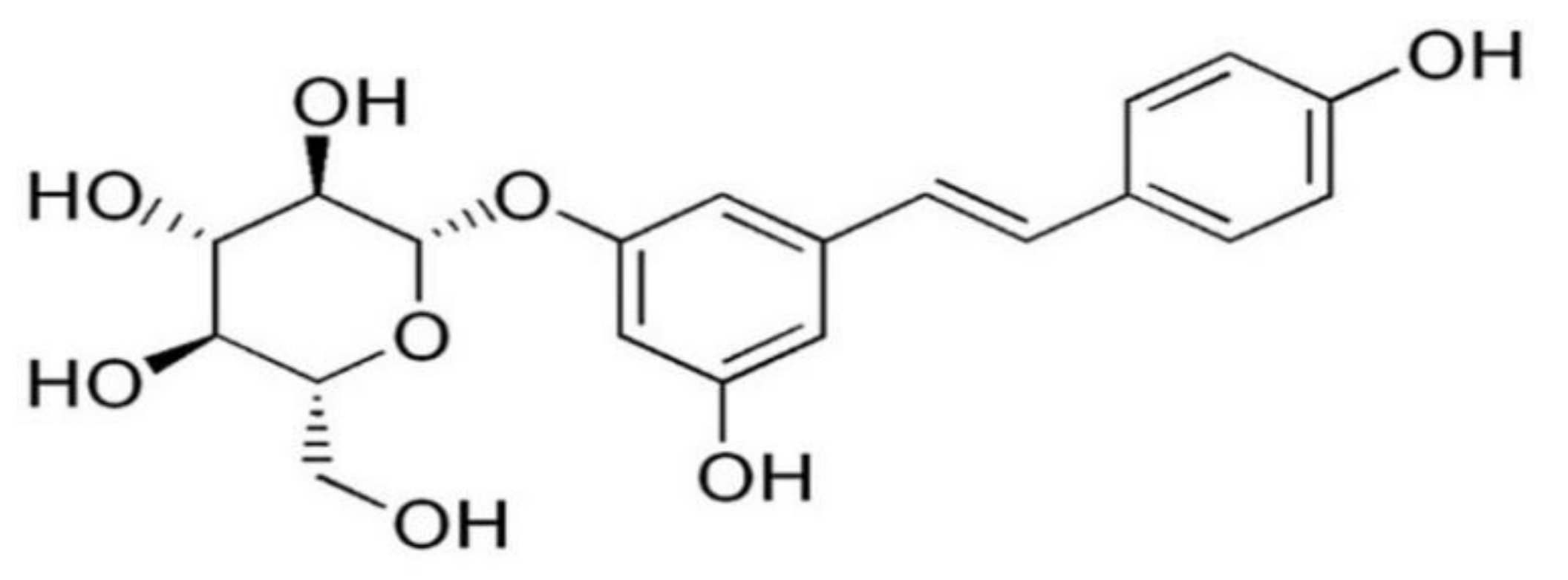
Polydatin Inhibits NLRP3 Inflammasome in Dry Eye Disease by Attenuating Oxidative Stress and Inhibiting the NF-κB Pathway

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals and Treatment
2.3. Tear Volume Measurement
2.4. Analysis of Corneal Irregularity
2.5. Ocular Surface Evaluation
2.6. Histology
2.7. Immunohistochemistry
2.8. Cell Cultures and Reagents
2.9. Western Blotting Analysis
2.10. Nuclear and Cytosol Protein Extraction
2.11. Quantitative Real-Time PCR
2.12. ROS Production Assay
2.13. Statistical Analysis
3. Results
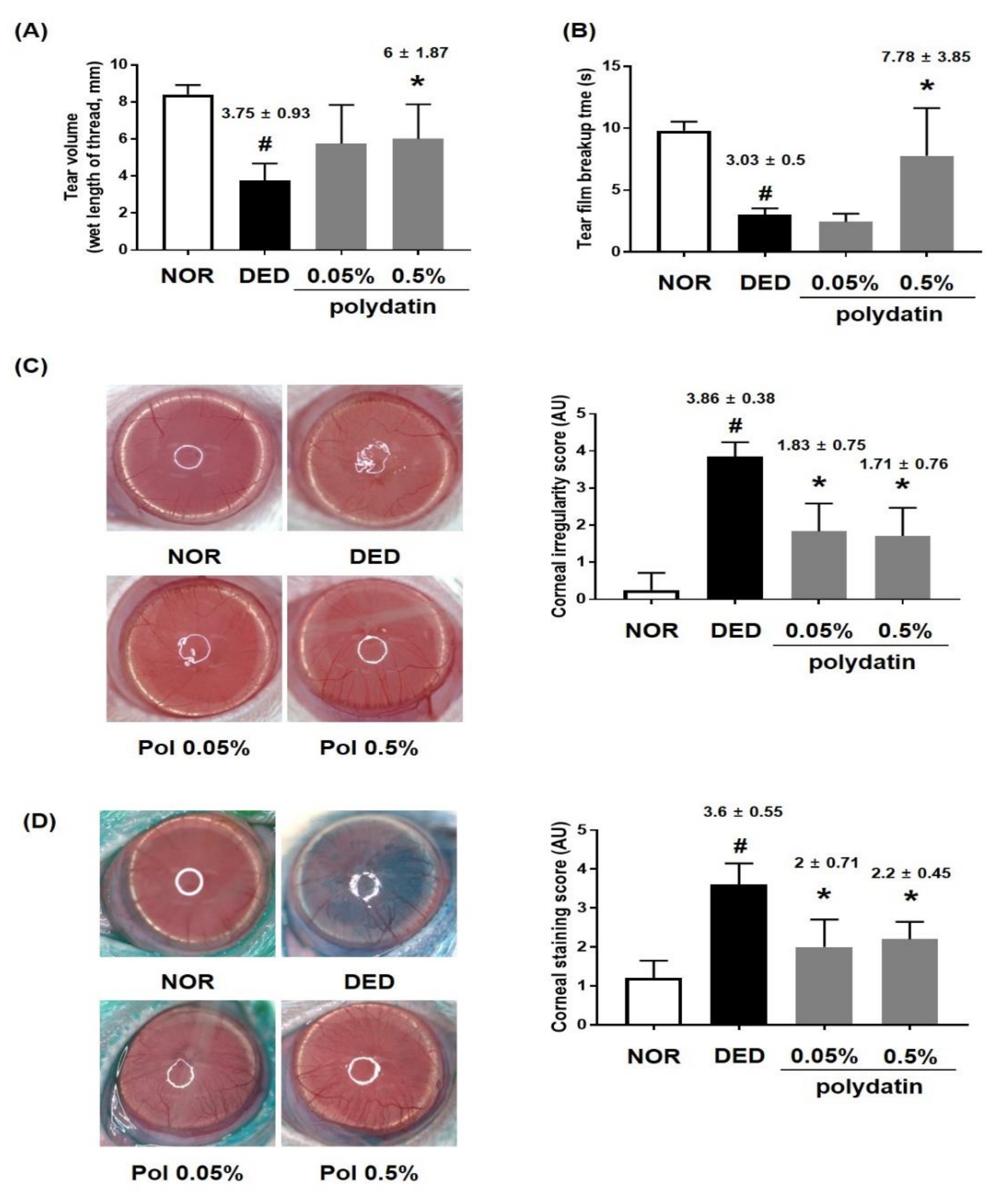
3.1. Effects of Polydatin on Dry Eye Disease In Vivo
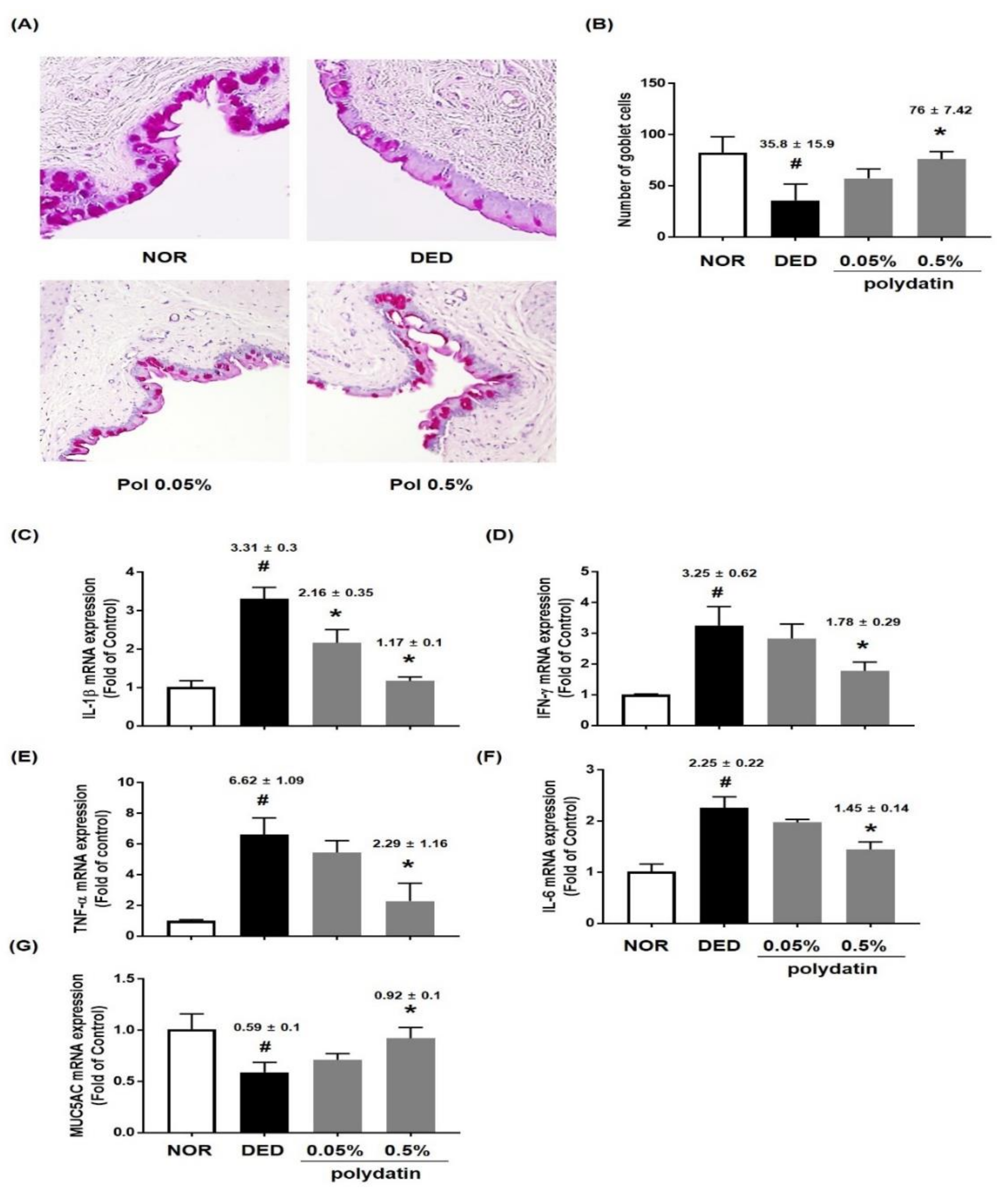
3.2. Effect of Polydatin on the Conjunctival Epithelium in Exorbital Lacrimal Gland-Excised Rats
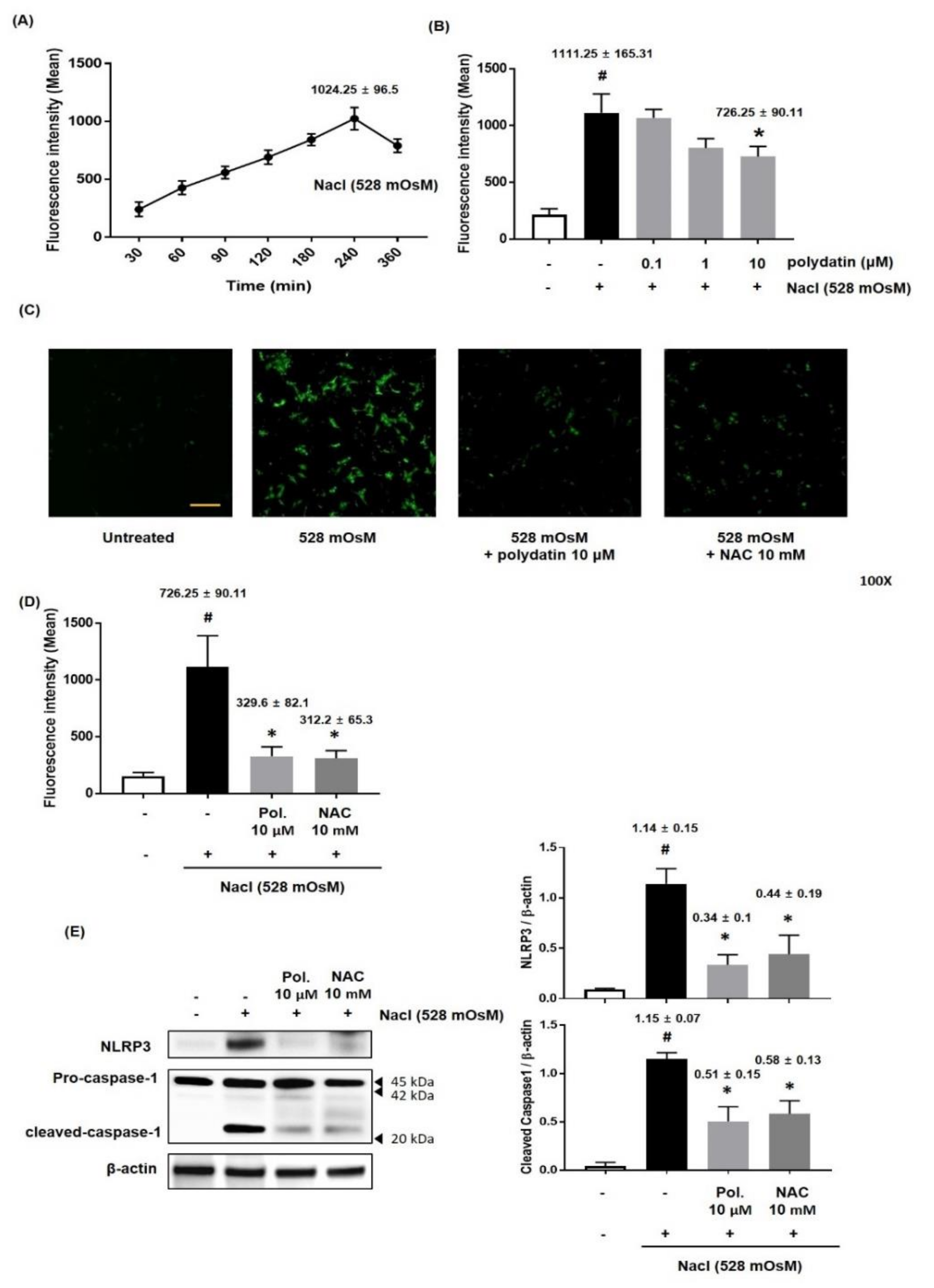
3.3. Effect of Polydatin on NLRP3 Inflammasome in Exorbital Lacrimal Gland-Excised Rats
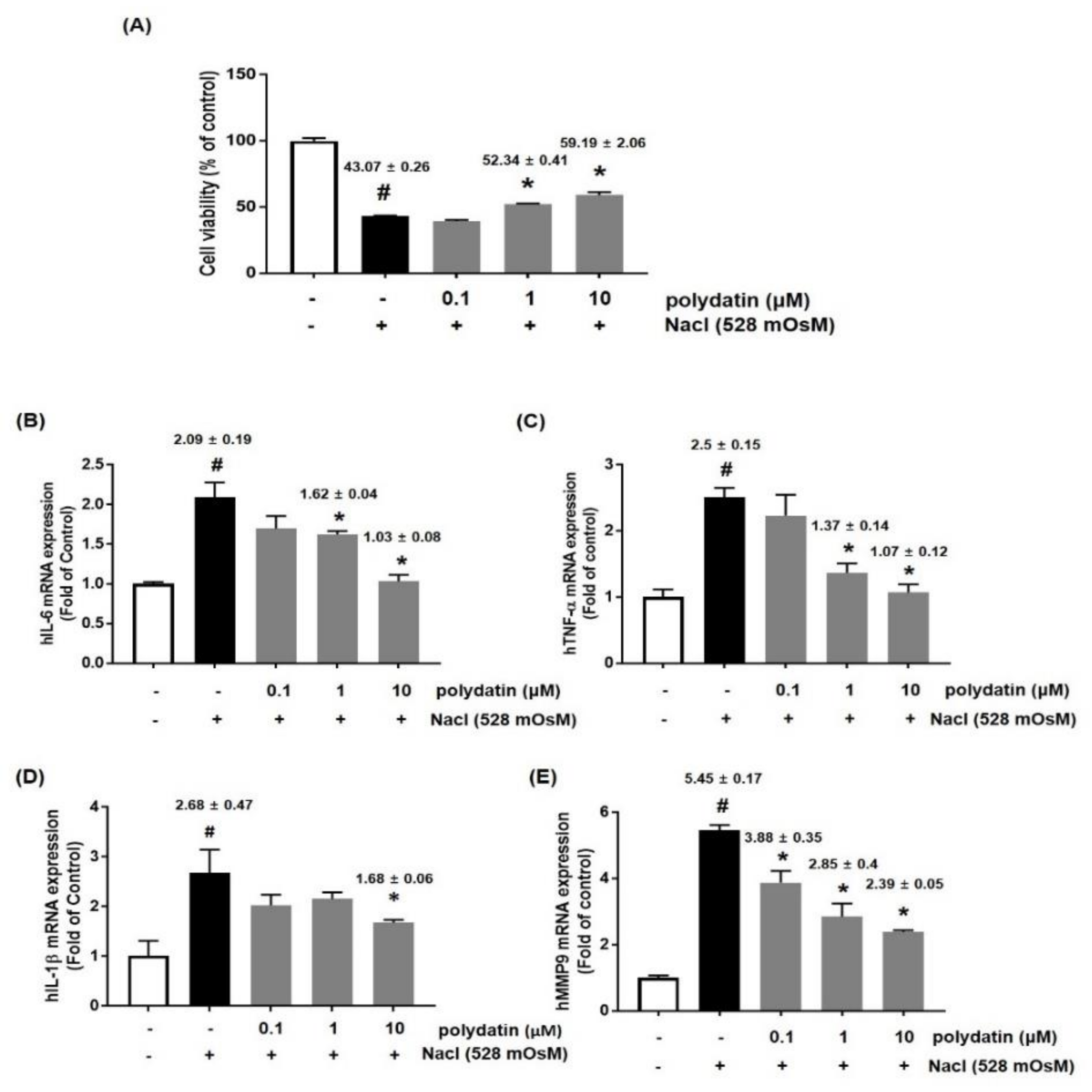
3.4. Effects of Polydatin on Hyperosmolar Stress-Induced Cytotoxicity and Inflammation
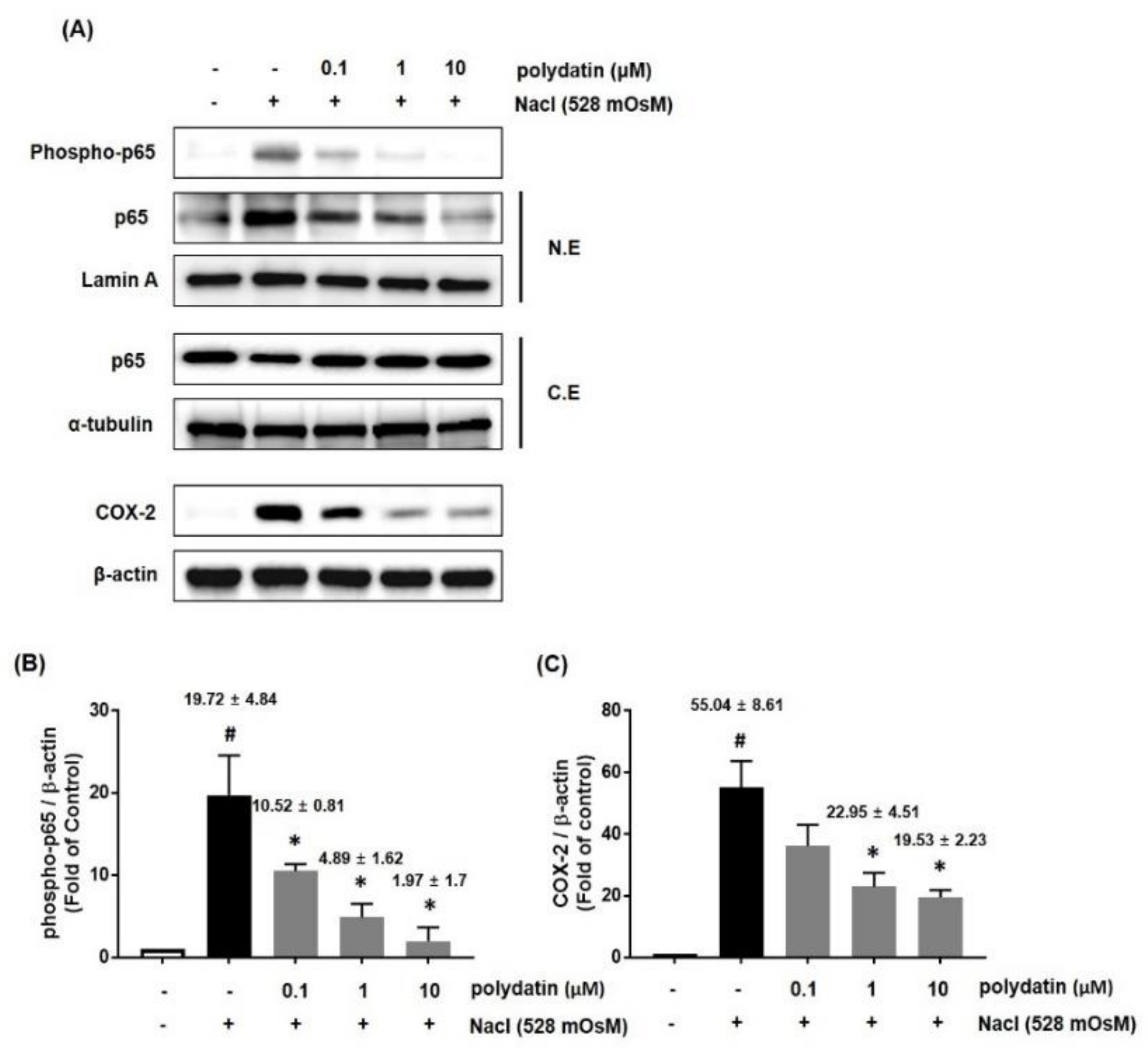
3.5. Effect of Polydatin on the Hyperosmolar Stress-Induced Inflammasome Signaling Pathway
3.6. Effect of Polydatin on ROS Production in Hyperosmolar Stress-Induced HCCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lemp, M.A.; Sullivan, B.D.; Crews, L.A. Biomarkers in dry eye disease. Eur. Ophthalmic Rev. 2012, 6, 157–163. [Google Scholar] [CrossRef]
- Narayanan, S.; Miller, W.L.; McDermott, A.M. Conjunctival Cytokine Expression in Symptomatic Moderate Dry Eye Subjects. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2445–2450. [Google Scholar] [CrossRef]
- Park, B.; Lee, I.S.; Hyun, S.-W.; Jo, K.; Lee, T.G.; Kim, J.S.; Kim, C.-S. The Protective Effect of Polygonum cuspidatum (PCE) Aqueous Extract in a Dry Eye Model. Nutrients 2018, 10, 1550. [Google Scholar] [CrossRef]
- Zheng, Q.; Ren, Y.; Reinach, P.S.; She, Y.; Xiao, B.; Hua, S.; Qu, J.; Chen, W. Reactive oxygen species activated NLRP3 inflammasomes prime environment-induced murine dry eye. Exp. Eye Res. 2014, 125, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.; Bartok, E.; Rieger, A.; Franchi, L.; Núñez, G.; Hornung, V. Cutting edge: Reactive oxygen species inhibitors block priming, but not activation, of the NLRP3 inflammasome. J. Immunol. 2011, 187, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Dixit, V.M. Inflammasomes: Guardians of cytosolic sanctity. Immunol. Rev. 2009, 227, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Kummer, J.A.; Broekhuizen, R.; Everett, H.; Agostini, L.; Kuijk, L.; Martinon, F.; Van Bruggen, R.; Tschopp, J. Inflammasome Components NALP 1 and 3 Show Distinct but Separate Expression Profiles in Human Tissues Suggesting a Site-specific Role in the Inflammatory Response. J. Histochem. Cytochem. 2007, 55, 443–452. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, Z.; Meng, Q.; Jiao, Y.; Xu, J.; Fan, S. Polydatin inhibits growth of lung cancer cells by inducing apoptosis and causing cell cycle arrest. Oncol. Lett. 2014, 7, 295–301. [Google Scholar] [CrossRef]
- Yan, X.-D.; Wang, Q.-M.; Tie, C.; Jin, H.-T.; Han, Y.-X.; Zhang, J.-L.; Yu, X.-M.; Hou, Q.; Zhang, P.-P.; Wang, A.-P.; et al. Polydatin protects the respiratory system from PM2.5 exposure. Sci. Rep. 2017, 7, 40030. [Google Scholar] [CrossRef]
- Xie, X.; Peng, J.; Huang, K.; Huang, J.; Shen, X.; Liu, P.; Huang, H. Polydatin ameliorates experimental diabetes-induced fibronectin through inhibiting the activation of NF-κB signaling pathway in rat glomerular mesangial cells. Mol. Cell. Endocrinol. 2012, 362, 183–193. [Google Scholar] [CrossRef]
- Xu, L.-Q.; Gui, S.-H.; Mo, Z.-Z.; Sun, C.-Y.; Luo, D.-D.; Zhang, Z.-B.; Su, Z.-R.; Xie, J.-H.; Xie, Y.-L.; Zhang, X.; et al. Polydatin attenuates d -galactose-induced liver and brain damage through its anti-oxidative, anti-inflammatory and anti-apoptotic effects in mice. Food Funct. 2016, 7, 4545–4555. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Sun, T.; Li, W.; Sun, X. Inhibiting effects of dietary polyphenols on chronic eye diseases. J. Funct. Foods 2017, 39, 186–197. [Google Scholar] [CrossRef]
- Kim, C.-S.; Jo, K.; Lee, I.-S.; Kim, J. Topical Application of Apricot Kernel Extract Improves Dry Eye Symptoms in a Unilateral Exorbital Lacrimal Gland Excision Mouse. Nutrients 2016, 8, 750. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tan, G.; Ding, X.; Wang, Y.; Wu, A.; Yang, Q.; Ye, L.; Shao, Y. A mouse dry eye model induced by topical administration of the air pollutant particulate matter 10. Biomed. Pharmacother. 2017, 96, 524–534. [Google Scholar] [CrossRef]
- Sohn, E.J.; Kim, C.-S.; Kim, Y.S.; Jung, D.H.; Jang, D.S.; Lee, Y.M.; Kim, J.S. Effects of magnolol (5,5′-diallyl-2,2′-dihydroxybiphenyl) on diabetic nephropathy in type 2 diabetic Goto–Kakizaki rats. Life Sci. 2007, 80, 468–475. [Google Scholar] [CrossRef]
- Yagci, A.; Gürdal, C. The role and treatment of inflammation in dry eye disease. Int. Ophthalmol. 2014, 34, 1291–1301. [Google Scholar] [CrossRef]
- Niu, L.; Zhang, S.; Wu, J.; Chen, L.; Wang, Y. Upregulation of NLRP3 Inflammasome in the Tears and Ocular Surface of Dry Eye Patients. PLoS ONE 2015, 10, e0126277. [Google Scholar] [CrossRef]
- Brignole-Baudouin, F.; Kessal, K.; Liang, H.; Rabut, G.; Baudouin, C.; Melik Parsadaniantz, S.; Daull, P.; Garrigue, J.-S.; Docquier, M. Conjunctival inflammatory gene expression profiling in dry eye disease: Correlations with HLA-DRA and HLA-DRB1. Front. Immunol. 2018, 9, 2271. [Google Scholar]
- Deng, R.; Hua, X.; Li, J.; Chi, W.; Zhang, Z.; Lu, F.; Zhang, L.; Pflugfelder, S.C.; Li, D.-Q. Oxidative Stress Markers Induced by Hyperosmolarity in Primary Human Corneal Epithelial Cells. PLoS ONE 2015, 10, e0126561. [Google Scholar] [CrossRef]
- Maynard, S.; Keijzers, G.; Gram, M.; Desler, C.; Bendix, L.; Budtz-Jørgensen, E.; Molbo, D.; Croteau, D.L.; Osler, M.; Stevnsner, T. Relationships between human vitality and mitochondrial respiratory parameters, reactive oxygen species production and dNTP levels in peripheral blood mononuclear cells. Aging 2013, 5, 850. [Google Scholar] [CrossRef]
- Devasagayam, T.P.A.; Tilak, J.C.; Boloor, K.K.; Sane, K.S.; Ghaskadbi, S.S.; Lele, R.D. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Physicians India 2004, 52, 4. [Google Scholar]
- Romero-Pérez, A.I.; Ibern-Gómez, M.; Lamuela-Raventós, R.M.; de la Torre-Boronat, M.C. Piceid, the major resveratrol derivative in grape juices. J. Agric. Food Chem. 1999, 47, 1533–1536. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, W.; Chauhan, S.K.; Dana, R. Dry eye disease: An immune-mediated ocular surface disorder. Arch. Ophthalmol. 2012, 130, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Gipson, I.K.; Argüeso, P.; Beuerman, R.; Bonini, S.; Butovich, I.; Dana, R.; Dartt, D.; Gamache, D.; Ham, B.; Jumblatt, M. Research in dry eye: Report of the Research Subcommittee of the International Dry Eye WorkShop (2007). Ocul. Surf. 2007, 5, 179–193. [Google Scholar]
- Stevenson, W.; Chen, Y.; Lee, S.-M.; Lee, H.S.; Hua, J.; Dohlman, T.; Shiang, T.; Dana, R. Extraorbital lacrimal gland excision: A reproducible model of severe aqueous tear-deficient dry eye disease. Cornea 2014, 33, 1336–1341. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Nikulina, K.; Lazarev, S.; Bahrami, A.F.; Noble, L.B.; Gallup, M.; McNamara, N.A. Interleukin-1 as a Phenotypic Immunomodulator in Keratinizing Squamous Metaplasia of the Ocular Surface in Sjögren’s Syndrome. Am. J. Pathol. 2010, 177, 1333–1343. [Google Scholar] [CrossRef]
- De Paiva, C.S.; Villarreal, A.L.; Corrales, R.M.; Rahman, H.T.; Chang, V.Y.; Farley, W.J.; Stern, M.E.; Niederkorn, J.Y.; Li, D.-Q.; Pflugfelder, S.C. Dry eye–induced conjunctival epithelial squamous metaplasia is modulated by interferon-γ. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2553–2560. [Google Scholar] [CrossRef]
- Corrales, R.M. Desiccating Stress Stimulates Expression of Matrix Metalloproteinases by the Corneal Epithelium. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3293–3302. [Google Scholar] [CrossRef]
- Eisenbarth, S.C.; Flavell, R.A. Innate instruction of adaptive immunity revisited: The inflammasome. EMBO Mol. Med. 2009, 1, 92–98. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef]
- Jo, E.-K.; Kim, J.K.; Shin, D.-M.; Sasakawa, C. Molecular mechanisms regulating NLRP3 inflammasome activation. Cell. Mol. Immunol. 2016, 13, 148. [Google Scholar] [CrossRef]
- Zheng, Q.; Ren, Y.; Reinach, P.S.; Xiao, B.; Lu, H.; Zhu, Y.; Qu, J.; Chen, W. Reactive oxygen species activated NLRP3 inflammasomes initiate inflammation in hyperosmolarity stressed human corneal epithelial cells and environment-induced dry eye patients. Exp. Eye Res. 2015, 134, 133–140. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence | |
|---|---|---|
| rIL-1β | Sense | 5’-CCAGGATGAGGACCCAAGCA-3’ |
| antisense | 5’-TCCCGACCATTGCTGTTTCC-3’ | |
| rIL-6 | Sense | 5’-AGAGACTTCCAGCCAGTTGC-3’ |
| antisense | 5’-AGCCTCCGACTTGTGAAGTG-3’ | |
| rTNF-α | Sense | 5’-TCGTCTACTCCTCAGAGCCC-3’ |
| antisense | 5’-ACTTCAGCGTCTCGTGTGTT-3’ | |
| rMUC5AC | Sense | 5’-TCCGGCCTCATCTTCTCC-3’ |
| antisense | 5’-ACTTGGGCACTGGTGCTG-3’ | |
| rIFN-γ | Sense | 5’-ATCTGGAGGAACTGGCAAAAGGACG-3’ |
| antisense | 5’-CCTTAGGCTAGATTCTGGTGACAGC-3’ | |
| rNLRP3 | Sense | 5’-ATCTGGAGGAACTGGCAAAAGGACG-3’ |
| antisense | 5’-CCTTAGGCTAGATTCTGGTGACAGC-3’ | |
| rGAPDH | Sense | 5’-GGGACTCAAGCTCCTCTGTG-3’ |
| antisense | 5’-GAGGCTCTGGTTATGGGTCA-3’ | |
| hIL-1β | Sense | 5’-AATCTGTACCTGTCCTGCGTGTT-3’ |
| antisense | 5’-TGGGTAATTTTTGGGATCTACACTCT-3’ | |
| hIL-6 | Sense | 5’-AAATTCGGTACATCCTCGAC-3’ |
| antisense | 5’-CAGGAACTGGATCAGGACTT-3’ | |
| hTNF-α | Sense | 5’-TTCTCCTTCCTGCTTGTG-3’ |
| antisense | 5’-CTGAGTGTGAGTGTCTGG-3’ | |
| hMMP9 | Sense | 5’-GGGACGCAGACATCGTCATC-3’ |
| antisense | 5’-TCGTCATCGTCGAAATGGGC-3’ | |
| hGAPDH | Sense | 5’-CCAGCCGAGCCACATCGCTC-3’ |
| antisense | 5’-ATGAGCCCCAGCCTTCTCCAT-3’ | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, B.; Jo, K.; Lee, T.G.; Hyun, S.-W.; Kim, J.S.; Kim, C.-S. Polydatin Inhibits NLRP3 Inflammasome in Dry Eye Disease by Attenuating Oxidative Stress and Inhibiting the NF-κB Pathway. Nutrients 2019, 11, 2792. https://doi.org/10.3390/nu11112792
Park B, Jo K, Lee TG, Hyun S-W, Kim JS, Kim C-S. Polydatin Inhibits NLRP3 Inflammasome in Dry Eye Disease by Attenuating Oxidative Stress and Inhibiting the NF-κB Pathway. Nutrients. 2019; 11(11):2792. https://doi.org/10.3390/nu11112792
Chicago/Turabian StylePark, Bongkyun, Kyuhyung Jo, Tae Gu Lee, Soo-Wang Hyun, Jin Sook Kim, and Chan-Sik Kim. 2019. "Polydatin Inhibits NLRP3 Inflammasome in Dry Eye Disease by Attenuating Oxidative Stress and Inhibiting the NF-κB Pathway" Nutrients 11, no. 11: 2792. https://doi.org/10.3390/nu11112792
APA StylePark, B., Jo, K., Lee, T. G., Hyun, S.-W., Kim, J. S., & Kim, C.-S. (2019). Polydatin Inhibits NLRP3 Inflammasome in Dry Eye Disease by Attenuating Oxidative Stress and Inhibiting the NF-κB Pathway. Nutrients, 11(11), 2792. https://doi.org/10.3390/nu11112792