Potential Mechanisms of Probiotics Action in the Prevention and Treatment of Colorectal Cancer



Abstract
1. Introduction
- Lactobacillus casei Shirota has an inhibitory effect on colorectal cancer and bladder cancer. It exhibits positive effects on maintaining the balance of intestinal microbiome, and also protects against intestinal disorders. It has immunomodulatory effects and can strengthen the immune defense of the host by inducing IL–12 (interleukin 12) production through phagocytes. It supports the treatment of colorectal cancer, decreases the activity of the fecal enzymes, and protects against mutagens from food,
- Lactobacillus fermentum NCIMB 5221 is potentially able to modulate hyperinsulinemia, insulin resistance, hypercholesterolemia, and hypertriglyceridemia. It has an antiproliferative effect,
- Weissella cibaria JW15 strengthens the function of the immune system by increasing the activity of NK cells (Natural Killer Cells),
- Saccharomyces cerevisiae var. boulardii has anti-inflammatory and antibacterial effects. It increases the secretion of immunoglobulin A (IgA) and maintains the integrity of the epithelial barrier. It helps in the treatment of travelers’ diarrhea.
2. Gastrointestinal Microbiota
Functions of the Intestinal Microbiota
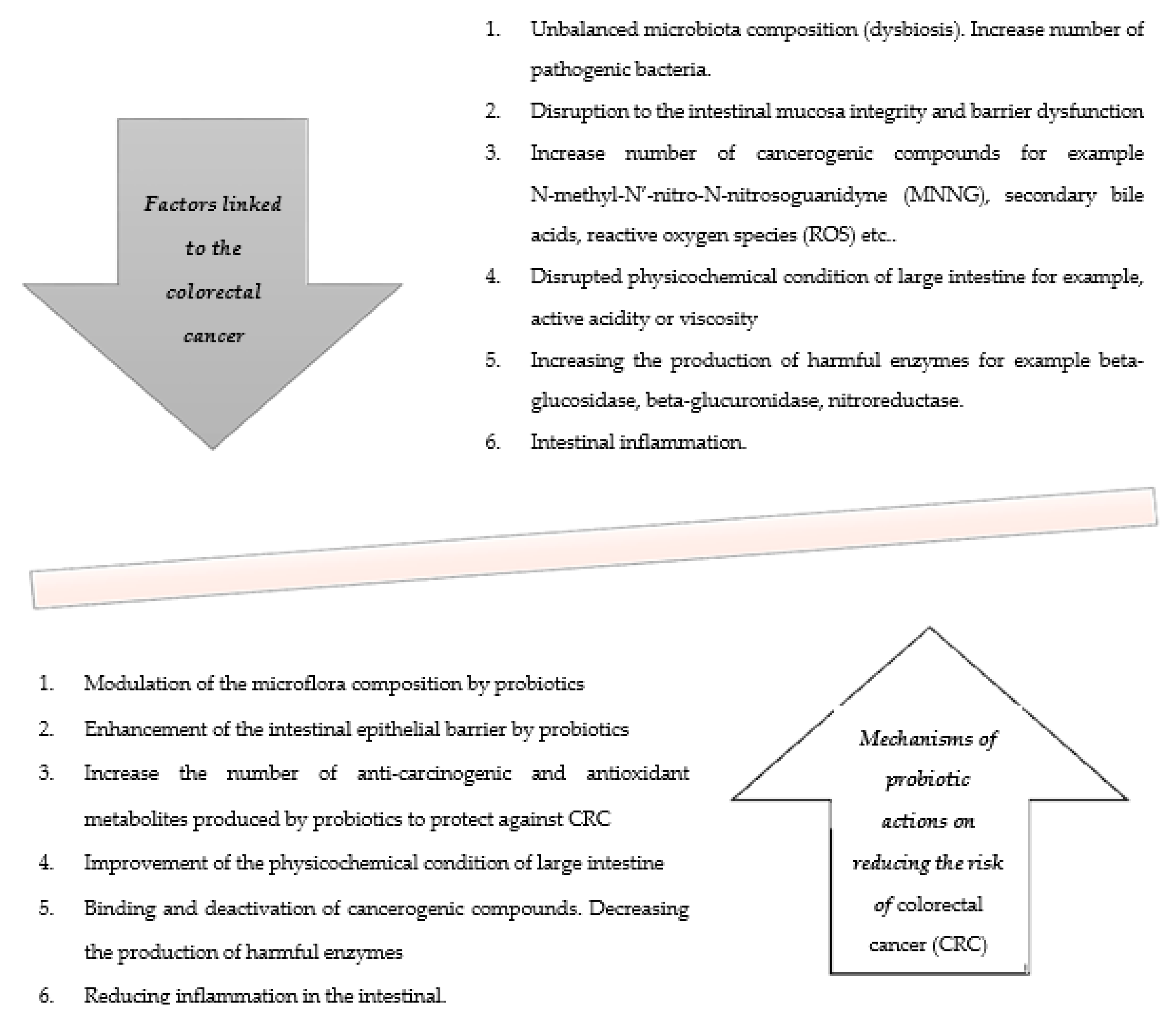
3. Mechanisms of Probiotic Actions for Reducing the Risk of Colorectal Cancer (CRC)
3.1. Modulation of the Microbiota Composition by Probiotics
3.2. Enhancement of the Intestinal Epithelial Barrier by Probiotics
3.3. Increase the Number of Anti-Carcinogenic and Antioxidant Metabolites Produced by Probiotics to Protect Against Colorectal Cancer (CRC)
3.4. Improvement of the Physicochemical Condition of Large Intestine
3.5. Binding and Deactivation of Cancerogenic Compounds. Decreasing the Production of Harmful Enzymes
3.6. Reducing Inflammation in the Intestine
- interact with TLR (Toll-like) receptors. TLRs play an important role in the initiation of the immune response, as well as the recognition of the threat,
- affect the production of IL–8 (interleukin 8) needed for neutrophilia,
- induce the production of anti-inflammatory cytokines, inhibit NF–kB in macrophages, and initiate the production of TNF (tumor necrosis factor, which is a pleiotropic cytokine involved in the pathogenesis of many physiological processes that control inflammation or antineoplastic response etc.),
- prevent the activation of the NF–kB transcription factor, which plays a key role in activating the immune system to various stimuli
- modify the MAPK kinase (mitogen-activated protein kinases) signaling pathway and the PPAR–γ receptor.
4. A Potential Dose of Probiotic Therapy Can Bring Health Benefits
- The number of probiotic bacteria consumed is an important factor to obtain a high concentration of bacteria in various sections of the digestive tract,
- The concentration of probiotics should be greater than or equal to 106 CFU/ml in the small intestine (ileum) and 108 CFU/g in the large intestine (the strength of scientific studies confirming this thesis is weak),
- Concentration in the colon has been proposed because, in justified cases, it can be expected that the bacterial flora is more likely to be more active than the flora present at lower levels.
5. The Importance of Probiotics in the Prevention and Treatment of Cancer Tumors
6. Probiotics and Operations
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- American Cancer Society Colorectal Cancer Facts & Figures 2017–2019; American Cancer Society: Atlanta, GA, USA, 2017; pp. 1–40.
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Tárraga López, P.J.; Albero, J.S.; Rodríguez-Montes, J.A. Primary and Secondary Prevention of Colorectal Cancer. Clin. Med. Insights Gastroenterol. 2014, 7, 33–46. [Google Scholar] [PubMed]
- Wronkowski, Z.; Brużewicz, S. Malignant neoplasms of the large intestine. General information. In Colorectal Cancer; PZWL Medical Publisher: Warsaw, Poland, 2008; pp. 25–40. ISBN 978-83-200-3333-5. (In Polish) [Google Scholar]
- Food and Agriculture Organization (FAO); World Health Organization (WHO). Guidelines for the Evaluation of Probiotics in Food; FAO/WHO: London, ON, Canada, 2002; pp. 1–11. [Google Scholar]
- Fiedurek, J. (Ed.) Probiotics. In Microbiome and Human Health; University of Maria Curie-Skłodowska Publishing House: Lublin, Poland, 2014; pp. 124–125. (In Polish) [Google Scholar]
- Fijan, S. Microorganisms with Claimed Probiotic Properties: An Overview of Recent Literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef] [PubMed]
- Jach, M.; Łoś, R.; Maj, M.; Malm, A. Probiotics—technological and manufacturing aspects. Post. Mikrobiol. 2013, 52, 161–170. (In Polish) [Google Scholar]
- Mojka, K. Probiotics, prebiotics and synbiotics—Characteristics and functions. Probl. Hig. Epidemiol. 2014, 95, 541–549. (In Polish) [Google Scholar]
- Kahouli, I.; Malhotra, M.; Alaoui-Jamali, M.A.; Prakash, S. In-Vitro Characterization of the Anti-Cancer Activity of the Probiotic Bacterium Lactobacillus Fermentum NCIMB 5221 and Potential against Colorectal Cancer Cells. J. Cancer Sci. Ther. 2015, 7, 224–235. [Google Scholar]
- Shida, K.; Nomoto, K. Probiotics as efficient immunopotentiators: Translational role in cancer prevention. Indian J. Med. Res. 2013, 138, 808–814. [Google Scholar]
- Lee, Y.J.; Lee, A.; Yoo, H.J.; Kim, M.; Noh, G.M.; Lee, J.H. Supplementation with the probiotic strain Weissella cibaria JW15 enhances natural killer cell activity in nondiabetic subjects. J. Funct. Foods 2018, 48, 153–158. [Google Scholar] [CrossRef]
- Łukaszewicz, M. Saccharomyces cerevisiae var. boulardii—Probiotic Yeast. In Probiotics; IntechOpen: London, UK, 2012; pp. 385–398. [Google Scholar]
- Stier, H.; Bischoff, S.C. Influence of Saccharomyces boulardii CNCM I-745 on the gut-associated immune system. Clin. Exp. Gastroenterol. 2016, 9, 269–279. [Google Scholar] [CrossRef]
- Liong, M.T. Roles of Probiotics and Prebiotics in Colon Cancer Prevention: Postulated Mechanisms and In-vivo Evidence. Int. J. Mol. Sci. 2008, 9, 854–863. [Google Scholar] [CrossRef]
- Wasilewska, E.; Złotkowska, D.; Pijagin, M.E. The role of intestinal microflora and probiotic bacteria in prophylactic and development of colorectal cancer. Postępy Hig. Med. Dosw. 2013, 67, 837–847. (In Polish) [Google Scholar] [CrossRef]
- Libudzisz, Z. Microflora of the human digestive tract and its effect on the body. In Microorganisms in Food and Nutrition]; Gawęcki, J., Libudzisz, Z., Eds.; Publisher of the University of Life Sciences: Poznan, Poland, 2016; pp. 31–40. ISBN 978-83-7160-776-9. (In Polish) [Google Scholar]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef] [PubMed]
- Niederreiter, L.; Adolph, T.E.; Tilg, H. Food, microbiome and colorectal cancer. Dig. Liver Dis. 2018, 50, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Tiihonen, K.; Ouwehand, A.C.; Rautonen, N. Human intestinal microbiota and healthy ageing. Ageing Res. Rev. 2010, 9, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, M.; Tokarz-Deptuła, B.; Deptuła, W. The human microbiome. Post. Mikrobiol. 2017, 56, 33–42. (In Polish) [Google Scholar]
- Górska, S.; Jarząb, A.; Gamian, A. Probiotic bacteria in the human gastrointestinal tract as a factor stimulating the immune system. Postepy Hig. Med. Dosw. 2009, 63, 653–667. (In Polish) [Google Scholar]
- Krakowiak, O.; Nowak, R. Human digestive tract microflora—Significance, development, modification. Post Fitoter 2015, 3, 193–200. (In Polish) [Google Scholar]
- Moraes-Filho, J.P.; Quigley, E.M. the Intestinal Microbiota and the Role of Probiotics in Irritable Bowel Syndrome: A review. Arq. Gastroenterol. 2015, 52, 331–338. [Google Scholar] [CrossRef]
- Dos Reis, S.A.; da Conceição, L.L.; Siqueira, N.P.; Rosa, D.D.; da Silva, L.L.; Peluzio, M.D. Review of the mechanisms of probiotic actions in the prevention of colorectal cancer. Nutr. Res. 2017, 37, 1–19. [Google Scholar] [CrossRef]
- Akin, H.; Tözün, N. Diet, microbiota, and colorectal cancer. J. Clin. Gastroenterol. 2014, 48, S67–S69. [Google Scholar] [CrossRef]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Tran Van Nhieu, J.; Furet, J.P. Microbial Dysbiosis in Colorectal Cancer (CRC) Patients. PLoS ONE 2011, 6, e16393. [Google Scholar] [CrossRef] [PubMed]
- Kahouli, I.; Tomaro-Duchesneau, C.; Prakash, S. Probiotics in colorectal cancer (CRC) with emphasis on mechanisms of action and current perspectives. J. Med. Microbiol. 2013, 62, 1107–1123. [Google Scholar] [CrossRef] [PubMed]
- Uccello, M.; Malaguarnera, G.; Basile, F.; D’agata, V.; Malaguarnera, M.; Bertino, G.; Vacante, M.; Drago, F.; Biondi, A. Potential role of probiotics on colorectal cancer prevention. BMC Surg. 2012, 12, S35. [Google Scholar] [CrossRef]
- Koziński, K.; Dobrzyń, A. Wnt signaling pathway—Its role in regulation of cell metabolism. Postępy Hig. Med. Dosw. 2013, 67, 1098–1108. (In Polish) [Google Scholar] [CrossRef]
- Boleij, A.; Hechenbleikner, E.M.; Goodwin, A.C.; Badani, R.; Stein, E.M.; Lazarev, M.G.; Ellis, B.; Carroll, K.C.; Albesiano, E.; Wick, E.C.; et al. The Bacteroides fragilis Toxin Gene Is Prevalent in the Colon Mucosa of Colorectal Cancer Patients. Clin. Infect. Dis. 2015, 60, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Sears, C.L.; Geis, A.L.; Housseau, F. Bacteroides fragilis subverts mucosal biology: From symbiont to colon carcinogenesis. J. Clin. Investig. 2014, 124, 4166–4172. [Google Scholar] [CrossRef]
- Ambalam, P.; Raman, M.; Purama, R.K.; Doble, M. Probiotics, prebiotics and colorectal cancer prevention. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 119–131. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Meyerson, M.; Garrett, W.S. Microbes and inflammation in colorectal cancer. Cancer Immunol. Res. 2013, 1, 150–157. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef]
- Karim, B.O.; Huso, D.L. Mouse models for colorectal cancer. Am. J. Cancer Res. 2013, 3, 240–250. [Google Scholar] [PubMed]
- Bonnet, M.; Buc, E.; Sauvanet, P.; Darcha, C.; Dubois, D.; Pereira, B.; Déchelotte, P.; Bonnet, R.; Pezet, D.; Darfeuille-Michaud, A. Colonization of the Human Gut by E. coli and Colorectal Cancer Risk. Clin. Cancer Res. 2014, 20, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Buc, E.; Dubois, D.; Sauvanet, P.; Raisch, J.; Delmas, J.; Darfeuille-Michaud, A.; Pezet, D.; Bonnet, R. High Prevalence of Mucosa-Associated, E. coli Producing Cyclomodulin and Genotoxin in Colon Cancer. PLoS ONE 2013, 8, e56964. [Google Scholar] [CrossRef] [PubMed]
- Gosai, V.; Ambalam, P.; Raman, M.; Kothari, C.R.; Kothari, R.K.; Vyas, B.R.M.; Sheth, N.R. Protective effect of Lactobacillus rhamnosus 231 against N-Methyl-N’-nitro-N-nitrosoguanidine in animal model. Gut Microbes 2011, 2, 319–325. [Google Scholar] [CrossRef][Green Version]
- Rao, R.K.; Samak, G. Protection and Restitution of Gut Barrier by Probiotics: Nutritional and Clinical Implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar]
- Liu, D.; Jiang, X.Y.; Zhou, L.S.; Song, J.H.; Zhang, X. Effects of Probiotics on Intestinal Mucosa Barrier in Patients with Colorectal Cancer after Operation: Meta-Analysis of Randomized Controlled Trials. Medicine 2016, 95, e3342. [Google Scholar] [CrossRef]
- Madsen, K.L. Enhancement of Epithelial Barrier Function by Probiotics. J. Epithel. Biol. Pharmacol. 2012, 5, 55–59. [Google Scholar] [CrossRef]
- Hsieh, C.Y.; Osaka, T.; Moriyama, E.; Date, Y.; Kikuchi, J.; Tsuneda, S. Strengthening of the intestinal epithelial tight junction by Bifidobacterium bifidum. Physiol. Rep. 2015, 3, e12327. [Google Scholar] [CrossRef]
- Blackwood, B.P.; Yuan, C.Y.; Wood, D.R.; Nicolas, J.D.; Grothaus, J.S.; Hunter, C.J. Probiotic Lactobacillus Species Strengthen Intestinal Barrier Function and Tight Junction Integrity in Experimental Necrotizing Enterocolitis. J. Probiotics Health 2017, 5. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, L.; Dou, X.; Wang, C.; Zhang, W.; Gao, K.; Liu, J.; Wang, H. Lactobacillus reuteri ZJ617 maintains intestinal integrity via regulating tight junction, autophagy and apoptosis in mice challenged with lipopolysaccharide. Oncotarget 2017, 8, 77489–77499. [Google Scholar] [CrossRef]
- Konieczna, C.; Olejnik-Schmidt, A.; Schmidt, M.T. Lactobacillus spp. belonging to the Casei group display a variety of adhesins. Acta Sci. Pol. Technol. Aliment. 2018, 17, 69–82. [Google Scholar] [PubMed]
- Czajkowska, A.; Szponar, B. Short chain fatty acids (SCFA), the products of gut bacteria metabolism and their role in the host. Postępy Hig. Med. Dosw. 2018, 72, 131–142. (In Polish) [Google Scholar] [CrossRef]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary Gut Microbial Metabolites, Short-chain Fatty Acids, and Host Metabolic Regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Yu, Y.N.; Wang, J.L.; Lin, Y.W.; Kong, X.; Yang, C.Q.; Yang, L.; Liu, Z.J.; Yuan, Y.Z.; Liu, F.; et al. Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 2013, 97, 1044–1052. [Google Scholar] [CrossRef]
- Ewaschuk, J.B.; Walker, J.W.; Diaz, H.; Madsen, K.L. Bioproduction of Conjugated Linoleic Acid by Probiotic Bacteria Occurs In Vitro and In Vivo in Mice. J. Nutr. 2006, 136, 1483–1487. [Google Scholar] [CrossRef]
- Bassaganya-Riera, J.; Viladomiu, M.; Pedragosa, M.; De Simone, C.; Carbo, A.; Shaykhutdinov, R.; Jobin, C.; Arthur, J.C.; Corl, B.A.; Vogel, H.; et al. Probiotic Bacteria Produce Conjugated Linoleic Acid Locally in the Gut That Targets Macrophage PPAR γ to Suppress Colitis. PLoS ONE 2012, 7, e31238. [Google Scholar] [CrossRef]
- Kruszewska, D.; Lan, J.; Lorca, G.; Yanagisawa, N.; Marklinder, I.; Ljungh, Å. Selection of lactic acid bacteria as probiotic strains by in vitro tests. Microecol. Ther. 2002, 29, 37–49. [Google Scholar]
- Lu, C.; Cheng, S.Y. Thyroid hormone receptors regulate adipogenesis and carcinogenesis via crosstalk signaling with peroxisome proliferator-activated receptors. J. Mol. Endocrinol. 2010, 44, 143–154. [Google Scholar] [CrossRef]
- Hojka, A.; Rapak, A. Peroxisome proliferator-activated receptors (PPAR). Antiproliferative properties. Postepy Hig. Med. Dosw. 2011, 65, 404–413. (In Polish) [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Lan, A.; Lagadic-Gossmann, D.; Lemaire, C.; Brenner, C.; Jan, G. Acidic extracellular pH shifts colorectal cancer cell death from apoptosis to necrosis upon exposure to propionate and acetate, major end-products of the human probiotic propionibacteria. Apoptosis 2007, 12, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Libudzisz, Z. Carcinogenic activity of intestinal microorganisms. Żywn. Nauka Technol. Jakość 2008, 6, 25–39. (In Polish) [Google Scholar]
- LeBlanc, J.G.; LeBlanc, A. Probiotics in Inflammatory Bowel Diseases and Cancer Prevention. In Probiotics, Prebiotics, and Synbiotics Bioactive Foods in Health Promotion; Watson, R.R., Preedy, V.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 755–771. ISBN 978-0-12-802189-7. [Google Scholar]
- Walia, S.; Kamal, R.; Dhawan, D.K.; Kanwar, S.S. Chemoprevention by Probiotics During 1,2-Dimethylhydrazine-Induced Colon Carcinogenesis in Rats. Dig. Dis. Sci. 2018, 63, 900–909. [Google Scholar] [CrossRef]
- Kuśmierska, A.; Fol, M. Immunomodulatory and therapeutic properties of probiotic microorganisms. Probl. Hig. Epidemiol. 2014, 95, 529–540. (In Polish) [Google Scholar]
- Ashraf, R.; Vasiljevic, T.; Day, S.L.; Smith, S.C.; Donkor, O.N. Lactic acid bacteria and probiotic organisms induce different cytokine profile and regulatory T cells mechanisms. J. Funct. Foods 2014, 6, 395–409. [Google Scholar] [CrossRef]
- Ding, Y.H.; Qian, L.Y.; Pang, J.; Lin, J.Y.; Xu, Q.; Wang, L.H.; Huang, D.S.; Zou, H. The regulation of immune cells by Lactobacilli: A potential therapeutic target for anti-atherosclerosis therapy. Oncotarget 2017, 8, 59915–59928. [Google Scholar] [CrossRef]
- Majewska, M.; Szczepanik, M. The role of toll-like receptors (TLR) in innate and adaptive immune responses and their function in immune response regulation]. Postepy Hig. Med. Dosw. 2006, 60, 52–63. (In Polish) [Google Scholar]
- Mehta, A.K.; Gracias, D.T.; Croft, M. TNF Activity and T cells. Cytokine 2018, 101, 14–18. [Google Scholar] [CrossRef]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, Prebiotics and Immunomodulation of Gut Mucosal Defences: Homeostasis and Immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef]
- McCudden, C.R.; Hains, M.D.; Kimple, R.J.; Siderovski, D.P.; Willard, F.S. G-protein signaling: Back to the future. Cell. Mol. Life Sci. 2005, 62, 551–577. [Google Scholar] [CrossRef]
- Duc, N.M.; Kim, H.R.; Chung, K.Y. Structural mechanism of G protein activation by G protein-coupled receptor. Eur. J. Pharmacol. 2015, 763, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Henc, I.; Bryl, E. Chemokines as important mediators of inflammation. Forum Med. Rodz. 2013, 7, 251–262. (In Polish) [Google Scholar]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed]
- Libudzisz, Z. Probiotic food. In Microorganisms in Food and Nutrition; Gawęcki, J., Libudzisz, Z., Eds.; University of Life Sciences in Poznan Publishing House: Poznan, Poland, 2016; pp. 91–100. ISBN 978-83-7160-776-9. (In Polish) [Google Scholar]
- Maleki, D.; Homayouni, A.; Khalili, L.; Golkhalkhali, B. Probiotics in Cancer Prevention, Updating the Evidence. In Probiotics, Prebiotics, and Synbiotics; Watson, R.R., Preedy, V.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 781–791. ISBN 978-0-12-802189-7. [Google Scholar]
- Aureli, P.; Capurso, L.; Castellazzi, A.M.; Clerici, M.; Giovannini, M.; Morelli, L.; Poli, A.; Pregliasco, F.; Salvini, F.; Zuccotti, G.V. Probiotics and health: An evidence-based review. Pharmacol. Res. 2011, 63, 366–376. [Google Scholar] [CrossRef]
- Marteau, P.; Pochart, P.; Flourié, B.; Pellier, P.; Santos, L.; Desjeux, J.F.; Rambaud, J.C. Effect of chronic ingestion of a fermented dairy product containing Lactobacillus acidophilus and Bifidobacterium bifidum on metabolic activities of the colonic flora in humans. Am. J. Clin. Nutr. 1990, 52, 685–688. [Google Scholar] [CrossRef]
- Baldwin, C.; Millette, M.; Oth, D.; Ruiz, M.T.; Luquet, F.M.; Lacroix, M. Probiotic Lactobacillus acidophilus and L. casei mix sensitize colorectal tumoral cells to 5-fluorouracil-induced apoptosis. Nutr. Cancer 2010, 62, 371–378. [Google Scholar] [CrossRef]
- Österlund, P.; Ruotsalainen, T.; Korpela, R.; Saxelin, M.; Ollus, A.; Valta, P.; Kouri, M.; Elomaa, I.; Joensuu, H. Lactobacillus supplementation for diarrhoea related to chemotherapy of colorectal cancer: A randomised study. Br. J. Cancer 2007, 97, 1028–1034. [Google Scholar] [CrossRef]
- Wada, M.; Nagata, S.; Saito, M.; Shimizu, T.; Yamashiro, Y.; Matsuki, T.; Asahara, T.; Nomoto, K. Effects of the enteral administration of Bifidobacterium breve on patients undergoing chemotherapy for pediatric malignancies. Supportive Care Cancer 2010, 18, 751–759. [Google Scholar] [CrossRef]
- Urbancsek, H.; Kazar, T.; Mezes, I.; Neumann, K. Results of a double-blind, randomized study to evaluate the efficacy and safety of Antibiophilus® in patients with radiation-induced diarrhoea. Eur. J. Gastroenterol. Hepatol. 2001, 13, 391–396. [Google Scholar] [CrossRef]
- Chitapanarux, I.; Chitapanarux, T.; Traisathit, P.; Kudumpee, S.; Tharavichitkul, E.; Lorvidhaya, V. Randomized controlled trial of live lactobacillus acidophilus plus bifidobacterium bifidum in prophylaxis of diarrhea during radiotherapy in cervical cancer patients. Radiat. Oncol. 2010, 5, 31. [Google Scholar] [CrossRef]
- Lopez, M.; Li, N.; Kataria, J.; Russell, M.; Neu, J. Live and ultraviolet-inactivated Lactobacillus rhamnosus GG decrease flagellin-induced interleukin-8 production in Caco-2 cells. J. Nutr. 2008, 138, 2264–2268. [Google Scholar] [CrossRef] [PubMed]
- Awaisheh, S.S.; Obeidat, M.M.; Al-Tamimi, H.J.; Assaf, A.M.; EL-Qudah, J.M.; Al-khaza’leh, J.M.; Rahahleh, R.J. In vitro cytotoxic activity of probiotic bacterial cell extracts against Caco-2 and HRT-18 colorectal cancer cells. Milchwissenschaft 2016, 69, 27–31. [Google Scholar]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; Aindelis, G.; Tompoulidou, E.; Lamprianidou, E.E.; Saxami, G.; Ypsilantis, P.; Lampri, E.S.; Simopoulos, C.; et al. Lactobacillus casei Exerts Anti-Proliferative Effects Accompanied by Apoptotic Cell Death and Up-Regulation of TRAIL in Colon Carcinoma Cells. PLoS ONE 2016, 11, e0147960. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Jeon, G.I.; Park, J.S.; Paik, H.D. A Probiotic Strain of Bacillus polyfermenticus Reduces DMH Induced Precancerous Lesions in F344 Male Rat. Biol. Pharm. Bull. 2007, 30, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Bertkova, I.; Hijova, E.; Chmelarova, A.; Mojzisova, G.; Petrasova, D.; Strojny, L.; Bomba, A.; Zitnan, R. The effect of probiotic microorganisms and bioactive compounds on chemically induced carcinogenesis in rats. Neoplasma 2010, 57, 422–428. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, N.K.; Sinha, P.R. Inhibition of 1,2-dimethylhydrazine induced colon genotoxicity in rats by the administration of probiotic curd. Mol. Biol. Rep. 2010, 37, 1373–1376. [Google Scholar] [CrossRef]
- Chang, J.H.; Shim, Y.Y.; Cha, S.K.; Reaney, M.J.T.; Chee, K.M. Effect of Lactobacillus acidophilus KFRI342 on the development of chemically induced precancerous growths in the rat colon. J. Med. Microbiol. 2012, 61, 361–368. [Google Scholar] [CrossRef]
- Lenoir, M.; del Carmen, S.; Cortes-Perez, N.G.; Lozano-Ojalvo, D.; Muñoz-Provencio, D.; Chain, F.; Langella, P.; de Moreno de LeBlanc, A.; LeBlanc, J.G.; Bermúdez-Humarán, L.G. Lactobacillus casei BL23 regulates Treg and Th17 T-cell populations and reduces DMH-associated colorectal cancer. J. Gastroenterol. 2016, 51, 862–873. [Google Scholar] [CrossRef]
- Yu, A.Q.; Li, L. The Potential Role of Probiotics in Cancer Prevention and Treatment. Nutr. Cancer 2016, 68, 535–544. [Google Scholar] [CrossRef]
- Gianotti, L.; Morelli, L.; Galbiati, F.; Rocchetti, S.; Coppola, S.; Beneduce, A.; Gilardini, C.; Zonenschain, D.; Nespoli, A.; Braga, M. A randomized double-blind trial on perioperative administration of probiotics in colorectal cancer patients. World J. Gastroenterol. 2010, 16, 167. [Google Scholar] [CrossRef]
- Liu, Z.H.; Huang, M.J.; Zhang, X.W.; Wang, L.; Huang, N.Q.; Peng, H.; Lan, P.; Peng, J.S.; Yang, Z.; Xia, Y.; et al. The effects of perioperative probiotic treatment on serum zonulin concentration and subsequent postoperative infectious complications after colorectal cancer surgery: A double-center and double-blind randomized clinical trial. Am. J. Clin. Nutr. 2013, 97, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qin, H.; Yang, Z.; Xia, Y.; Liu, W.; Yang, J.; Jiang, Y.; Zhang, H.; Yang, Z.; Wang, Y.; et al. Randomised clinical trial: The effects of perioperative probiotic treatment on barrier function and post-operative infectious complications in colorectal cancer surgery—A double-blind study. Aliment. Pharmacol. Ther. 2011, 33, 50–63. [Google Scholar] [CrossRef]
- Pala, V.; Sieri, S.; Berrino, F.; Vineis, P.; Sacerdote, C.; Palli, D.; Masala, G.; Panico, S.; Mattiello, A.; Tumino, R.; et al. Yogurt consumption and risk of colorectal cancer in the Italian European prospective investigation into cancer and nutrition cohort. Int. J. Cancer 2011, 129, 2712–2719. [Google Scholar] [CrossRef] [PubMed]
- Hatakka, K.; Holma, R.; El-Nezami, H.; Suomalainen, T.; Kuisma, M.; Saxelin, M.; Poussa, T.; Mykkänen, H.; Korpela, R. The influence of Lactobacillus rhamnosus LC705 together with Propionibacterium freudenreichii ssp. shermanii JS on potentially carcinogenic bacterial activity in human colon. Int. J. Food Microbiol. 2008, 128, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Yoshino, K.; Kitajima, M. Possibility of preventing colorectal carcinogenesis with probiotics. Hepatogastroenterology 2010, 57, 1411–1415. [Google Scholar] [PubMed]
- Zhang, J.W.; Du, P.; Gao, J.; Yang, B.R.; Fang, W.J.; Ying, C.M. Preoperative probiotics decrease postoperative infectious complications of colorectal cancer. Am. J. Med. Sci. 2012, 343, 199–205. [Google Scholar] [CrossRef]
- Kotzampassi, K.; Stavrou, G.; Damoraki, G.; Georgitsi, M.; Basdanis, G.; Tsaousi, G.; Giamarellos-Bourboulis, E.J. A Four-Probiotics Regimen Reduces Postoperative Complications After Colorectal Surgery: A Randomized, Double-Blind, Placebo-Controlled Study. World J. Surg. 2015, 39, 2776–2783. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gastrointestinal Tract | Total Colonic Number (log CFU/mL) | Main Types of Microorganisms |
|---|---|---|
| Oral cavity | 108 | Streptococcus, Eubacteria, Capnocytophaga, Veillonella, Fusobacterium, Porphyromonas, Prevotella, Neisseria, Treponema, Lactobacterium, Eikenella, Leptotrichia, Peptostreptococcus, Propionibacterium, Rothia, Scardovia, Parascardovia, Alloscardovia, Candida, Saccharomyces, Penicillium, Scopularis, Aspergillus, Fusarium, Cryptococcus, Alternaria, Geotrichum |
| Oesophagus | 104–6 | Streptococcus, Prevotella, Veillonella |
| Stomach | 102–4 | Helicobacter (species pylori), Lactobacillus, Staphylococcus, Streptococcus, Clostridium, Capnocytophaga, Deinococcus, Veillonella, Escherichia, Bifidobacterium, Prevotella, Caulobacter, Actinobacillus, Corynebacterium, Rothia, Gemella, Leptotrichia, Porphyromonas |
| Duodenum | 103 | Enterococcus, Lactobacillus, Bacteroides, Bifidobacterium, Clostridium, Enterobacteriaceae, yeast |
| Jejunum | 104 | |
| Ileum | 107 | |
| Large intestine | 1010–11 | Enterococcus, Lactobacillus, Bacteroides, Fusobacterium, Bifidobacterium, Clostridium, Enterobacteriaceae, Peptococcus, Peptostreptococcus, Staphylococcus, Ruminococcus, Eubacterium, Streptococcus, Actinomyces, Finegoldia (species magna), Micromonas (species micros), Peptococcus (species niger), Veillonella, Escherichia (species coli), Klebsiella, Proteus, Pseudomonas, Enterococcus (species faecalis), Bacillus |
| Rectum | 1011–12 |
| RESEARCH ON CELL LINES/IN VITRO | |||
| Probiotic Bacteria | Cell Lines | Effects/Mechanisms | Source |
| Lactobacillus rhamnosus GG | Caco–2 | Decreased level of IL–8. | LOPEZ et al. 2008 [80] |
| 40 different probiotic bacteria isolates | Caco–2, HRT–18 Vero cells Using Trypan Blue assays (TBE) and 3–(4, 5–dimethylthiazolyl–2)–2, 5–diphenyltetrazolium bromide (MTT) | Two isolates of Lactobacillus acidophilus LA102 and Lactobacillus casei LC232 showed clear cytotoxic activity. They showed no cytotoxic activity on normal Vero cells. | AWAISHEH et al. 2016 [81] |
| Lactobacillus casei ATCC393 | CT26 (murine colon carcinoma cell lines); HT29 (human colon carcinoma cell lines) Administration of live L. casei and bacterial components to cell lines. | Anti-proliferative activity. Live L. casei induced apoptotic death of CT26 and HT29 cells. | TIPTIRI-KOURPETI et al. 2016 [82] |
| RESEARCH ON ANIMAL MODELS/IN VIVO | |||
| Probiotic Bacteria | Animal Models | Effects/Mechanisms | Source |
| Bacillus polyfermenticus | Five-week-old male F344 rats. Three research groups two of which were administered DMH (one was the control group, the other the study group). | Reduction in the formation of ACF (aberrant crypt foci) of about 50%, in the group with supplementation of B. polyfermenticus. Increased of antioxidant potential. | PARK et al. 2007 [83] |
| Lactobacillus plantarum | Six-month-old male and female Wistar albino rats with induced colon cancer with DMH. | Reduced concentration of bile acid and bacterial enzymes. Increased level of TNF-alpha in the serum and the number of bacteria of the Lactobacillus genus. | BERTKOVA et al. 2010 [84] |
| Lactobacillus acidophilus, Lactobacillus casei and lactis biotype diacetylactis DRC-1 | Rats DMH-induced CRC model. 100 rats were divided into four groups (DMH control group, probiotic curd group, normal curd group, and normal diet group). | Decreasing the incidence, number and size of tumors. Significant reduction in DNA damage. | KUMAR et al. 2010 [85] |
| Lactobacillus rhamnosus 231 (Lr 231) | Male Wistar rats exposed to MNNG (N–Methyl–N’–Nitro–Nitrosoguanidine). | Decreased fecal activity of azoreductase, nitroreductase, GST. Increased GSH. | GOSAI et al. 2011 [40] |
| Lactobacillus acidophilus KFRI342 (isolated from kimchi) | Forty-five male F344 rats with DMH chemically induced premalignant lesions in the colon. | Reduction in ACF, beta-glucuronidase, beta-glucosidase activity, decreased intestinal population of aerobic bacteria and Escherichia coli (in stool samples). | CHANG et al. 2012 [86] |
| Lactobacillus casei BL23 | C57BL/6 mice 1,2–dimethylhydrazine (DMH) was injected subcutaneously every week for 10 weeks. L. casei BL23 was also administered orally in drinking water for up to 10 weeks. | Modulation of host immune response. L. casei BL23 protect mice against DMH-induced colorectal cancer. | LENOIR et al. 2016 [87] |
| Research on Human | |||
|---|---|---|---|
| Prevention | |||
| Probiotic Bacteria | Subjects | Effects/Mechanisms | Source |
| Lactobacillus rhamnosus LC705 and Propionibacterium freudenreichii ssp. shermanii JS | 38 men (between 24 and 55 years old). | Decreased beta-glucosidase activity (by 10%) and urease (by 13%). Increasing the fecal amount of bacteria of the genus Lactobacillus and propionibacteria. | HATAKKA et al. 2008 [93] |
| Lactobacillus gasseri OLL2716 (LG21) | 10 people with colorectal cancer and 20 healthy patients | Increasing the number of bacteria from the genus Lactobacillus, synthesis of isobutyric acid, NK cell activity. Reducing the amount of Clostridium perfringens. | OHARA et al. 2010 [94] |
| Streptococcus thermophilus and Lactobacillus delbruckii subsp. bulgaricus | 45 241 healthy people (14 178 men, 31 063 women) | Reduction in the risk of colorectal cancer correlated with increased consumption of yogurt (especially in men). | PALA et al. 2011 [92] |
| TREATMENT | |||
| Bifidobacterium longum | 60 patients with colorectal cancer undergoing colon resection | Increasing the amount of bacteria of the genus Bifidobacterium, and reducing the amount of bacteria of the genus Escherichia ratio of these bacteria was different to the pre-operative. | ZHANG et al. 2012 [95] |
| Bifidobacterium breve strain Yakult | 42 patients during chemotherapy (19 people were in the study group, 23 in the control group) | Reduction in the incidence of fever and the use of intravenous antibiotics was lower in the study group than in the control group. | WADA et al. 2010 [77] |
| Lactobacillus acidophilus, L. plantarum, Bifidobacterium lactis and Saccharomyces boulardii | 164 patients with colorectal cancer undergoing colorectal surgery | Significantly decreased the risk of postoperative complications. In the probiotic group, a positive correlation was observed between the expression of the SOCS3 gene and the expression of the TNF gene and circulating IL–6. | KOTZAMPASSI et al. 2015 [96] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molska, M.; Reguła, J. Potential Mechanisms of Probiotics Action in the Prevention and Treatment of Colorectal Cancer. Nutrients 2019, 11, 2453. https://doi.org/10.3390/nu11102453
Molska M, Reguła J. Potential Mechanisms of Probiotics Action in the Prevention and Treatment of Colorectal Cancer. Nutrients. 2019; 11(10):2453. https://doi.org/10.3390/nu11102453
Chicago/Turabian StyleMolska, Marta, and Julita Reguła. 2019. "Potential Mechanisms of Probiotics Action in the Prevention and Treatment of Colorectal Cancer" Nutrients 11, no. 10: 2453. https://doi.org/10.3390/nu11102453
APA StyleMolska, M., & Reguła, J. (2019). Potential Mechanisms of Probiotics Action in the Prevention and Treatment of Colorectal Cancer. Nutrients, 11(10), 2453. https://doi.org/10.3390/nu11102453