Can Carob-Fruit-Extract-Enriched Meat Improve the Lipoprotein Profile, VLDL-Oxidation, and LDL Receptor Levels Induced by an Atherogenic Diet in STZ-NAD-Diabetic Rats?

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Carob Fruit Extract (CFE)
2.2. Restructured Meat and Diet Preparation
2.3. Experimental Design
2.4. Growth Rate and Fecal Fat Extraction
2.5. Glycemia
2.6. Lipoprotein Isolation
2.7. Plasma Lipid Analysis and Lipoprotein Composition
2.8. Arylesterase Activity Measurement
2.9. VLDL and Liver Oxidation (TBARS Assay)
2.10. LDL-Receptor Levels by Western Blotting and Immunohistochemistry
2.11. Statistical Analyses
3. Results
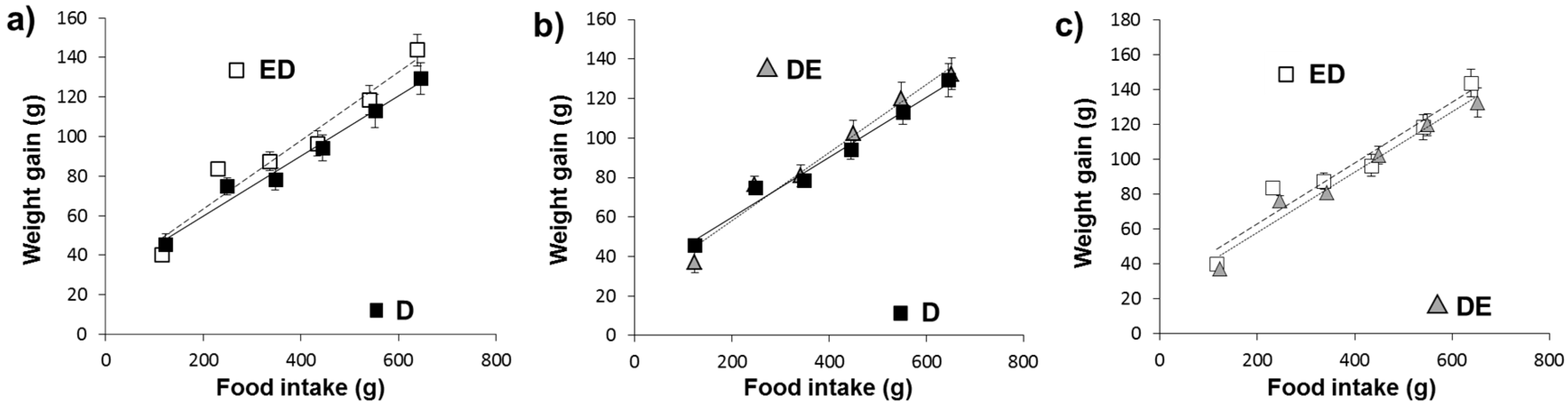
3.1. Growth Rate, Feed Consumption and Fecal Excretion
3.2. Plasma Glucose and Lipid Concentrations
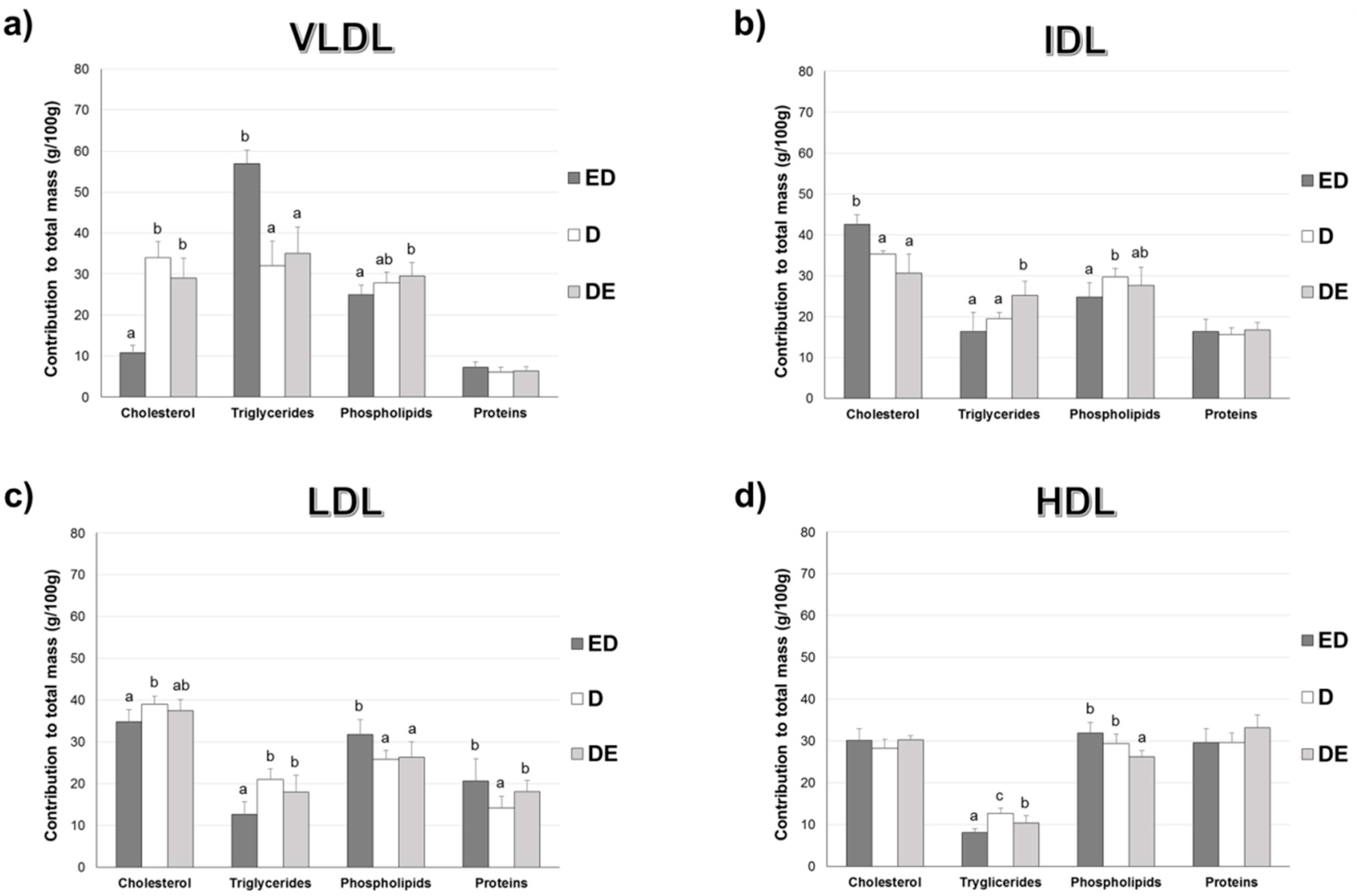
3.3. Lipoprotein Composition
3.4. Percentage Contribution of Lipids to Lipoprotein Composition
3.5. Arylesterase Activity, VLDL and Liver Oxidation
3.6. Liver Macroscopic Aspect
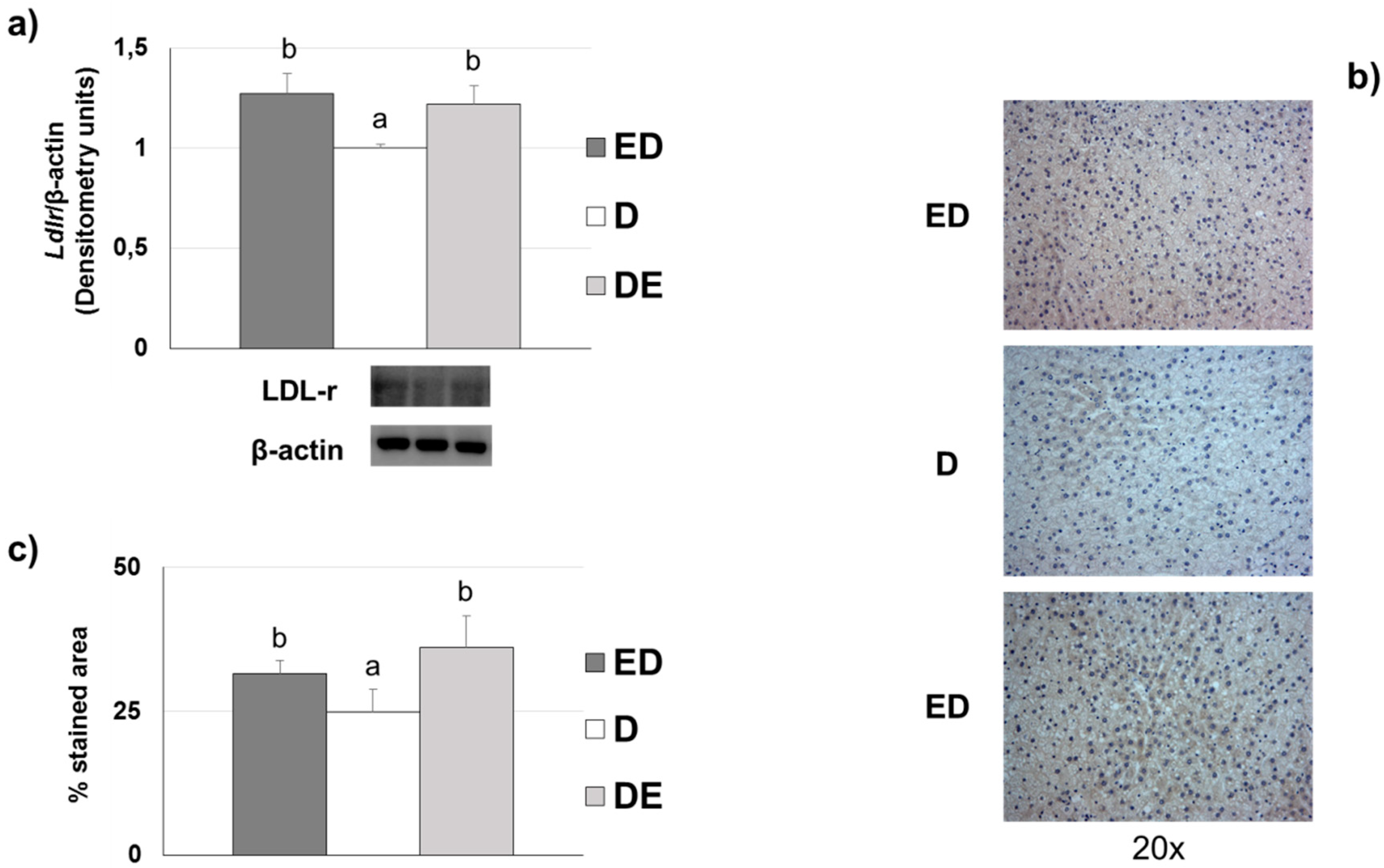
3.7. Liver LDL Receptor Levels
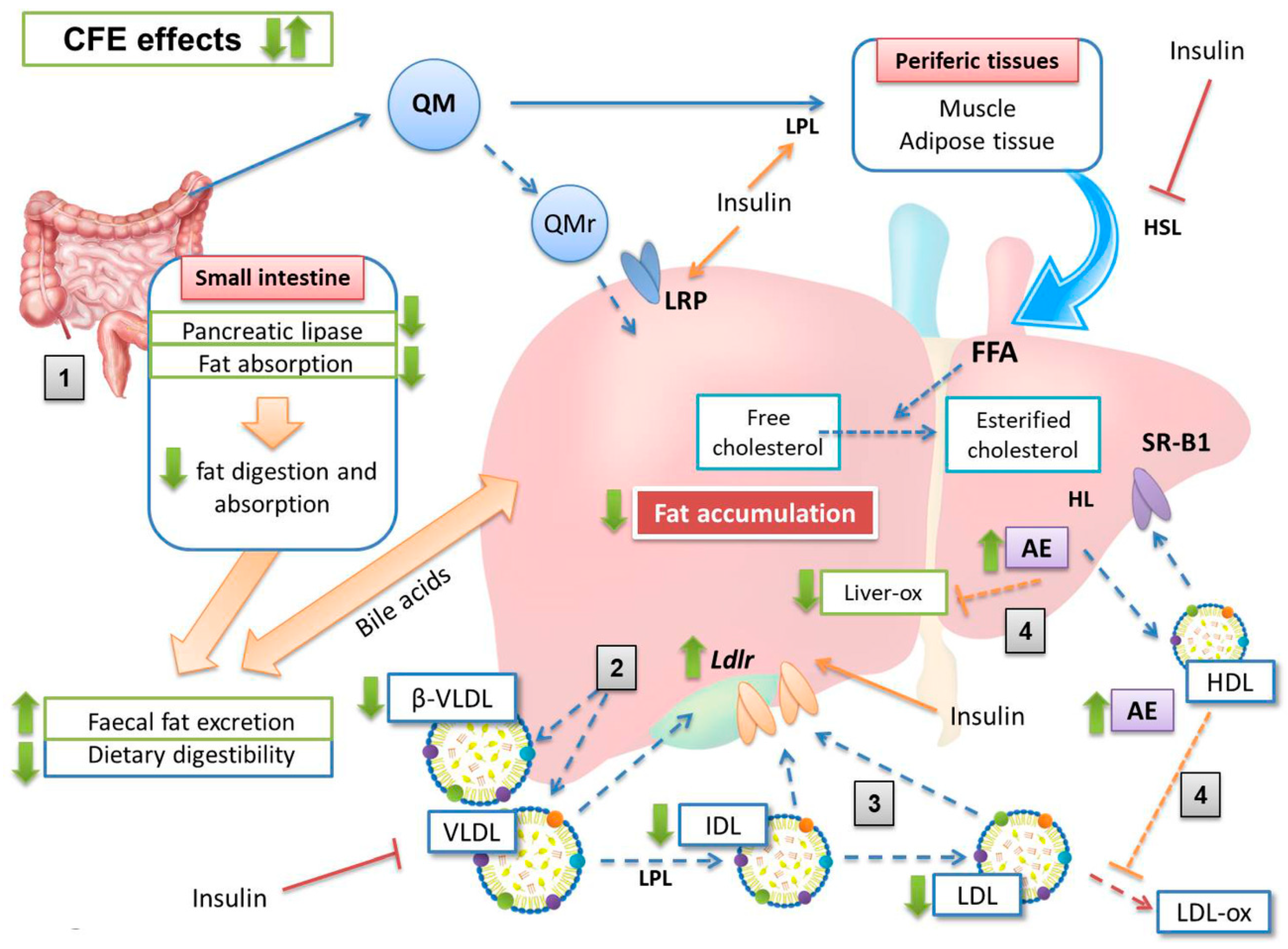
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Jiménez-Colmenero, F.; Carballo, J.; Cofrades, S. Healthier meat and meat products: Their role as functional foods. Meat Sci. 2001, 59, 5–13. [Google Scholar] [CrossRef]
- Tian, S.; Xu, Q.; Jiang, R.; Han, T.; Sun, C.; Na, L. Dietary protein consumption and the risk of type 2 diabetes: A systematic review and meta-analysis of cohort studies. Nutrients 2017, 9, 982. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Hoffmann, G.; Lampousi, A.M.; Knuppel, S.; Iqbal, K.; Schwedhelm, C.; Bechthold, A.; Schlesinger, S.; Boeing, H. Food groups and risk of type 2 diabetes mellitus: A systematic review and meta-analysis of prospective studies. Eur. J. Epidemiol. 2017, 32, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.; Jones, S.A.; Nuttall, F.Q.; Gannon, M.C. A fasting-induced decrease in plasma glucose concentration does not affect the insulin response to ingested protein in people with type 2 diabetes. Metabolism 2002, 51, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.J.C.; Kruijshoop, M.; Menheere, P.; Wagenmakers, A.J.M.; Saris, W.H.M.; Keizer, H.A. Amino acid ingestion strongly enhances insulin secretion in patients with long-term type 2 diabetes. Diabetes Care 2003, 26, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Verges, B. Pathophysiology of diabetic dyslipidaemia: Where are we? Diabetologia 2015, 58, 886–899. [Google Scholar] [CrossRef] [PubMed]
- Sugden, M.; Holness, M. Pathophysiology of diabetic dyslipidemia: Implications for atherogenesis and treatment. Clin. Lipidol. 2011, 6, 401–411. [Google Scholar]
- Mbue, N.D.; Mbue, J.E.; Anderson, J.A. Management of lipids in patients with diabetes. Nur. Clin. N. Am. 2017, 52, 605–619. [Google Scholar] [CrossRef]
- Garcimartín, A.; Santos-López, J.A.; Bastida, S.; Benedi, J.; Sánchez-Muniz, F.J. Silicon-enriched restructured pork affects the lipoprotein profile, vldl oxidation, and ldl receptor gene expression in aged rats fed an atherogenic diet. J. Nutr. 2015, 145, 2039–2045. [Google Scholar] [CrossRef]
- Moreira, A.R.S.; Olivero-David, R.; Vázquez-Velasco, M.; González-Torres, L.; Benedi, J.; Bastida, S.; Sánchez-Muniz, F.J. Protective Effects of sea spaghetti-enriched restructured pork against dietary cholesterol: Effects on arylesterase and lipoprotein profile and composition of growing rats. J. Med. Food. 2014, 17, 921–928. [Google Scholar] [CrossRef]
- Olivero-David, R.; Schultz-Moreira, A.; Vázquez-Velasco, M.; González-Torres, L.; Bastida, S.; Benedi, J.; Sánchez-Reus, M.I.; González-Muñoz, M.J.; Sánchez-Muniz, F.J. Effects of nori- and wakame-enriched meats with or without supplementary cholesterol on arylesterase activity, lipaemia and lipoproteinaemia in growing Wistar rats. Br. J. Nutr. 2011, 106, 1476–1486. [Google Scholar] [CrossRef] [PubMed]
- Viejo, J.; García-Linares, M.C.; Bastida, S.; García-Arias, M.T.; Sánchez-Muniz, F.J. Effect of olive oil-fried sardine consumption on liver lipid composition and fatty acid cholesterol esterification in hypercholesterolemic rats. Food Sci. Technol. Int. 2003, 9, 329–338. [Google Scholar] [CrossRef]
- Sántos-Lopez, J.A.; Garcimartín, A.; Bastida, S.; Bautista-Ávila, M.; González-Muñoz, M.J.; Benedi, J.; Sánchez-Muniz, F.J. Lipoprotein profile in aged rats fed chia oil- or hydroxytyrosol-enriched pork in high cholesterol/high saturated fat diets. Nutrients 2018, 10, 1830. [Google Scholar] [CrossRef] [PubMed]
- Van den Driessche, J.J.; Plat, J.; Mensink, R.P. Effects of superfoods on risk factors of metabolic syndrome: A systematic review of human intervention trials. Food Funct. 2018, 9, 1944–1966. [Google Scholar] [CrossRef] [PubMed]
- Asgary, S.; Rastqar, A.; Keshvari, M. Functional food and cardiovascular disease prevention and treatment: A review. J. Am. Coll. Nutr. 2018, 37, 429–455. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, A.; Tsang, C.; Tiss, A.; Bahorun, T.; Arefanian, H.; Barake, R.; Khadir, A.; Tuomilehto, J. Functional foods and lifestyle approaches for diabetes prevention and management. Nutrients 2017, 9, 1310. [Google Scholar] [CrossRef] [PubMed]
- Ballali, S.; Lanciai, F. Functional food and diabetes: A natural way in diabetes prevention? Int. J. Food Sci. Nutr. 2012, 63, 51–61. [Google Scholar] [CrossRef]
- Mirmiran, P.; Bahadoran, Z.; Azizi, F. Functional foods-based diet as a novel dietary approach for management of type 2 diabetes and its complications: A. review. World J. Diabetes 2014, 5, 267–281. [Google Scholar] [CrossRef]
- Bahadoran, Z.; Mirmiran, P.; Azizi, F. Dietary polyphenols as potential nutraceuticals in management of diabetes: A review. J. Diabetes Metab. Disord. 2013, 12, 43. [Google Scholar] [CrossRef]
- Delgado-Pando, G.; Celada, P.; Sánchez-Muniz, F.J.; Jiménez-Colmenero, F.; Olmedilla-Alonso, B. Effects of improved fat content of frankfurters and pates on lipid and lipoprotein profile of volunteers at increased cardiovascular risk: A placebo-controlled study. Eur. J. Nutr. 2014, 53, 83–93. [Google Scholar] [CrossRef]
- Macho-González, A.; Garcimartín, A.; López-Oliva, M.E.; Bertocco, G.; Naes, F.; Bastida, S.; Sánchez-Muniz, F.J.; Benedi, J. Fiber purified extracts of carob fruit decrease carbohydrate absorption. Food Funct. 2017, 8, 2258–2265. [Google Scholar] [CrossRef] [PubMed]
- Macho-González, A.; Garcimartín, A.; Naes, F.; López-Oliva, M.E.; Amores-Arrojo, A.; González-Muñoz, M.J.; Bastida, S.; Benedi, J.; Sánchez-Muniz, F.J. Effects of fiber purified extract of carob fruit on fat digestion and postprandial lipemia in healthy rats. J. Agric. Food Chem. 2018, 66, 6734–6741. [Google Scholar] [CrossRef] [PubMed]
- Bastida, S.; Sánchez-Muniz, F.J.; Olivero, R.; Pérez-Olleros, L.; Ruiz-Roso, B.; Jiménez-Colmenero, F. Antioxidant activity of Carob fruit extracts in cooked pork meat systems during chilled and frozen storage. Food Chem. 2009, 116, 748–754. [Google Scholar] [CrossRef]
- Zulim Botega, D.; Bastida, S.; Marmesat Rodas, S.; Pérez-Olleros, L.; Ruiz-Roso, B.; Sánchez-Muniz, F. Carob fruit polyphenols reduce tocopherol loss, triacylglycerol polymerization and oxidation in heated sunflower oil. J. Am. Oil. Chem. Soc. 2009, 86, 419–425. [Google Scholar] [CrossRef]
- Gheibi, S.; Kashfi, K.; Ghasemi, A. A practical guide for induction of type-2 diabetes in rat: Incorporating a high-fat diet and streptozotocin. Biomed. Pharmacother. 2017, 95, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Tyurenkov, I.N.; Kurkin, D.V.; Bakulin, D.A.; Volotova, E.V.; Chafeev, M.A.; Smirnov, A.V.; Morkovin, E.I. ZB-16, a Novel GPR119 agonist, relieves the severity of streptozotocin-nicotinamide-induced diabetes in rats. Front. Endocrinol. 2017, 8, 152. [Google Scholar] [CrossRef] [PubMed]
- De Mora, B.R.R.C.; Marcos, A.M.R.; Conde, L.P.O.; Casero, M.M.; Haber, B. Denatured Carob Flour (DCF) with a Low Content of Soluble Tannins and Sugars, Meant for Human Consumption and Process to Obtain It. U.S. Patent 7,393,551, 1 July 2008. WO2004/014150. [Google Scholar]
- Salvadó, M.J.; Casanova, E.; Fernandez-Iglesias, A.; Arola, L.; Blade, C. Roles of proanthocyanidin rich extracts in obesity. Food Funct. 2015, 6, 1053–1071. [Google Scholar] [CrossRef]
- De Mora, B.R.R.C.; Marcos, A.M.R.; Conde, L.P.O.; Hueso, J.A.H. Product of Vegetal Origin Comprising Proanthocyanidines and its Preparation Process. U.S. Patent 8,663,708, 4 March 2014. [Google Scholar]
- Moreira, A.S.; González-Torres, L.; Olivero-David, R.; Bastida, S.; Benedi, J.; Sánchez-Muniz, F.J. Wakame and nori in restructured meats included in cholesterol-enriched diets affect the antioxidant enzyme gene expressions and activities in wistar rats. Plant Foods Hum. Nutr. 2010, 65, 290–298. [Google Scholar] [CrossRef]
- Terpstra, A.H.M.; Woodward, C.J.H.; Sánchez-Muniz, F.J. Improved techniques for the separation of serum-lipoproteins by density gradient ultra-centrifugation–visualization by pre-staining and rapid separation of serum-lipoproteins from small volumes of serum. Anal. Biochem. 1981, 111, 149–157. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nus, M.; Sánchez-Muniz, F.J.; Gago, J.V.S.; Lopez-Oliva, E.; Sanchez-Montero, M. Determination of rat and mice arylesterase activity using serum mimetics. Enzyme Microb. Technol. 2008, 43, 252–256. [Google Scholar] [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Sánchez-Muniz, F.J.; Bastida, S. Do not use the Friedewald formula to calculate LDL-cholesterol in hypercholesterolaemic rats. Eur. J. Lipid Sci. Technol. 2008, 110, 295–301. [Google Scholar] [CrossRef]
- Bocanegra, A.; Benedi, J.; Sánchez-Muniz, F.J. Differential effects of konbu and nori seaweed dietary supplementation on liver glutathione status in normo- and hypercholesterolaemic growing rats. Br. J. Nutr. 2006, 95, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Muniz, F.J. Dietary fibre and cardiovascular health. Nutr. Hosp. 2012, 27, 31–45. [Google Scholar] [PubMed]
- Ruiz-Roso, B.; Quintela, J.C.; de la Fuente, E.; Haya, J.; Pérez-Olleros, L. insoluble carob fiber rich in polyphenols lowers total and LDL cholesterol in hypercholesterolemic sujects. Plant Foods Hum. Nutr. 2010, 65, 50–56. [Google Scholar] [CrossRef]
- Katsiki, N.; Tentolouris, N.; Mikhailidis, D.P. Dyslipidaemia in type 2 diabetes mellitus: Bad for the heart. Curr. Opin. Cardiol. 2017, 32, 422–429. [Google Scholar] [CrossRef]
- Carmena, R. Dyslipidemia in Type 2 Diabetes Mellitus. In Type 2 Diabetes Mellitus; Serrano Ríos, N., Gutiérrez Fuentes, J.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 219–230. [Google Scholar]
- Mahley, R.W. Atherogenic hyperlipoproteinemia. The cellular and molecular biology of plasma lipoproteins altered by dietary fat and cholesterol. Med. Clin. N. Am. 1982, 66, 375–402. [Google Scholar] [CrossRef]
- Sánchez-Muniz, F.J.; Bastida, S.; Viejo, J.M.; Terpstra, A.H. Small supplements of N-3 fatty acids change serum low density lipoprotein composition by decreasing phospholid and apolipoprotein B concentrations in young adult women. Eur. J. Nutr. 1999, 38, 20–27. [Google Scholar] [CrossRef]
- Grundy, S.M.; Denke, M.A. Dietary influences on serum lipids and lipoproteins. J. Lipid Res. 1990, 31, 1149–1172. [Google Scholar] [PubMed]
- Hazarika, A.; Kalita, H.; Kalita, M.C.; Devi, R. Withdrawal from high-carbohydrate, high-saturated-fat diet changes saturated fat distribution and improves hepatic low-density-lipoprotein receptor expression to ameliorate metabolic syndrome in rats. Nutrition 2017, 38, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Tolman, K.G.; Fonseca, V.; Dalpiaz, A.; Tan, M.H. Spectrum of liver disease in type 2 diabetes and management of patients with diabetes and liver disease. Diabetes Care 2007, 30, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Dietschy, J.M. Dietary fatty acids and the regulation of plasma low density lipoprotein cholesterol concentrations. J. Nutr. 1998, 128, 444s–448s. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Zhang, Z.; Dai, X.; Jiang, Y.; Bao, L.; Li, Y.; Li, Y. Grape seed proanthocyanidins ameliorate pancreatic beta-cell dysfunction and death in low-dose streptozotocin- and high-carbohydrate/high-fat diet-induced diabetic rats partially by regulating endoplasmic reticulum stress. Nutr. Metab. 2013, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, Z.; Silver, D.L.; Jiang, X.C. Cholesteryl ester transfer protein (CETP) expression enhances HDL cholesteryl ester liver delivery, which is independent of scavenger receptor BI, LDL receptor related protein and possibly LDL receptor. Biochim. Biophys. Acta 2006, 1761, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Mohamadin, A.M.; Elberry, A.A.; Abdel Gawad, H.S.; Morsy, G.M.; Al-Abbasi, F.A. Protective effects of simvastatin, a lipid lowering agent, against oxidative damage in experimental diabetic rats. J. Lipids 2011, 2011, 167958. [Google Scholar] [CrossRef]
- Zimmermann, A.M.; Rivero, A.C.; Bezerra, A.S.; Ruviaro, A.R.; Novack, M.M.E.; Nornberg, J.L. Antioxidant potential of barley extract in rats subjected to a high-fat diet. Food Sci. Technol. 2013, 33, 167–171. [Google Scholar] [CrossRef]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharmacol. 2017, 174, 1244–1262. [Google Scholar]
- Estrada-Luna, D.; Martinez-Hinojosa, E.; Cancino-Diaz, J.C.; Belefant-Miller, H.; Lopez-Rodriguez, G.; Betanzos-Cabrera, G. Daily supplementation with fresh pomegranate juice increases paraoxonase 1 expression and activity in mice fed a high-fat diet. Eur. J. Nutr. 2018, 57, 383–389. [Google Scholar] [CrossRef]
- Rock, W.; Rosenblat, M.; Miller-Lotan, R.; Levy, A.P.; Elias, M.; Aviram, M. Consumption of wonderful variety pomegranate juice and extract by diabetic patients increases paraoxonase 1 association with high-density lipoprotein and stimulates its catalytic activities. J. Agric. Food Chem. 2008, 56, 8704–8713. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.M.; Gu, L.J.; Xia, Y.R.; Navab, M.; Li, W.F.; Hama, S.; Castellani, L.W.; Furlong, C.E.; Costa, L.G.; Fogelman, A.M.; et al. Mice lacking serum paraoxonase are susceptible to organophosphate toxicity and atherosclerosis. Nature 1998, 394, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.; Durrington, P.; McElduff, P.; Yarnell, J.; Azam, N.; Watt, M.; Mackness, M. Low paraoxonase activity predicts coronary events in the caerphilly prospective study. Circulation 2003, 107, 2775–2779. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Components | Chol-Diet | CFE-Diet |
|---|---|---|
| Protein, % | 17.03 | 17.03 |
| Fat, % | 27.02 | 27.02 |
| Cholesterol, g/kg | 9.83 | 9.83 |
| SFA:MUFA:PUFA ratio | 2.76/2.99/1 | 2.76/2.99/1 |
| Energy content *, MJ/kg | 20.36 | 20.36 |
| Ingredients, g/kg | ||
| Sucrose | 68.25 | 68.25 |
| Corn starch | 275.73 | 275.73 |
| Casein | 94.25 | 94.25 |
| Maltodextrin | 94.25 | 94.25 |
| Cellulose | 48.86 | 48.86 |
| PM 205B SAFE | 50.05 | 50.05 |
| PV 200 SAFE | 7.15 | 7.15 |
| Soybean oil | 47.91 | 47.91 |
| L-Cysteine | 2.02 | 2.02 |
| Cholesterol | 9.1 | 9.1 |
| Cholic acid | 1.3 | 1.3 |
| Freeze-dried restructured meat | 301.14 | 301.14 |
| ANOVA | ||||
|---|---|---|---|---|
| ED | D | DE | p | |
| Cholesterol intake (g/wk) | 1.07 ± 0.04 | 1.08 ± 0.08 | 1.09 ± 0.08 | p > 0.05 |
| Growth rate * | 0.18 ± 0.03 b | 0.15 ± 0.01 a | 0.18 ± 0.01 b | p = 0.019 |
| Final weight (g) | 380.1 ± 39.3 | 366.6 ± 32.3 | 380.4 ± 22.3 | p > 0.05 |
| Fecal excretion (g/wk) ** | 14.10 ± 1.43 b | 11.16 ± 0.43 a | 13.57 ± 1.69 b | p = 0.001 |
| Fecal moisture (%) | 18.56 ± 1.55 b | 14.04 ± 1.66 a | 16.62 ± 1.40 b | p < 0.001 |
| Fecal fat (mg/g feces) ** | 226.6 ± 26.93 b | 170.6 ± 10.07 a | 236.5 ± 25.20 b | p < 0.001 |
| Dietary digestibility *** | 0.85 ± 0.01 a | 0.88 ± 0.01 b | 0.86 ± 0.02 ab | p = 0.004 |
| ANOVA | ||||
|---|---|---|---|---|
| ED | D | DE | p | |
| Glycemia 3rd week (mmol/L) | 8.02 ± 0.57 | 8.04 ± 0.96 | 8.27 ± 0.70 | p > 0.05 |
| Glycemia 8th week (mmol/L) | 15.76 ± 0.76 b | 18.11 ± 1.65 c | 13.94 ± 1.13 a | p < 0.001 |
| Total cholesterol (mmol/L) | 2.26 ± 0.12 a | 2.95 ± 0.22 c | 2.57 ± 0.16 b | p < 0.001 |
| Triglycerides (mmol/L) | 0.74 ± 0.05 a | 0.87 ± 0.11 b | 0.71 ± 0.13 ab | p = 0.013 |
| Phospholipids (mmol/L) | 1.14 ± 0.13 | 1.28 ± 0.09 | 1.26 ± 0.17 | p > 0.05 |
| Total lipids (mg/dL) * | 235 ± 14.1 a | 282 ± 29.8 b | 252 ± 24.6 ab | p = 0.003 |
| Cholesterol:phospholipids (mol/mol) | 1.01 ± 0.09 | 1.13 ± 0.10 | 1.02 ± 0.16 | p > 0.05 |
| Cholesterol:HDL cholesterol (mol/mol) | 1.65 ± 0.29 a | 2.37 ± 0.45 b | 2.10 ± 0.18 b | p < 0.001 |
| Atherogenic index ** | 0.65 ± 0.29 a | 1.37 ± 0.45 b | 1.10 ± 0.18 b | p < 0.001 |
| ANOVA | ||||
|---|---|---|---|---|
| Lipoprotein | ED | D | DE | p |
| Cholesterol, mmol/L | ||||
| VLDL | 0.22 ± 0.03 a | 0.78 ± 0.09 c | 0.62 ± 0.12 b | p < 0.001 |
| IDL | 0.44 ± 0.08 a | 0.55 ± 0.03 b | 0.41 ± 0.05 a | p < 0.001 |
| LDL | 0.16 ± 0.03 a | 0.34 ± 0.06 c | 0.24 ± 0.01 b | p < 0.001 |
| HDL | 1.40 ± 0.16 | 1.28 ± 0.19 | 1.22 ± 0.08 | p > 0.05 |
| Triglycerides, mmol/L | ||||
| VLDL | 0.48 ± 0.07 b | 0.30 ± 0.07 a | 0.32 ± 0.08 a | p < 0.001 |
| IDL | 0.07 ± 0.02 a | 0.13 ± 0.02 b | 0.14 ± 0.05 b | p < 0.001 |
| LDL | 0.02 ± 0.01 a | 0.07 ± 0.02 c | 0.05 ± 0.01 b | p < 0.001 |
| HDL | 0.15 ± 0.01 a | 0.23 ± 0.02 b | 0.17 ± 0.03 a | p < 0.001 |
| Phospholipids, mmol/L | ||||
| VLDL | 0.25 ± 0.03 a | 0.31 ± 0.03 b | 0.31 ± 0.03 b | p = 0.001 |
| IDL | 0.12 ± 0.03 a | 0.23 ± 0.01 c | 0.18 ± 0.04 b | p < 0.001 |
| LDL | 0.07 ± 0.01 a | 0.11 ± 0.01 b | 0.08 ± 0.02 a | p < 0.001 |
| HDL | 0.72 ± 0.02 c | 0.64 ± 0.06 b | 0.52 ± 0.04 a | p < 0.001 |
| Total lipids *, mg/dL | ||||
| VLDL | 69.72 ± 7.67 | 79.50 ± 7.13 | 73.54 ± 8.67 | p > 0.05 |
| IDL | 31.32 ± 4.69 a | 48.41 ± 2.57 c | 41.35 ± 4.28 b | p < 0.001 |
| LDL | 13.57 ± 2.14 a | 27.70 ± 4.01 c | 19.24 ± 1.25 b | p < 0.001 |
| HDL | 119.54 ± 5.82 b | 115.94 ± 11.30 b | 99.47 ± 7.11 a | p < 0.001 |
| Proteins, mg/dL | ||||
| VLDL | 5.43 ± 0.83 | 5.15 ± 0.86 | 4.89 ± 0.52 | p > 0.05 |
| IDL | 6.31 ± 2.20 a | 8.93 ± 1.32 b | 8.26 ± 0.96 ab | p = 0.005 |
| LDL | 3.64 ± 1.34 | 4.51 ± 0.93 | 4.27 ± 0.76 | p > 0.05 |
| HDL | 50.8 ± 8.94 | 48.8 ± 5.80 | 49.3 ± 5.79 | p > 0.05 |
| Total mass **, mg/dL | ||||
| VLDL | 75.15 ± 6.33 a | 84.65 ± 6.36 b | 78.43 ± 8.62 ab | p = 0.045 |
| IDL | 37.63 ± 6.48 a | 57.34 ± 3.13 c | 49.61 ± 4.73 b | p < 0.001 |
| LDL | 17.21 ± 2.92 a | 32.22 ± 4.32 c | 23.51 ± 1.36 b | p < 0.001 |
| HDL | 170.3 ± 12.19 b | 164.7 ± 14.92 ab | 148.8 ± 7.54 a | p < 0.001 |
| ANOVA | ||||
|---|---|---|---|---|
| ED | D | DE | p | |
| Plasma AE (U/L) * | 237 ± 43.06 b | 163 ± 29.56 a | 368 ± 58.59 c | p < 0.001 |
| Plasma AE:cholesterol (U/mg) ** | 0.29 ± 0.05 b | 0.15 ± 0.02 a | 0.39 ± 0.05 c | p < 0.001 |
| Liver AE (U/g protein) | 7.70 ± 0.97 b | 5.36 ± 0.83 a | 10.93 ± 3.28 b | p < 0.001 |
| VLDL-ox (TBARS, mg MDA/L) | 2.31 ± 0.29 ab | 2.58 ± 0.16 b | 2.22 ± 0.23 a | p = 0.024 |
| Liver-ox (TBARS, mg MDA/mg protein) | 2.63 ± 0.58 a | 3.63 ± 0.25 b | 2.49 ± 0.33 a | p < 0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macho-González, A.; Garcimartín, A.; López-Oliva, M.E.; Ruiz-Roso, B.; Martín de la Torre, I.; Bastida, S.; Benedí, J.; Sánchez-Muniz, F.J. Can Carob-Fruit-Extract-Enriched Meat Improve the Lipoprotein Profile, VLDL-Oxidation, and LDL Receptor Levels Induced by an Atherogenic Diet in STZ-NAD-Diabetic Rats? Nutrients 2019, 11, 332. https://doi.org/10.3390/nu11020332
Macho-González A, Garcimartín A, López-Oliva ME, Ruiz-Roso B, Martín de la Torre I, Bastida S, Benedí J, Sánchez-Muniz FJ. Can Carob-Fruit-Extract-Enriched Meat Improve the Lipoprotein Profile, VLDL-Oxidation, and LDL Receptor Levels Induced by an Atherogenic Diet in STZ-NAD-Diabetic Rats? Nutrients. 2019; 11(2):332. https://doi.org/10.3390/nu11020332
Chicago/Turabian StyleMacho-González, Adrián, Alba Garcimartín, María Elvira López-Oliva, Baltasar Ruiz-Roso, Isabel Martín de la Torre, Sara Bastida, Juana Benedí, and Francisco José Sánchez-Muniz. 2019. "Can Carob-Fruit-Extract-Enriched Meat Improve the Lipoprotein Profile, VLDL-Oxidation, and LDL Receptor Levels Induced by an Atherogenic Diet in STZ-NAD-Diabetic Rats?" Nutrients 11, no. 2: 332. https://doi.org/10.3390/nu11020332
APA StyleMacho-González, A., Garcimartín, A., López-Oliva, M. E., Ruiz-Roso, B., Martín de la Torre, I., Bastida, S., Benedí, J., & Sánchez-Muniz, F. J. (2019). Can Carob-Fruit-Extract-Enriched Meat Improve the Lipoprotein Profile, VLDL-Oxidation, and LDL Receptor Levels Induced by an Atherogenic Diet in STZ-NAD-Diabetic Rats? Nutrients, 11(2), 332. https://doi.org/10.3390/nu11020332