Potential of Schisandra chinensis (Turcz.) Baill. in Human Health and Nutrition: A Review of Current Knowledge and Therapeutic Perspectives

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Biologically Active Compounds in SCE
3. Health Promoting Effects of SCE Constituents
3.1. Antimicrobial Activity
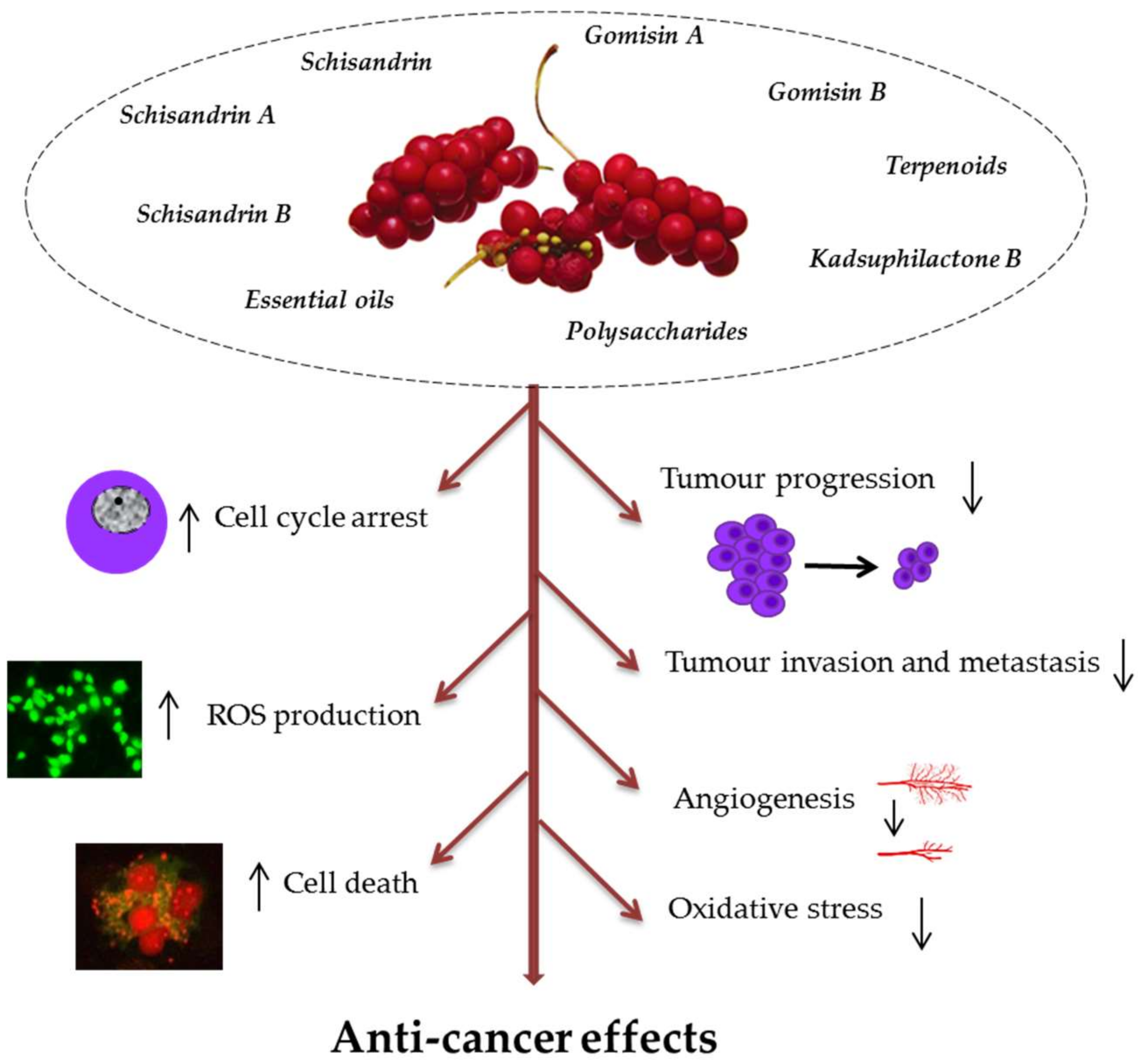
3.2. Anti-Cancer Effect
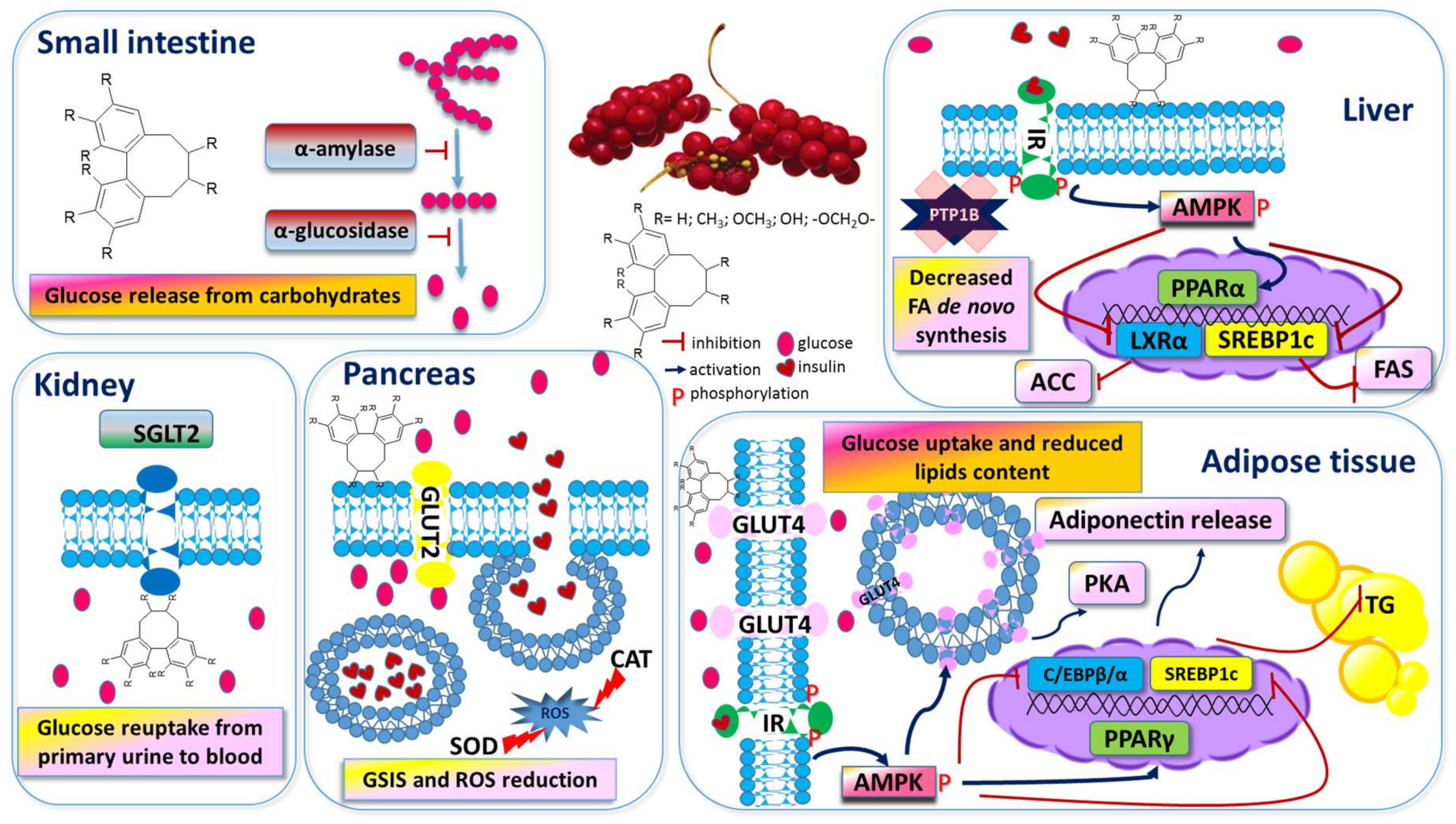
3.3. Anti-Obesity and Anti-Diabetic Action
3.3.1. SCE as A Carbohydrate Metabolism Modulator
3.3.2. SCE as A Lipid Metabolism Modulator
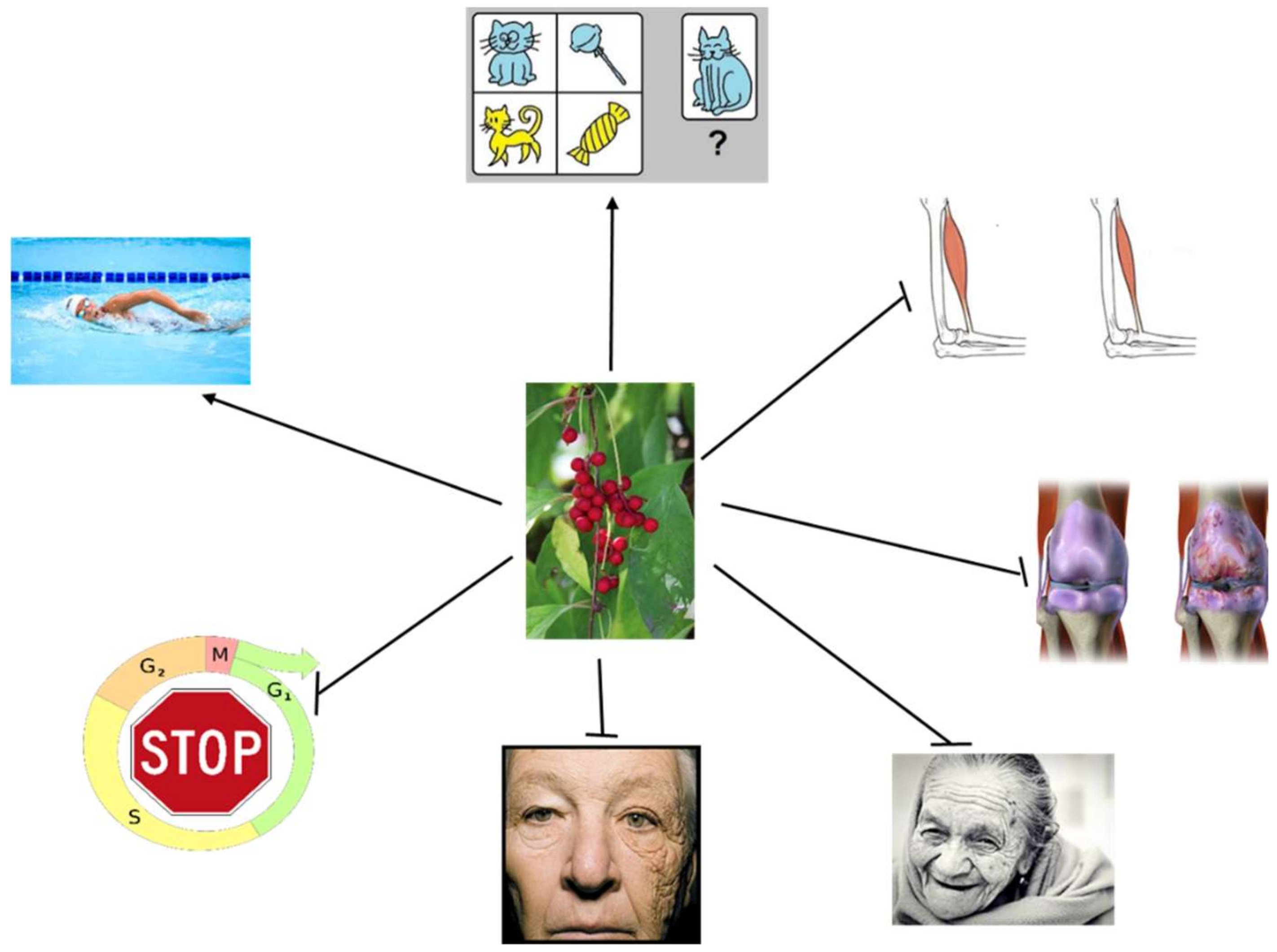
3.4. Aging-Related Effects of SCE
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Saxena, M.; Saxena, J.; Nema, R.; Singh, D.; Gupta, A. Phytochemistry of medicinal plants. J. Pharmacogn. Phytochem. 2013, 1, 168–182. [Google Scholar]
- Da Silva, B.V.; Barreira, J.C.M.; Oliveira, M.B.P.P. Natural phytochemicals and probiotics as bioactive ingredients for functional foods: Extraction, biochemistry and protected-delivery technologies. Trends Food Sci. Technol. 2016, 50, 144–158. [Google Scholar] [CrossRef]
- Panossian, A.; Wikman, G. Pharmacology of Schisandra chinensis Bail.: An overview of Russian research and uses in medicine. J. Ethnopharmacol. 2008, 118, 183–212. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Ekiert, R.; Ekiert, H. Current knowledge of Schisandra chinensis (Turcz.) Baill. (Chinese magnolia vine) as a medicinal plant species: A review on bioactive components, pharmacological properties, analytical and biotechnological studies. Phytochem. Rev. 2017, 16, 195–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, H.; Tian, J.; Chen, S. Antioxidant and anti-proliferative activities of five compounds from Schisandra chinensis fruit. Ind. Crop. Prod. 2013, 50, 690–693. [Google Scholar] [CrossRef]
- Ranouille, E.; Boutot, C.; Bony, E.; Bombarde, O.; Grosjean, S.; Lazewski, A.; Berthon, J.-Y.; Filaire, E. Schisandra chinensis protects the skin from global pollution by inflammatory and redox balance pathway modulations: An in vitro study. Cosmetics 2018, 5, 36. [Google Scholar] [CrossRef]
- Yang, B.Y.; Han, W.; Han, H.; Liu, Y.; Guan, W.; Li, X.M.; Kuang, H.X. Effects of lignans from Schisandra chinensis rattan stems against Aβ1-42-induced memory impairment in rats and neurotoxicity in primary neuronal cells. Molecules 2018, 23, 870. [Google Scholar] [CrossRef]
- Yuan, R.; Tao, X.; Liang, S.; Pan, Y.; He, L.; Sun, J.; Wenbo, J.; Li, X.; Chen, J.; Wang, C. Protective effect of acidic polysaccharide from Schisandra chinensis on acute ethanol-induced liver injury through reducing CYP2E1-dependent oxidative stress. Biomed. Pharmacother. 2018, 99, 537–542. [Google Scholar] [CrossRef]
- Li, Z.; He, X.; Liu, F.; Wang, J.; Feng, J. A review of polysaccharides from Schisandra chinensis and Schisandra sphenanthera: Properties, functions and applications. Carbohydr. Polym. 2018, 184, 178–190. [Google Scholar] [CrossRef]
- Sowndhararajan, K.; Deepa, P.; Kim, M.; Park, S.J.; Kim, S. An Overview of neuroprotective and cognitive enhancement properties of lignans from Schisandra chinensis. Biomed. Pharmacother. 2018, 97, 958–968. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, L.; Yang, H. Schisandra chinensis Fructus and its active ingredients as promising resources for the treatment of neurological diseases. Int. J. Mol. Sci. 2018, 19, 1970. [Google Scholar] [CrossRef] [PubMed]
- Ekiert, R.J.; Szopa, A.; Ekiert, H.; Krzek, J.; Dzik, E. Analysis of lignans in Schisandra chinensis fruits, leaves, biomasses from in vitro cultures and food supplements. J. Funct. Foods 2013, 5, 1576–1581. [Google Scholar] [CrossRef]
- Mocan, A.; Crisan, G.; Vlase, L.; Crissan, O.; Vodnar, D.C.; Raita, O.; Gheldiu, A.M.; Toiu, A.; Oprean, R.; Tilea, I. Comparative studies on polyphenolic composition, antioxidant and antimicrobial activities of Schisandra chinensis leaves and fruits. Molecules 2014, 19, 15162–15179. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Ekiert, H. The importance of applied light quality on the production of lignans and phenolic acids in Schisandra chinensis (Turcz.) Baill. cultures in vitro. Plant Cell Tissue Organ Cult. 2016, 127, 115–121. [Google Scholar] [CrossRef]
- World Health Organisation. WHO Monographs on Selected Medicinal Plants. 2007. Available online: http://apps.who.int/medicinedocs/en/m/abstract/Js14213e/ (accessed on 2 December 2018).
- Xue, Y.B.; Zhang, Y.L.; Yang, J.H.; Du, X.; Pu, J.X.; Zhao, W.; Li, X.N.; Xiao, W.L.; Sun, H.D. Nortriterpenoids and lignans from the fruit of Schisandra chinensis. Chem. Pharm. Bull. 2010, 58, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.Y.; Chen, Z.L.; Liu, Y.; Guo, J.T.; Kuang, H.X. New lignan from the rattan stems of Schisandra chinensis. Nat. Prod. Res. 2018, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Inbathamizh, L.; Padmini, E. Effect of geographical properties on the phytochemical composition and antioxidant potential of Moringa oleifera flowers. BioMedRx 2013, 1, 239–247. [Google Scholar]
- Zhang, H.; Zhang, G.; Zhu, Z.; Zhao, L.; Fei, Y.; Jing, J.; Chai, Y. Determination of six lignans in Schisandra chinensis (Turcz.) Baill. fruits and related Chinese multiherb remedies by HPLC. Food Chem. 2009, 115, 735–739. [Google Scholar] [CrossRef]
- Liu, H.; Lai, H.; Jia, X.; Liu, J.; Zhang, Z.; Qi, Y.; Zhang, J.; Song, J.; Wu, C.; Zhang, B.; et al. Comprehensive chemical analysis of Schisandra chinensis by HPLC-DAD-MS combined with chemometrics. Phytomedicine 2013, 20, 1135–1143. [Google Scholar] [CrossRef]
- Wang, X.; Yu, J.; Li, W.; Wang, C.; Li, H.; Ju, W.; Chen, J.; Sun, J. Characteristics and antioxidant activity of lignans in Schisandra chinensis and Schisandra sphenanthera from different locations. Chem. Biodivers. 2018, 15, e1800030. [Google Scholar] [CrossRef]
- Xia, Y.G.; Yang, B.Y.; Kuang, H.X. Schisandraceae triterpenoids: A review. Phytochem. Rev. 2015, 14, 155–187. [Google Scholar] [CrossRef]
- Yang, S.; Shan, L.; Luo, H.; Seng, X.; Du, J.; Li, Y. Rapid classification and identification of chemical components of Schisandra chinensis by UPLC-Q-TOF/MS combined with data post-processing. Molecules 2017, 22, 1778. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.X.; Han, Q.B.; Lei, C.; Pu, J.X.; Xiao, W.L.; Yu, J.L.; Yang, L.M.; Xu, H.X.; Zheng, Y.T.; Sun, H.D. Isolation and characterization of miscellaneous terpenoids of Schisandra chinensis. Tetrahedron 2008, 64, 4260–4267. [Google Scholar] [CrossRef]
- Shi, Y.M.; Wang, L.Y.; Zou, X.S.; Li, X.N.; Shang, S.Z.; Gao, Z.H.; Liang, C.Q.; Luo, H.R.; Li, H.L.; Xiao, W.L.; et al. Nortriterpenoids from Schisandra chinensis and their absolute configurational assignments by electronic circular dichroism study. Tetrahedron 2014, 70, 859–868. [Google Scholar] [CrossRef]
- Song, Q.-Y.; Gao, K.; Nan, Z.-B. Highly oxygenated triterpenoids from the roots of Schisandra chinensis and their anti-inflammatory activities. J. Asian Nat. Prod. Res. 2016, 18, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.Y.; Chen, Z.L.; Liu, Y.; Kuang, H.X. Three new nortriterpenoids from the rattan stems of Schisandra chinensis. Phytochem. Lett. 2018, 24, 145–149. [Google Scholar] [CrossRef]
- Mocan, A.; Zengin, G.; Crisan, G.; Mollica, A. Enzymatic assays and molecular modeling studies of Schisandra chinensis lignans and phenolics from fruit and leaf extracts. J. Enzyme Inhib. Med. Chem. 2016, 31 (Suppl. 4), 200–210. [Google Scholar] [CrossRef]
- Szopa, A.; Kokotkiewicz, A.; Bednarz, M.; Luczkiewicz, M.; Ekiert, H. Studies on the accumulation of phenolic acids and flavonoids in different in vitro culture systems of Schisandra chinensis (Turcz.) Baill. using a DAD-HPLC method. Phytochem. Lett. 2017, 20, 462–469. [Google Scholar] [CrossRef]
- Ma, C.; Yang, L.; Yang, F.; Wang, W.; Zhao, C.; Zu, Y. Content and color stability of anthocyanins isolated from Schisandra chinensis fruit. Int. J. Mol. Sci. 2012, 13, 14294–14310. [Google Scholar] [CrossRef]
- Liao, J.; Zang, J.; Yuan, F.; Liu, S.; Zhang, Y.; Li, H.; Piao, Z.; Li, H. Identification and analysis of anthocyanin components in fruit color variation in Schisandra chinensis. J. Sci. Food Agric. 2016, 96, 3213–3219. [Google Scholar] [CrossRef]
- Yue, D.; Yang, L.; Liu, S.; Li, J.; Li, W.; Ma, C. A continuous procedure based on column chromatography to purify anthocyanins from Schisandra chinensis by a macroporous resin plus gel filtration chromatography. Molecules 2016, 21, 204. [Google Scholar] [CrossRef] [PubMed]
- Li, X.N.; Cui, H.; Song, Y.Q.; Liang, Y.Z.; Chau, F.T. Analysis of volatile fractions of Schisandra chinensis (Turcz.) Baill. using GC-MS and chemometric resolution. Phytochem. Anal. 2003, 14, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Sowndhararajan, K.; Kim, T.; Kim, H.; Kim, S. Evaluation of proximate composition, bioactive lignans and volatile composition of Schisandra chinensis fruits from Inje and Mungyeong, Republic of Korea. J. Appl. Pharm. Sci. 2016, 6, 001–008. [Google Scholar] [CrossRef]
- Xu, C.L.; Li, Y.H.; Dong, M.; Wu, X.; Wang, X.C.; Xiao, X.S. Inhibitory effect of Schisandra chinensis leaf polysaccharide against L5178Y lymphoma. Carbohydr. Polym. 2012, 88, 21–25. [Google Scholar] [CrossRef]
- Ye, C.; Han, N.; Teng, F.; Wang, X.; Xue, R.; Yin, J. Extraction optimization of polysaccharides of Schisandrae Fructus and evaluation of their analgesic activity. Int. J. Biol. Macromol. 2013, 57, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Mao, G.; Mao, R.; Zou, Y.; Zheng, D.; Feng, W.; Ren, Y.; Wang, W.; Zheng, W.; Song, J.; et al. Antitumor and immunomodulatory activity of a water-soluble low molecular weight polysaccharide from Schisandra chinensis (Turcz.) Baill. Food Chem. Toxicol. 2013, 55, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Feng, Y.; Li, J.; Mao, R.; Zou, Y.; Feng, W.; Zheng, D.; Wang, W.; Chen, Y.; Yang, L.; et al. Schisandra polysaccharide evokes immunomodulatory activity through TLR 4-mediated activation of macrophages. Int. J. Biol. Macromol. 2014, 65, 33–40. [Google Scholar] [CrossRef]
- Tong, H.; Zhao, B.; Du, F.; Tian, D.; Feng, K.; Sun, X. Isolation and physicochemical characterization of polysaccharide fractions isolated from Schisandra chinensis. Chem. Nat. Compd. 2012, 47, 969–970. [Google Scholar] [CrossRef]
- Chi, A.; Zhang, Y.; Kang, Y.; Shen, Z. Metabolic mechanism of a polysaccharide from Schisandra chinensis to relieve chronic fatigue syndrome. Int. J. Biol. Macromol. 2016, 93, 322–332. [Google Scholar] [CrossRef]
- Zhong, S.; Liu, X.D.; Nie, Y.C.; Gan, Z.Y.; Yang, L.Q.; Huang, C.Q.; Lai, K.F.; Zhong, N.S. Antitussive activity of the Schisandra chinensis fruit polysaccharide (SCFP-1) in guinea pigs models. J. Ethnopharmacol. 2016, 194, 378–385. [Google Scholar] [CrossRef]
- Regulation (EC) No 1924/2006 of the European Parliament and of the Council of 20 December 2006 on Nutrition and Health Claims Made on Foods. OJ L 404, 30 December 2006; pp. 9–25. Available online: https://eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX%3A32006R1924 (accessed on 18 December 2018).
- Teng, H.; Lee, W.Y. Antibacterial and antioxidant activities and chemical compositions of volatile oils extracted from Schisandra chinensis Baill. seeds using simultaneous distillation extraction method, and comparison with soxhlet and microwave-assisted extraction. Biosci. Biotechnol. Biochem. 2014, 78, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Hakala, E.; Hanski, L.; Uvell, H.; Yrjönen, T.; Vuorela, H.; Elofsson, M.; Vuorela, P.M. Dibenzocyclooctadiene lignans from Schisandra spp. selectively inhibit the growth of the intracellular bacteria Chlamydia pneumoniae and Chlamydia trachomatis. J. Antibiot. 2015, 68, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Park, B.; Hwang, H.-J.; Mah, J.-H. The ability of Schisandra chinensis fruit to inhibit the growth of foodborne pathogenic bacteria and the viability and heat resistance of Bacillus cereus spores. Int. J. Food Sci. Technol. 2015, 50, 2193–2200. [Google Scholar] [CrossRef]
- Choi, E.J.; Jang, S.R.; Kang, O.J.; Bang, W.S. Antimicrobial activity of Psoralea corylifolia, Schisandra chinensis, and Spatholobus suberectus extracts. Korean J. Food Sci. Technol. 2013, 45, 495–500. [Google Scholar] [CrossRef]
- Yu, H. Antimicrobial activity and mechanism of Schisandra chinensis extract. In Proceedings of the 5th International Conference on Environment, Materials, Chemistry and Power Electronics (EMCPE 2016), Zhengzhou, China, 11–12 April 2016; Ankang University: Ankang, China; pp. 192–196. [Google Scholar]
- Min, K.J.; Kwon, T.K. Anticancer effects and molecular mechanisms of epigallocatechin-3-gallate. Integr. Med. Res. 2014, 3, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Yim, S.Y.; Lee, Y.J.; Lee, Y.K.; Jung, S.E.; Kim, J.H.; Kim, H.J.; Son, B.G.; Park, Y.H.; Lee, Y.G.; Choi, Y.W.; et al. Gomisin N isolated from Schisandra chinensis significantly induces anti-proliferative and pro-apoptotic effects in hepatic carcinoma. Mol. Med. Rep. 2009, 2, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Venkanna, A.; Siva, B.; Poornima, B.; Vadaparthi, P.R.; Prasad, K.R.; Reddy, K.A.; Reddy, G.B.; Babu, K.S. Phytochemical investigation of sesquiterpenes from the fruits of Schisandra chinensis and their cytotoxic activity. Fitoterapia 2014, 95, 102–108. [Google Scholar] [CrossRef]
- Lv, X.J.; Zhao, L.J.; Hao, Y.Q.; Su, Z.Z.; Li, J.Y.; Du, Y.W.; Zhang, J. Schisandrin B inhibits the proliferation of human lung adenocarcinoma A549 cells by inducing cycle arrest and apoptosis. Int. J. Clin. Exp. Med. 2015, 8, 6926–6936. [Google Scholar] [CrossRef]
- Dai, X.; Yin, C.; Guo, G.; Zhang, Y.; Zhao, C.; Qian, J.; Wang, O.; Zhang, X.; Liang, G. Schisandrin B exhibits potent anticancer activity in triple negative breast cancer by inhibiting STAT3. Toxicol. Appl. Pharmacol. 2018, 358, 110–119. [Google Scholar] [CrossRef]
- Desai, A.G.; Qazi, G.N.; Ganju, R.K.; El-Tamer, M.; Singh, J.; Saxena, A.K.; Bedi, Y.S.; Taneja, S.C.; Bhat, H.K. Medicinal plants and cancer chemoprevention. Curr. Drug Metab. 2008, 9, 581–591. [Google Scholar] [CrossRef]
- Kee, J.Y.; Han, Y.H.; Mun, J.G.; Park, S.H.; Jeon, H.D.; Hong, S.H. Gomisin a suppresses colorectal lung metastasis by inducing AMPK/P38-mediated apoptosis and decreasing metastatic abilities of colorectal cancer cells. Front. Pharmacol. 2018, 9, 986. [Google Scholar] [CrossRef] [PubMed]
- Casarin, E.; Dall’Acqua, S.; Smejka, K.; Slapetová, T.; Innocenti, G.; Carrara, M. Molecular mechanisms of antiproliferative effects induced by schisandra-derived dibenzocyclooctadiene lignans (+)-deoxyschisandrin and (-)-gomisin N in human tumour cell lines. Fitoterapia 2014, 98, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shi, S.; Wang, H.; Li, N.; Su, J.; Chou, G.; Wang, S. A homogeneous polysaccharide from Fructus Schisandra chinensis (Turz.) Baill. induces mitochondrial apoptosis through the hsp90/AKT signalling pathway in HepG2 cells. Int. J. Mol. Sci. 2016, 17, 1015. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Ahn, J.H.; Lee, K.T.; Jang, D.S.; Choi, J.H. Deoxyschizandrin, isolated from Schisandra berries, induces cell cycle arrest in ovarian cancer cells and inhibits the protumoural activation of tumour-associated macrophages. Nutrients 2018, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Chiu, P.Y.; Luk, K.F.; Leung, H.Y.; Ng, K.M.; Ko, K.M. Schisandrin B stereoisomers protect against hypoxia/reoxygenation-induced apoptosis and inhibit associated changes in Ca2+-induced mitochondrial permeability transition and mitochondrial membrane potential in H9c2 cardiomyocytes. Life Sci. 2008, 82, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Thandavarayan, R.A.; Giridharan, V.V.; Arumugam, S.; Suzuki, K.; Ko, K.M.; Krishnamurthy, P.; Watanabe, K.; Konishi, T. Schisandrin B prevents doxorubicin induced cardiac dysfunction by modulation of DNA damage, oxidative stress and inflammation through inhibition of MAPK/P53 signaling. PLoS ONE 2015, 10, e0119214. [Google Scholar] [CrossRef] [PubMed]
- Justus, C.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L. In vitro cell migration and invasion assays. J. Vis. Exp. 2014, 88. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, B.; Liu, K.; Ding, Z.; Hu, X. Schisandrin B attenuates cancer invasion and metastasis via inhibiting epithelial-mesenchymal transition. PLoS ONE 2012, 7, e40480. [Google Scholar] [CrossRef]
- Qu, H.M.; Liu, S.J.; Zhang, C.Y. Antitumor and antiangiogenic activity of Schisandra chinensis polysaccharide in a renal cell carcinoma model. Int. J. Biol. Macromol. 2014, 66, 52–56. [Google Scholar] [CrossRef]
- Liu, S.J.; Qu, H.M.; Ren, Y.P. SCP, a polysaccharide from Schisandra chinensis, induces apoptosis in human renal cell carcinoma Caki-1 cells through mitochondrial-dependent pathway via inhibition of ERK activation. Tumor Biol. 2014, 35, 5369–5374. [Google Scholar] [CrossRef]
- Yu, G.J.; Choi, I.W.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Kim, C.M.; Kim, W.J.; Yoo, Y.H.; Choi, Y.H. Induction of reactive oxygen species-mediated apoptosis by purified Schisandrae semen essential oil in human leukemia U937 cells through activation of the caspase cascades and nuclear relocation of mitochondrial apoptogenic factors. Nutr. Res. 2015, 35, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, S.; Park, B.K.; Lee, S.I.; Lim, Y.; Lee, K.; Kwon, H.J. Gomisin G inhibits the growth of triple-negative breast cancer cells by suppressing AKT phosphorylation and decreasing cyclin D1. Biomol. Ther. 2018, 26, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Gao, W.; Wang, D.; Liu, Q.; Zheng, S.; Wang, Y. The protecting effect of deoxyschisandrin and schisandrin B on HaCaT cells against UVB-induced damage. PLoS ONE 2015, 10, e0127177. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, J.H.; Park, H.S.; Lee, G.S.; Kim, H.W.; Ha, K.T.; Kim, B.J. Schizandra chinensis extracts induce apoptosis in human gastric cancer cells via JNK/P38 MAPK activation and the ROS-mediated/mitochondria-dependent pathway. Pharm. Biol. 2015, 53, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Suh, W.S.; Park, S.Y.; Min, B.S.; Kim, S.H.; Song, J.H.; Shim, S.H. The antiproliferative effects of compounds isolated from Schisandra chinensis. Korean J. Food Sci. Technol. 2014, 46, 665–670. [Google Scholar] [CrossRef]
- Jeong, M.; Kim, H.M.; Kim, H.J.; Choi, J.H.; Jang, D.S. Kudsuphilactone B, a nortriterpenoid isolated from Schisandra chinensis fruit, induces caspase-dependent apoptosis in human ovarian cancer A2780 cells. Arch. Pharm. Res. 2017, 40, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhong, F.; Zhao, C.; Liu, S.; Xiao, H. The modulatory effect of Schisandra chinensis seed extract on cisplatin induced cytotoxicity and cell proliferation inhibition in human osteosarcoma. Biomed. Res. 2017, 28, 1851–1854. [Google Scholar]
- Liu, H.; Wu, C.; Wang, S.; Gao, S.; Liu, J.; Dong, Z.; Zhang, B.; Liu, M.; Sun, X.; Guo, P. Extracts and lignans of Schisandra chinensis fruit alter lipid and glucose metabolism in vivo and in vitro. J. Funct. Foods 2015, 19, 296–307. [Google Scholar] [CrossRef]
- Zakłos-Szyda, M.; Majewska, I.; Redzynia, M.; Koziołkiewicz, M. Antidiabetic effect of polyphenolic extracts from selected edible plants as α-amylase α-glucosidase and PTP1B inhibitors and β- pancreatic cells cytoprotective agents—A comparative study. Curr. Top. Med. Chem. 2015, 15, 2431–2444. [Google Scholar] [CrossRef]
- Jo, S.-H.; Ha, K.-S.; Moon, K.-S.; Lee, O.-H.; Jang, H.-D.; Kwon, Y.-I. In vitro and in vivo anti-hyperglycemic effects of omija (Schizandra chinensis) fruit. Int. J. Mol. Sci. 2011, 12, 1359–1370. [Google Scholar] [CrossRef]
- Qu, Y.; Chan, Y.Y.-W.; Wong, C.-W.; Cheng, L.; Xu, C.; Leung, A.W.-N.; Lau, C.B.-S. Antidiabetic effect of Schisandrae chinensis Fructus involves inhibition of the sodium glucose cotransporter. Drug Dev. Res. 2015, 76, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Zhao, T.; Feng, W.W.; Mao, G.H.; Zou, Y.; Wang, W.; Li, Q.; Chen, Y.; Wang, X.T.; Yang, L.Q.; et al. Schisandra polysaccharide increased glucose consumption by up-regulating the expression of GLUT-4. Int. J. Biol. Macromol. 2016, 87, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Asp. Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.F.; Chen, B.Q.; Zhu, S.J.; Jiang, L.; Zhou, Y.F. Effects of GLUT4 expression on insulin resistance in patients with advanced liver cirrhosis. J. Zhejiang Univ. Sci. B 2011, 12, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. Regulation of AMP-activated protein kinase by natural and synthetic activators. Acta Pharm. Sin. B 2016, 6, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.L.; Cao, J.Q.; Duan, L.L.; Tang, Y.Y.; Zhao, Q. Protein tyrosine phosphatase 1B (PTP1B) and α-glucosidase inhibitory activities of Schisandra chinensis (Turcz.) Baill. J. Funct. Foods 2014, 9, 264–270. [Google Scholar] [CrossRef]
- An, L.; Wang, Y.; Wang, C.; Fan, M.; Han, X.; Xu, G.; Yuan, G.; Li, H.; Sheng, Y.; Wang, M.; et al. Protective effect of Schisandrae chinensis oil on pancreatic β-cells in diabetic rats. Endocrine 2015, 48, 818–825. [Google Scholar] [CrossRef]
- Park, S.; Hong, S.M.; Ahn, I.S.; Kim, Y.I.; Lee, J.B. Huang-Lian-Jie-Du-Tang supplemented with Schisandra chinensis Baill. and Polygonatum odoratum druce improved glucose tolerance by potentiating insulinotropic actions in islets in 90% pancreatectomized diabetic rats. Biosci. Biotechnol. Biochem. 2009, 73, 2384–2392. [Google Scholar] [CrossRef]
- Zhang, R.-J.; Hao, H.-Y.; Liu, Q.-L.; Zuo, H.-Y.; Chang, Y.-N.; Zhi, Z.-J.; Guo, P.-P.; Hao, Y.-M. Protective effects of Schisandrin on high glucose-induced changes of RhoA and eNOS activity in human umbilical vein endothelial cells. Pathol. Res. Pract. 2018, 214, 1324–1329. [Google Scholar] [CrossRef]
- Kwan, H.Y.; Wu, J.; Su, T.; Chao, X.-J.; Yu, H.; Liu, B.; Fu, X.; Tse, A.K.W.; Chan, C.L.; Fong, W.F.; et al. Schisandrin B regulates lipid metabolism in subcutaneous adipocytes. Sci. Rep. 2017, 7, 10266. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Kimb, D.S.; Yanga, Y.J.; Park, S. The lignan-rich fractions of Fructus Schisandrae improve insulin sensitivity via the PPAR-γ pathways in in vitro and in vivo studies. J. Ethnopharmacol. 2011, 135, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Cho, J.Y.; Kim, M.K.; Koh, P.O.; Cho, K.W.; Kim, C.H.; Lee, K.S.; Chung, B.Y.; Kim, G.S.; Cho, J.H. Anti-obesity effect of Schisandra chinensis in 3T3-L1 cells and high fat diet-induced obese rats. Food Chem. 2012, 134, 227–234. [Google Scholar] [CrossRef]
- Liu, X.; Wang, S.; Wu, Z.; Wang, Z.; Zheng, Q.; Li, D. Deoxyschizandrin loaded liposomes on the suppression lipid accumulation in 3T3-L1 adipocytes. Molecules 2018, 23, 2158. [Google Scholar] [CrossRef] [PubMed]
- Na, M.K.; Hung, T.M.; Oh, W.K.; Min, B.S.; Lee, S.H.; Bae, K.H. Fatty acid synthase inhibitory activity of dibenzocyclooctadiene lignans isolated from Schisandra chinensis. Phytother. Res. 2010, 24 (Suppl. 2), 2008–2011. [Google Scholar] [CrossRef]
- Zhu, P.-L.; Pan, S.-Y.; Zhou, S.-F.; Zhang, Y.; Wang, X.-Y.; Sun, N.; Chu, Z.-S.; Yu, Z.-L.; Ko, K-M. Effects of combined dietary supplementation with fenofibrate and Schisandrae Fructus pulp on lipid and glucose levels and liver function in normal and hypercholesterolemic mice. Drug Des. Dev. Ther. 2015, 9, 923–935. [Google Scholar] [CrossRef]
- Jang, M.K.; Nama, J.S.; Kim, J.H.; Yun, Y.R.; Han, C.W.; Kim, B.J.; Jeong, H.S.; Ha, K.T.; Jung, M.H. Schisandra chinensis extract ameliorates nonalcoholic fatty liver via inhibition of endoplasmic reticulum stress. J. Ethnopharmacol. 2016, 185, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.-Y.; Shin, E.J.; Choi, H.-K.; Kim, S.H.; Sung, M.J.; Park, J.H.; Hwang, J.-T. Schisandra chinensis berry extract protects against steatosis by inhibiting histone acetylation in oleic acid–treated HepG2 cells and in the livers of diet-induced obese mice. Nutr. Res. 2017, 46, 1–10. [Google Scholar] [CrossRef]
- Yun, Y.-R.; Kim, J.-H.; Kim, J.H.; Jung, M.H. Protective effects of gomisin n against hepatic steatosis through AMPK activation. Biochem. Biophys. Res. Commun. 2017, 482, 1095–1101. [Google Scholar] [CrossRef]
- Wang, C.-M.; Yuan, R.-S.; Zhuang, W.-Y.; Sun, J.-H.; Wu, J.-Y.; Li, H.; Chen, J.-G. Schisandra polysaccharide inhibits hepatic lipid accumulation by downregulating expression of SREBPs in NAFLD mice. Lipids Health Dis. 2016, 15, 195. [Google Scholar] [CrossRef]
- Jeyapalan, J.C.; Sedivy, J.M. Cellular senescence and organismal aging. Mech. Ageing Dev. 2008, 129, 467–474. [Google Scholar] [CrossRef]
- Campisi, J.; Robert, L. Cell senescence: Role in aging and age-related diseases. Interdiscip. Top. Gerontol. 2014, 39, 45–61. [Google Scholar] [CrossRef]
- Song, X.; Bao, M.; Li, D.; Li, Y.M. Advanced glycation in D-galactose induced mouse aging model. Mech. Ageing Dev. 1999, 108, 239–251. [Google Scholar] [CrossRef]
- Chiu, P.Y.; Lam, P.Y.; Yan, C.W.; Ko, K.M. Schisandrin B protects against solar irradiation-induced oxidative injury in BJ human fibroblasts. Fitoterapia 2011, 82, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Lam, P.Y.; Yan, C.W.; Chiu, P.Y.; Leung, H.Y.; Ko, K.M. Schisandrin B protects against solar irradiation-induced oxidative stress in rat skin tissue. Fitoterapia 2011, 82, 393–400. [Google Scholar] [CrossRef]
- Ding, M.; Shu, P.; Gao, S.; Wang, F.; Gao, Y.; Chen, Y.; Deng, W.; He, G.; Hu, Z.; Li, T. Schisandrin B protects human keratinocyte-derived HaCaT cells from tert-butyl hydroperoxide-induced oxidative damage through activating the Nrf2 signaling pathway. Int. J. Mol. Med. 2018, 42, 3571–3581. [Google Scholar] [CrossRef]
- Marks, R. Successful aging and chronic osteoarthritis. Medicines 2018, 5, 105. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.W.; Lee, H.H.; Choi, E.O.; Lee, K.W.; Kim, K.Y.; Kim, S.G.; Hong, S.H.; Kim, G.Y.; Park, C.; Kim, H.K.; et al. Schisandrae Fructus inhibits IL-1β-induced matrix metalloproteinases and inflammatory mediators production in SW1353 human chondrocytes by suppressing NF-κB and MAPK activation. Drug Dev. Res. 2015, 76, 474–483. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Aran, L.; Bulli, G.; Curcio, F.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Sarcopenia: Assessment of disease burden and strategies to improve outcomes. Clin. Interv. Aging 2018, 13, 913–927. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yoo, S.R.; Chae, C.K.; Jung, U.J.; Choi, M.S. Omija fruit extract improves endurance and energy metabolism by upregulating PGC-1 α expression in the skeletal muscle of exercised rats. J. Med. Food 2014, 17, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Han, M.H.; Kim, G.Y.; Kim, C.M.; Chung, H.Y.; Hwang, H.J.; Kim, B.W.; Choi, Y.H. Schisandrae semen essential oil attenuates oxidative stress-induced cell damage in C2C12 murine skeletal muscle cells through Nrf2-mediated upregulation of HO-1. Int. J. Mol. Med. 2015, 35, 453–459. [Google Scholar] [CrossRef]
- Kim, J.W.; Ku, S.K.; Han, M.H.; Kim, K.Y.; Kim, S.G.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Kim, C.M.; Choi, Y.H. The administration of Fructus Schisandrae attenuates dexamethasone-induced muscle atrophy in mice. Int. J. Mol. Med. 2015, 36, 29–42. [Google Scholar] [CrossRef]
- Kim, J.W.; Ku, S.K.; Kim, K.Y.; Kim, S.G.; Han, M.H.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Kim, C.M.; Choi, Y.H. Schisandrae Fructus supplementation ameliorates sciatic neurectomy-induced muscle atrophy in mice. Oxid. Med. Cell. Longev. 2015, 2015, 872428. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Shin, J.H.; Hwang, S.J.; Choi, Y.H.; Kim, D.S.; Kim, C.M. Schisandrae Fructus enhances myogenic differentiation and inhibits atrophy through protein synthesis in human myotubes. Int. J. Nanomed. 2016, 11, 2407–2415. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Han, M.H.; Kim, G.Y.; Kim, C.M.; Kim, B.W.; Hwang, H.J.; Hyun, Y. Nrf2-mediated HO-1 induction contributes to antioxidant capacity of a Schisandrae Fructus ethanol extract in C2C12 myoblasts. Nutrients 2014, 6, 5667–5578. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Ku, S.K.; Lee, K.W.; Song, C.H.; An, W.G. Muscle-protective effects of Schisandrae Fructus extracts in old mice after chronic forced exercise. J. Ethnopharmacol. 2018, 212, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Jeong, S.H.; Han, S.H.; Yi, H.K. Gomisin A Modulates aging progress via mitochondrial biogenesis in human diploid fibroblast cells. Clin. Exp. Pharmacol. Physiol. 2018, 45, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.M.; Chen, N.; Leung, H.Y.; Leong, E.P.; Poon, M.K.; Chiu, P.Y. Long-term schisandrin B treatment mitigates age-related impairments in mitochondrial antioxidant status and functional ability in various tissues, and improves the survival of aging C57BL/6J mice. BioFactors 2008, 34, 331–342. [Google Scholar] [CrossRef]
- Meng, Q.; Wong, Y.T.; Chen, J.; Ruan, R. Age-related changes in mitochondrial function and antioxidative enzyme activity in Fischer 344 rats. Mech. Ageing Dev. 2007, 128, 286–292. [Google Scholar] [CrossRef]
- Sun, J.; Jing, S.; Jiang, R.; Wang, C.; Zhang, C.; Chen, J.; Li, H. Metabolomics study of the therapeutic mechanism of Schisandra chinensis lignans on aging rats induced by d-galactose. Clin. Interv. Aging 2018, 13, 829–841. [Google Scholar] [CrossRef]
- Poddar, J.; Pradhan, M.; Ganguly, G.; Chakrabarti, S. Biochemical deficits and cognitive decline in brain aging: Intervention by dietary supplements. J. Chem. Neuroanat. 2019, 95, 70–80. [Google Scholar] [CrossRef]
- Yan, T.; Shang, L.; Wang, M.; Zhang, C.; Zhao, X.; Bi, K.; Jia, Y. Lignans from Schisandra chinensis ameliorate cognition deficits and attenuate brain oxidative damage induced by d-galactose in rats. Metab. Brain Dis. 2016, 31, 653–661. [Google Scholar] [CrossRef]
- World Health Organisation. Global Report on Diabetes. 2016. Available online: http://apps.who.int/iris/bitstream/handle/10665/204871/9789241565257_eng.pdf;jsessionid=3862A156C0FCBE418F52FFD23635E893?sequence=1 (accessed on 26 November 2018).
- World Health Organisation. Obesity and Overweight—Key Facts. 2018. Available online: http://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 26 November 2018).
- World Health Organisation. Cancer—Key Facts. 2018. Available online: http://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 26 November 2018).
- Xue, Y.; Li, X.; Du, X.; Li, X.; Wang, W.; Yang, J.; Chen, J.; Pu, J.; Sun, H. Isolation and anti-hepatitis B virus activity of dibenzocyclooctadiene lignans from the fruits of Schisandra chinensis. Phytochemistry 2015, 116, 253–261. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Deng, L.L.; Liu, J.J.; Zhang, J.X.; Hao, X.J.; Mu, S.Z. Schisanhenol derivatives and their biological evaluation against tobacco mosaic virus (TMV). Fitoterapia 2015, 101, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.; Wu, Y.H.; Tseng, C.K.; Lin, C.K.; Hsu, Y.C.; Chen, Y.H.; Lee, J.C. Schisandrin A inhibits dengue viral replication via upregulating antiviral interferon responses through STAT signaling pathway. Sci. Rep. 2017, 7, 45171. [Google Scholar] [CrossRef] [PubMed]
- Song, M.Y.; Wang, J.H.; Eom, T.; Kim, H. Schisandra chinensis fruit modulates the gut microbiota composition in association with metabolic markers in obese women: A randomized, double-blind placebo-controlled study. Nutr. Res. 2015, 35, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Ozsurekci, Y.; Aykac, K. Oxidative stress related diseases in newborns. Oxid. Med. Cell. Longev. 2016, 2016, 2768365. [Google Scholar] [CrossRef] [PubMed]
- Leong, P.K.; Wong, H.S.; Chen, J.; Chan, W.M.; Leung, H.Y.; Ko, K.M. Differential action between Schisandrin A and Schisandrin B in eliciting an anti-inflammatory action: The depletion of reduced glutathione and the induction of an antioxidant response. PLoS ONE 2016, 11, e0155879. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H. Schisandrin A prevents oxidative stress-induced DNA damage and apoptosis by attenuating ROS generation in C2C12 cells. Biomed. Pharmacother. 2018, 106, 902–909. [Google Scholar] [CrossRef]
- Valle, I.; Alvarez-Barrientos, A.; Arza, E.; Lamas, S.; Monsalve, M. PGC-1alpha regulates the mitochondrial antioxidant defense system in vascular endothelial cells. Cardiovasc. Res. 2005, 66, 562–573. [Google Scholar] [CrossRef]
- Iacovelli, J.; Rowe, G.C.; Khadka, A.; Diaz-Aguilar, D.; Spencer, C.; Arany, Z.; Saint-Geniez, M. PGC-1alpha induces human RPE oxidative metabolism and antioxidant capacity. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1038–1051. [Google Scholar] [CrossRef]
- Konishi, T. Brain oxidative stress as basic target of antioxidant traditional oriental medicines. Neurochem. Res. 2009, 34, 711–716. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| SCE constituent/extract | In vitro effects | Reference |
|---|---|---|
| Schisandrin B | Inhibition of triple negative breast cancer growth by inducing cell cycle arrest/apoptosis. | [52] |
| Schisandrin B | Cell cycle arrest at G0/G1, apoptosis, inhibition of invasion, and migration of A549 cells. | [51] |
| Sesquiterpenes from fruits (ethanol extract) | Cytotoxicity in Caco2 cells. | [50] |
| Dibenzocyclooctadiene lignans (+)-deoxyschisandrin (1) and (−)-gomisin N (2), from S. chinensis fruits | Inhibition of adenocarcinoma cells (ovarian 2008 and colon LoVo) growth by apoptosis correlated with G2/M arrest and tubulin polymerisation. | [55] |
| Polysaccharides | Apoptosis of renal cell carcinoma Caki-1 by blocking secretion of endothelial growth factor. | [62] |
| Polysaccharide-0-1 | Inhibition of human hepatocellular liver carcinoma (HepG2) cells growth via G2/M arrest/apoptosis. | [56] |
| Polysaccharides | Mitochondria-mediated apoptosis in human renal carcinoma cell line (Caki-1) through the inactivation of the extracellular-signal-regulated kinase (ERK) pathway. | [63] |
| Semen essential oil | Apoptosis of human leukaemia U937 cells by ROS- and caspase-dependent mitochondrial dysfunction and nuclear translocation of mitochondrial pro-apoptotic proteins. | [64] |
| Gomisin G | Suppression of proliferation and cell cycle arrest in G1 in triple-negative breast cancer cells. | [65] |
| Gomisin A | Decreased viability of various colorectal cancer cell lines; G0/G1 arrest and apoptosis of CT26 and HT29 cells by regulating cyclin D1/cyclin-dependent kinase 4 (CDK4) expression and apoptotic proteins; reduced invasion and migration of cancer cells. | [54] |
| Deoxyschisandrin from fruits (ethanol extract) | G0/G1 arrest in human ovarian cancer cells (A2780, SKOV3, and OVCAR3); reduced protumoural phenotype of tumour-associated macrophages (TAMs). | [57] |
| Deoxyschisandrin Schisandrin B | Protection against UVB-induced DNA damage and apoptosis in HaCaT cells via ROS scavenging. | [66] |
| Fruit water extract | Inhibited proliferation, depolarised mitochondrial membrane and apoptosis by ROS generation and activation of caspase-9 and -3 in AGS human gastric cancer cells. | [67] |
| Terpenoids from the n-hexane fraction of S. chinensis extract | Cytotoxicity in human leukemic HL-60 cells. | [68] |
| Nineteen compounds isolated from hexane and ethyl acetate soluble fractions from fruits (ethanol extract) | Cytotoxicity in human ovarian (A2780) and endometrial cancer (Ishikawa) cells. | [69] |
| Kadsuphilactone B | Apoptosis via the Bcl-2 (B-cell lymphoma 2) and MAPK (mitogen activated protein kinase) signalling pathways. | |
| In vivo effects | ||
| Schisandrin B | Inhibition of migration and colony formation of tumour cells, prevention of growth of TNBC cells in mice via suppression of signal transducer and activator of transcription-3 (STAT3) phosphorylation and nuclear translocation. | [52] |
| Polysaccharides | Antitumor and antiangiogenic activity associated with the up-regulation of Bax and p53, downregulation of Bcl-2, as well as the reduction of CD31 and CD34 in tumours in BALB/c nude mice. | [62] |
| Gomisin A | Inhibition of metastasis of colorectal cancer in a lung metastasis mouse model. | [54] |
| Seed extract | Apoptosis in human osteosarcoma cells (1547). | [70] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak, A.; Zakłos-Szyda, M.; Błasiak, J.; Nowak, A.; Zhang, Z.; Zhang, B. Potential of Schisandra chinensis (Turcz.) Baill. in Human Health and Nutrition: A Review of Current Knowledge and Therapeutic Perspectives. Nutrients 2019, 11, 333. https://doi.org/10.3390/nu11020333
Nowak A, Zakłos-Szyda M, Błasiak J, Nowak A, Zhang Z, Zhang B. Potential of Schisandra chinensis (Turcz.) Baill. in Human Health and Nutrition: A Review of Current Knowledge and Therapeutic Perspectives. Nutrients. 2019; 11(2):333. https://doi.org/10.3390/nu11020333
Chicago/Turabian StyleNowak, Adriana, Małgorzata Zakłos-Szyda, Janusz Błasiak, Agnieszka Nowak, Zhuo Zhang, and Bolin Zhang. 2019. "Potential of Schisandra chinensis (Turcz.) Baill. in Human Health and Nutrition: A Review of Current Knowledge and Therapeutic Perspectives" Nutrients 11, no. 2: 333. https://doi.org/10.3390/nu11020333
APA StyleNowak, A., Zakłos-Szyda, M., Błasiak, J., Nowak, A., Zhang, Z., & Zhang, B. (2019). Potential of Schisandra chinensis (Turcz.) Baill. in Human Health and Nutrition: A Review of Current Knowledge and Therapeutic Perspectives. Nutrients, 11(2), 333. https://doi.org/10.3390/nu11020333