Y.; et al. Pigment epithelium-derived factor inhibits advanced glycation end product-induced retinal vascular hyperpermeability by blocking reactive oxygen species-mediated vascular endothelial growth factor expression. J. Biol. Chem. 2006, 281, 20213–20220. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Rashid, K.; Chowdhury, S.; Ghosh, S.; Sil, P.C. Curcumin attenuates oxidative stress induced NFkappaB mediated inflammation and endoplasmic reticulum dependent apoptosis of splenocytes in diabetes. Biochem. Pharmacol. 2017, 143, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Hu, L.; Hong, X.J.S.S. Chinese Materia Medica; Technology Press: Shanghai, China, 1999; Volume 4, p. 166. [Google Scholar]
- Lee, J.; Jang, D.S.; Yoo, N.H.; Lee, Y.M.; Kim, J.H.; Kim, J.S. Single-step separation of bioactive flavonol glucosides from Osteomeles schwerinae by high-speed counter-current chromatography. J. Sep. Sci. 2010, 33, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.S.; Jung, S.H.; Lee, Y.M.; Choi, S.J.; Sun, H.; Kim, J.S. Phenolic compounds from the leaves and twigs of osteomeles schwerinae that inhibit rat lens aldose reductase and vessel dilation in zebrafish larvae. J. Nat. Prod. 2015, 78, 2249–2254. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Kim, J.; Kim, C.S.; Jo, K.; Yoo, N.H.; Sohn, E.; Kim, J.S. Anti-glycation and anti-angiogenic activities of 5’-methoxybiphenyl-3,4,3’-triol, a novel phytochemical component of Osteomeles schwerinae. Eur. J. Pharmacol. 2015, 760, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Kim, J.; Jo, K.; Lee, Y.M.; Sohn, E.; Yoo, N.H.; Kim, J.S. OSSC1E-K19, a novel phytochemical component of Osteomeles schwerinae, prevents glycated albumin-induced retinal vascular injury in rats. Mol. Med. Rep. 2015, 12, 7279–7284. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Jung, D.H.; Lee, I.S.; Pyun, B.J.; Kim, J.S. Osteomeles schwerinae extracts inhibits the binding to receptors of advanced glycation end products and TGF-beta1 expression in mesangial cells under diabetic conditions. Phytomedicine 2016, 23, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Sohn, E.; Kim, J.; Kim, C.S.; Jo, K.; Kim, J.S. Osteomeles schwerinae Extract Prevents Diabetes-Induced Renal Injury in Spontaneously Diabetic Torii Rats. Evid. Based Complement. Alternat. Med. 2018, 2018, 6824215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Sethiel, M.S.; Shen, W.; Liao, S.; Zou, Y. Hyperoside downregulates the receptor for advanced glycation end products (RAGE) and promotes proliferation in ECV304 cells via the c-Jun N-terminal kinases (JNK) pathway following stimulation by advanced glycation end-products in vitro. Int. J. Mol. Sci. 2013, 14, 22697–22707. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Jung, D.H.; Lee, I.S.; Choi, S.J.; Yu, S.Y.; Ku, S.K.; Kim, M.H.; Kim, J.S. Effects of Allium victorialis leaf extracts and its single compounds on aldose reductase, advanced glycation end products and TGF-beta1 expression in mesangial cells. BMC Complement. Altern. Med. 2013, 13, 251. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Jung, S.H.; Jung, D.H.; Choi, S.J.; Lee, Y.R.; Kim, J.S. Gas6 stimulates angiogenesis of human retinal endothelial cells and of zebrafish embryos via ERK1/2 signaling. PLoS ONE 2014, 9, e83901. [Google Scholar] [CrossRef] [PubMed]
- Sohn, E.J.; Kim, C.S.; Kim, Y.S.; Jung, D.H.; Jang, D.S.; Lee, Y.M.; Kim, J.S. Effects of magnolol (5,5’-diallyl-2,2’-dihydroxybiphenyl) on diabetic nephropathy in type 2 diabetic Goto-Kakizaki rats. Life Sci. 2007, 80, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Tchobroutsky, G. Relation of diabetic control to development of microvascular complications. Diabetologia 1978, 15, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Tamborlane, W.V.; Puklin, J.E.; Bergman, M.; Verdonk, C.; Rudolf, M.C.; Felig, P.; Genel, M.; Sherwin, R. Long-term improvement of metabolic control with the insulin pump does not reverse diabetic microangiopathy. Diabetes Care 1982, 5, 58–64. [Google Scholar] [PubMed]
- Miao, G.; Ito, T.; Uchikoshi, F.; Kamei, M.; Akamaru, Y.; Kiyomoto, T.; Komoda, H.; Nozawa, M.; Matsuda, H. Stage-dependent effect of pancreatic transplantation on diabetic ocular complications in the Spontaneously Diabetic Torii rat. Transplantation 2004, 77, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Byun, A.R.; Kwon, S. Effect of Yeonryeonggobon-dan (YRGBD), an herbal complex, on glycemic control in patients with Type 2 diabetes mellitus: A case series. Complement. Ther. Med. 2014, 22, 1037–1040. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Moon, E.; Kwon, S. Effect of Astragalus membranaceus extract on diabetic nephropathy. Endocrinol. Diabetes Metab. Case Rep. 2014, 2014, 140063. [Google Scholar] [CrossRef] [PubMed]
- Sasase, T.; Ohta, T.; Ogawa, N.; Miyajima, K.; Ito, M.; Yamamoto, H.; Morinaga, H.; Matsushita, M. Preventive effects of glycaemic control on ocular complications of Spontaneously Diabetic Torii rat. Diabetes Obes. Metab. 2006, 8, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Masuyama, T.; Komeda, K.; Hara, A.; Noda, M.; Shinohara, M.; Oikawa, T.; Kanazawa, Y.; Taniguchi, K. Chronological characterization of diabetes development in male Spontaneously Diabetic Torii rats. Biochem. Biophys. Res. Commun. 2004, 314, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Denis, U.; Lecomte, M.; Paget, C.; Ruggiero, D.; Wiernsperger, N.; Lagarde, M. Advanced glycation end-products induce apoptosis of bovine retinal pericytes in culture: Involvement of diacylglycerol/ceramide production and oxidative stress induction. Free Radic. Biol. Med. 2002, 33, 236–247. [Google Scholar] [CrossRef]
- Yamagishi, S.; Amano, S.; Inagaki, Y.; Okamoto, T.; Koga, K.; Sasaki, N.; Yamamoto, H.; Takeuchi, M.; Makita, Z. Advanced glycation end products-induced apoptosis and overexpression of vascular endothelial growth factor in bovine retinal pericytes. Biochem. Biophys. Res. Commun. 2002, 290, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Lecleire-Collet, A.; Tessier, L.H.; Massin, P.; Forster, V.; Brasseur, G.; Sahel, J.A.; Picaud, S. Advanced glycation end products can induce glial reaction and neuronal degeneration in retinal explants. Br. J. Ophthalmol. 2005, 89, 1631–1633. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death. Dis. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Campello, S.; Scorrano, L. Mitochondrial shape changes: Orchestrating cell pathophysiology. EMBO Rep. 2010, 11, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Rasola, A.; Bernardi, P. The mitochondrial permeability transition pore and its involvement in cell death and in disease pathogenesis. Apoptosis 2007, 12, 815–833. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Karl, B.; Mathew, A.V.; Gangoiti, J.A.; Wassel, C.L.; Saito, R.; Pu, M.; Sharma, S.; You, Y.H.; Wang, L.; et al. Metabolomics reveals signature of mitochondrial dysfunction in diabetic kidney disease. J. Am. Soc. Nephrol. 2013, 24, 1901–1912. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, G.; Simon, L.; Koska, P.; Telek, G.; Nagy, Z. Visualization of mitochondrial membrane potential and reactive oxygen species via double staining. Neurosci. Lett. 2006, 399, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.H.; Shen, S.C.; Hsu, C.S.; Chen, Y.C. Mitochondrial-dependent, reactive oxygen species-independent apoptosis by myricetin: Roles of protein kinase C, cytochrome c, and caspase cascade. Biochem. Pharmacol. 2005, 69, 913–927. [Google Scholar] [CrossRef] [PubMed]
- Al-Shabrawey, M.; Bartoli, M.; El-Remessy, A.B.; Ma, G.; Matragoon, S.; Lemtalsi, T.; Caldwell, R.W.; Caldwell, R.B. Role of NADPH oxidase and Stat3 in statin-mediated protection against diabetic retinopathy. Invest. Ophthalmol. Vis. Sci. 2008, 49, 3231–3238. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, R.P.; Tang, J.; Zhang, J.H. Hyperbaric oxygen suppresses NADPH oxidase in a rat subarachnoid hemorrhage model. Stroke 2006, 37, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Didion, S.P.; Faraci, F.M. Angiotensin II produces superoxide-mediated impairment of endothelial function in cerebral arterioles. Stroke 2003, 34, 2038–2042. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Simeone, M.; Patel, R. Inhibition of NADPH oxidase reduces myocardial oxidative stress and apoptosis and improves cardiac function in heart failure after myocardial infarction. Free Radic. Biol. Med. 2007, 43, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zalewski, A.; Liu, Y.; Mazurek, T.; Cowan, S.; Martin, J.L.; Hofmann, S.M.; Vlassara, H.; Shi, Y. Diabetes-induced oxidative stress and low-grade inflammation in porcine coronary arteries. Circulation 2003, 108, 472–478. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nor | SDT | OSSCE-100 | OSSCE-250 | |
|---|---|---|---|---|
| Blood glucose (mg/dL) | 144.1 ± 21.0 | 419.2 ± 21.1 * | 393.4 ± 47.7 | 391.5 ± 52.2 |
| HbA1c (%) | 3.49 ± 0.07 | 9.13 ± 0.37 * | 9.14 ± 0.30 | 8.74 ± 0.48 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.-S.; Kim, J.; Kim, Y.S.; Jo, K.; Lee, Y.M.; Jung, D.H.; Lee, I.S.; Kim, J.-H.; Kim, J.S. Improvement in Diabetic Retinopathy through Protection against Retinal Apoptosis in Spontaneously Diabetic Torii Rats Mediated by Ethanol Extract of Osteomeles schwerinae C.K. Schneid. Nutrients 2019, 11, 546. https://doi.org/10.3390/nu11030546
Kim C-S, Kim J, Kim YS, Jo K, Lee YM, Jung DH, Lee IS, Kim J-H, Kim JS. Improvement in Diabetic Retinopathy through Protection against Retinal Apoptosis in Spontaneously Diabetic Torii Rats Mediated by Ethanol Extract of Osteomeles schwerinae C.K. Schneid. Nutrients. 2019; 11(3):546. https://doi.org/10.3390/nu11030546
Chicago/Turabian StyleKim, Chan-Sik, Junghyun Kim, Young Sook Kim, Kyuhyung Jo, Yun Mi Lee, Dong Ho Jung, Ik Soo Lee, Joo-Hwan Kim, and Jin Sook Kim. 2019. "Improvement in Diabetic Retinopathy through Protection against Retinal Apoptosis in Spontaneously Diabetic Torii Rats Mediated by Ethanol Extract of Osteomeles schwerinae C.K. Schneid" Nutrients 11, no. 3: 546. https://doi.org/10.3390/nu11030546
APA StyleKim, C.-S., Kim, J., Kim, Y. S., Jo, K., Lee, Y. M., Jung, D. H., Lee, I. S., Kim, J.-H., & Kim, J. S. (2019). Improvement in Diabetic Retinopathy through Protection against Retinal Apoptosis in Spontaneously Diabetic Torii Rats Mediated by Ethanol Extract of Osteomeles schwerinae C.K. Schneid. Nutrients, 11(3), 546. https://doi.org/10.3390/nu11030546