Omega-3 Polyunsaturated Fatty Acids: Benefits and Endpoints in Sport

 ,
, 
Abstract
1. Introduction
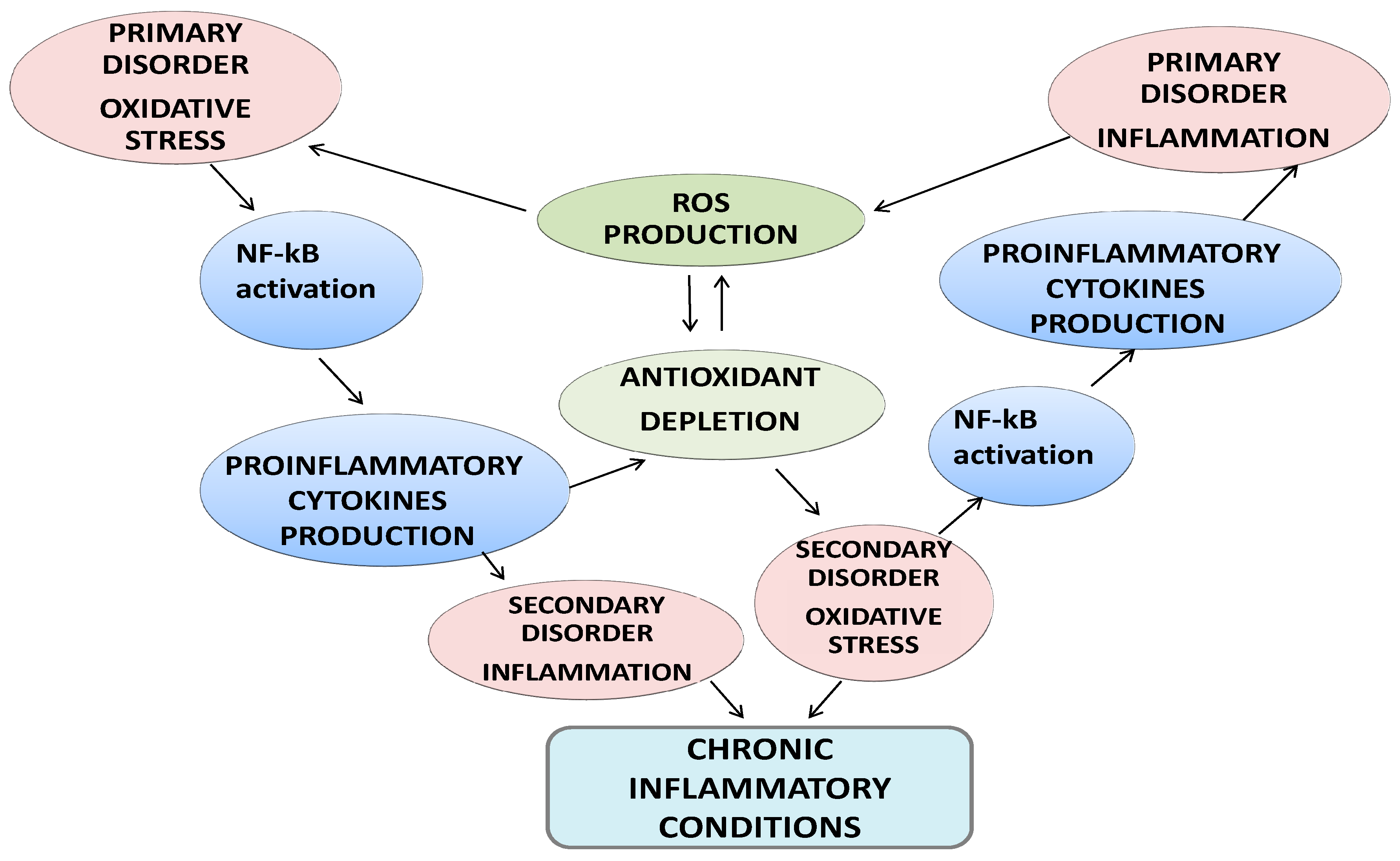
2. Exercise-Induced Oxidative Stress and Inflammation: The Paradox of Intense Sport Exercise
3. N-3 PUFAs and the Health of Skeletal Muscle
4. N-3 PUFAs and the Availability of Energy
5. Immuno-Stimulating Effect of n-3 PUFAs
6. N-3 PUFAs and Cardiovascular Health: Anti-Arrhythmic Potential and Vasodilatation
7. N-3 PUFAs and Inflammatory Diseases
7.1. Role of n-3 PUFAs in Asthma and Exercise-Induced Bronchoconstriction
7.2. Role of n-3 PUFAs in Osteoarthritis and Joint Pain
8. Potential Adverse Effects of n-3 PUFAs
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Simopoulos, A.P. Omega-3 fatty acids and athletics. Curr. Sports Med. Rep. 2007, 6, 230–236. [Google Scholar] [PubMed]
- Ulven, S.M.; Kirkhus, B.; Lamglait, A.; Basu, S.; Elind, E.; Haider, T.; Berge, K.; Vik, H.; Pedersen, J.I. Metabolic Effects of Krill Oil are Essentially Similar to Those of Fish Oil but at Lower Dose of EPA and DHA, in Healthy Volunteers. Lipids 2011, 46, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, A.P.; Sperling, L. Understanding omega-3′s. Am. Heart J. 2006, 151, 564–570. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Lim, D.K.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Fact. 2012, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, N.; Gemello, E.; Gammone, M.A.; DeGirolamo, M.; Ficoneri, C.; Riccioni, G. Fucoxantin: A treasure from the sea. Mar. Drugs 2012, 10, 604–616. [Google Scholar] [CrossRef]
- Lunn, J.; Theobald, H. The health effects of dietary unsaturated fatty acids. Nutr. Bull. 2006, 31, 178–224. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibers in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Pechenkina-Shubina, E.E.; Rozentsvet, O.A. Glycolipids and fatty acids of some seaweeds and marine grasses from the black sea. Phytochemistry 1991, 30, 2279–2283. [Google Scholar] [CrossRef]
- Bocanegra, A.; Bastida, S.; Benedí, J.; Ródenas, S.; Sánchez-Muniz, F.J. Characteristics and nutritional and cardiovascular-health properties of seaweeds. J. Med. Food 2009, 12, 236–258. [Google Scholar] [CrossRef]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signalling 2007, 9, 2277–2293. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Med. Cell Longev. 2016, e5698931. [Google Scholar] [CrossRef]
- Sen, C.K. Oxidants and antioxidants in exercise. J. Appl. Physiol. 1995, 79, 675–686. [Google Scholar] [CrossRef]
- Bogdanovskaya, N.V.; Kotsuruba, A.V.; Golubenko, A.V. Induction of oxidative and nitrosative stress in boys in adapting to physical stress during training and competitive periods. Fiziol. Zh. 2016, 62, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Škrgat, S.; Korošec, P.; Kern, I.; Šilar, M.; Šelb, J.; Fležar, M.; Marčun, R. Systemic and airway oxidative stress in competitive swimmers. Respir. Med. 2018, 137, 129–133. [Google Scholar] [CrossRef]
- Fisher-Wellman, K.; Bloomer, R.J. Acute exercise and oxidative stress: A 30 years history. Dyn. Med. 2009, 8, 1–25. [Google Scholar] [CrossRef]
- Reid, M.B. Nitric oxide, reactive oxygen species, and skeletal muscle contraction. Med. Sci. Sports Exercise 2001, 33, 371–376. [Google Scholar] [CrossRef]
- Niess, A.M.; Simon, P. Response and adaptation of skeletal muscle to exercise: The role of reactive oxygen species. Front. Biosci. 2007, 12, 4826–4838. [Google Scholar] [CrossRef]
- Black, K.E.; Witard, O.C.; Baker, D.; Healey, P.; Lewis, V.; Tavares, F.; Christensen, S.; Pease, T.; Smith, B. Adding omega-3 fatty acids to a protein-based supplement during pre-season training results in reduced muscle soreness and the better maintenance of explosive power in professional Rugby Union players. Eur. J. Sport Sci. 2018, 18, 1357–1367. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Bakke, S.S.; Fredriksson, K.; Boekschoten, M.V.; Fjørkenstad, A.; Koster, G.; Hesselink, M.K.; Kersten, S.; Kase, E.T.; Rustan, A.C.; et al. Metabolic switching of human myotubes is improved by n-3 fatty acids. J. Lipid Res. 2010, 51, 2090–2104. [Google Scholar] [CrossRef] [PubMed]
- Philp, L.K.; Heilbronn, L.K.; Janovska, A.; Wittert, G.A. Dietary enrichment with fish oil prevents high fat-induced metabolic dysfunction in skeletal muscle in mice. PLoS ONE 2015, 10, e0117494. [Google Scholar] [CrossRef] [PubMed]
- You, J.S.; Park, M.N.; Song, W.; Lee, Y.S. Dietary fish oil alleviates soleus atrophy during immobilization in association with Akt signaling to p70s6k and E3 ubiquitin ligases in rats. Appl. Physiol. Nutr. Metab. 2010, 35, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Julliand, S.; Reeds, D.N.; Sinacore, D.R.; Klein, S.; Mittendorfer, B. Fish oil-derived n-3 PUFA therapy increases muscle mass and function in healthy older adults. Am. J. Clin. Nutr. 2015, 102, 115–122. [Google Scholar] [CrossRef]
- Mickleborough, T.D.; Lindley, M.R.; Ionescu, A.A.; Fly, A.D. Protective effect of fish oil supplementation on exercise-induced bronchoconstriction in asthma. Chest 2006, 129, 39–49. [Google Scholar] [CrossRef]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rennie, M.J.; Mittendorfer, B. Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: A randomized controlled trial. Am. J. Clin. Nutr. 2011, 93, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Rodacki, C.L.N.; Rodacki, A.L.F.; Pereira, G.; Naliwaiko, K.; Coelho, I.; Pequito, D.; Fernandes, L.C. Fish-oil supplementation enhances the effects of strength training in elderly women. Am. J. Clin. Nutr. 2012, 95, 428–436. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Importance of the ratio of omega-6/omega-3 essential fatty acids: Evolutionary aspects. World Rev. Nutr. Diet 2003, 92, 1–22. [Google Scholar]
- Simopoulos, A.P. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 494–505. [Google Scholar] [CrossRef]
- Vaid, M.; Singh, T.; Prasad, R.; Katiyar, S.K. Intake of high-fat diet stimulates the risk of ultraviolet radiation-induced skin tumors and malignant progression of papillomas to carcinoma in SKH-1 hairless mice. Toxicol. appl. pharmacol. 2014, 274, 147–155. [Google Scholar] [CrossRef]
- Pilkington, S.M. Impact of EPA ingestion on COX-and LOX-mediated eicosanoid synthesis in skin with and without a pro-inflammatory UVR challenge–Report of a randomised controlled study in humans. Mol. Nutr. Food Res. 2014, 58, 580–590. [Google Scholar] [CrossRef] [PubMed]
- McCusker, M.; Meagen, M.; Grant-Kels, J.M. Healing fats of the skin: The structural and immunologic roles of the ω-6 and ω-3 fatty acids. Clinics Dermatology 2010, 20, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Jeromson, S.; Gallagher, I.J.; Galloway, S.D.; Hamilton, D.L. Omega-3 fatty acids and skeletal muscle health. Mar. Drugs 2015, 13, 6977–7004. [Google Scholar] [CrossRef] [PubMed]
- Wigmore, S.J.; Barber, M.D.; Ross, J.A.; Tisdale, M.J.; Fearon, K.C. Effect of oral eicosapentaenoic acid on weight loss in patients with pancreatic cancer. Nutr. Cancer 2000, 36, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Gingras, A.A.; White, P.J.; Chouinard, P.Y.; Julien, P.; Davis, T.A.; Dombrowski, L. Long-chain n-3 PUFA fatty acids regulate bovine whole-body protein metabolism by promoting muscle insulin signalling to the Akt–mTOR–S6 K1pathway and insulin sensitivity. J. Physiol. 2007, 579, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Kamolrat, T.; Gray, S.R. The effect of eicosapentaenoic and docosahexaenoic acid on protein synthesis and breakdown in murine C2C12 myotubes. Biochem. Biophys. Res. Commun. 2013, 432, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rankin, D.; Rennie, M.J.; Mittendorfer, B. N-3 PUFA polyunsaturated fatty acids augment the muscle protein anabolic response to hyperinsulinaemia- hyperaminoacidaemia in healthy young and middle-aged men and women. Clin. Sci. 2011, 121, 267–278. [Google Scholar] [CrossRef]
- Jouris, K.B.; McDaniel, J.L.; Weiss, E.P. The effect of n-3 PUFA fatty acid supplementation on the inflammatory response to eccentric strength exercise. J. Sports Sci. Med. 2011, 10, 432–438. [Google Scholar] [PubMed]
- Lembke, P.; Capodice, J.; Hebert, K.; Swenson, T. Influence of n-3 omega-3 PUFA (n3) index on performance and wellbeing in young adults after heavy eccentric exercise. J. Sports Sci. Med. 2014, 13, 151–156. [Google Scholar]
- Corder, K.E.; Newsham, K.R.; McDaniel, J.L.; Ezekiel, U.R.; Weiss, E.P. Effects of short-term docosahexaenoic acid supplementation on markers of inflammation after eccentric strength exercise in women. J. Sports Sci. Med. 2016, 15, 176–183. [Google Scholar]
- Tsuchiya, Y.; Yanagimoto, K.; Nakazato, K.; Hayamizu, K.; Ochi, E. Eicosapentaenoic and docosahexaenoic acids-rich fish oil supplementation attenuates strength loss and limited joint range of motion after eccentric contractions: A randomized, double-blind, placebo-controlled, parallel-group trial. Eur. J. Appl. Physiol. 2016, 116, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Tinsley, G.M.; Gann, J.J.; Huber, S.R.; Andre, T.L.; LaBounty, P.M.; Bowden, R.G.; Gordon, P.M.; Grandjean, P.W. Effects of fish oil supplementation on post-resistance exercise muscle soreness. J. Diet Suppl. 2016, 21, 1–12. [Google Scholar]
- Mickleborough, T.D.; Sinex, J.A.; Platt, D.; Chapman, R.F.; Hirt, M. The effects PCSO-524®, a patented marine oil lipid and omega-3 Supplementation with eicosapentaenoic acid-rich mussel (Perna canaliculus), on indirect markers of muscle damage and inflammation after muscle damaging exercise in untrained men: A randomized, placebo controlled trial. J. Int. Soc. Sports Nutr. 2015, 1, 10. [Google Scholar] [CrossRef]
- Vaughan, R.A.; Garcia-Smith, R.; Bisoffi, M.; Conn, C.A.; Trujillo, K.A. Conjugated linoleic acid or omega 3 fatty acids increase mitochondrial biosynthesis and metabolism in skeletal muscle cells. Lipids Health Dis. 2012, 11, 142. [Google Scholar] [CrossRef]
- Gammone, M.A.; Gemello, E.; Riccioni, G.; D’Orazio, N. Marine bioactives and potential application in sports. Mar. Drugs 2014, 12, 2357–2382. [Google Scholar] [CrossRef] [PubMed]
- Patten, G.S.; Abeywardena, M.Y.; McMurchie, E.J.; Jahangiri, A. Dietary fish oil increases acetylcholine- and eicosanoid-induced contractility of isolated rat ileum. J. Nutr. 2002, 132, 2506–2513. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.J.H.; Radonic, P.W.; Wolever, T.M.S.; Wells, G.D. 21-days mammalian omega-3 fatty acid supplementation improves aspects of neuromuscular function and performance in male athletes compared to olive oil placebo. J. Int. Soc. Sports Nutr. 2015, 12, 28. [Google Scholar] [CrossRef]
- DaBoit, M.; Hunter, A.M.; Gray, S.R. Fit with good fat? The role of n-3 polyunsaturated fatty acids on exercise performance. Metab. Clin. Exp. 2017, 66, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Żebrowska, A.; Mizia-Stec, K.; Mizia, M.; Gąsior, Z.; Poprzęcki, S. Omega-3 fatty acids supplementation improves endothelial function and maximal oxygen uptake in endurance-trained athletes. Eur. J. Sport Sci. 2015, 15, 305–314. [Google Scholar] [CrossRef]
- Peoples, G.E.; McLennan, P.L.; Howe, P.R.; Groeller, H.J. Fish oil reduces heart rate and oxygen consumption during exercise. J. Cardiovasc. Pharmacol. 2008, 52, 540–547. [Google Scholar] [CrossRef]
- Macartney, M.J.; Hingley, L.; Brown, M.A.; Peoples, G.E.; McLennan, P.L. Intrinsic heart rate recovery after dynamic exercise is improved with an increased omega-3 index in healthy males. Br. J. Nutr. 2014, 112, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.D.; Burgess, S.; Murphy, K.J.; Howe, P.R. DHA-rich fish oil lowers heart rate during submaximal exercise in elite Australian Rules footballers. J. Sci. Med. Sport 2009, 12, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Peoples, G.E.; McLennan, P.L. Dietary fish oil reduces skeletal muscle oxygen consumption, provides fatigue resistance and improves contractile recovery in the rat in vivo hindlimb. Br. J. Nutr. 2010, 104, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Bishop, N.C.; Stensel, D.J.; Lindley, M.R.; Mastana, S.S.; Nimmo, M.A. The anti-inflammatory effects of exercise: Mechanisms and implications for the prevention and treatment of disease. Nat. Rev. Immunol. 2011, 11, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.; Gabriel, B.; Thies, F.; Gray, S.R. Fish oil supplementation augments post-exercise immune function in young males. Brain Behav. Immunity 2012, 26, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Bishop, N.C. The T cell and NK cell immune response to exercise. Ann Transplant 2005, 10, 43–48. [Google Scholar] [PubMed]
- Starkie, R.L.; Rolland, J.; Febbraio, M.A. Effect of adrenergic blockade on lymphocyte cytokine production at rest and during exercise. Am. J. Physiol. Cell. Physiol. 2001, 281, C1233–C1240. [Google Scholar] [CrossRef] [PubMed]
- Andrade, P.M.M.; Ribeiro, B.G.; Bozza, M.T.; Costa-Rosa, L.F.B.; DoCarmo, M.G.T. Effects of the fish-oil supplementation on the immune and inflammatory responses in elite swimmers. Prost Leuk Essent. Fatty Acids 2007, 77, 139–145. [Google Scholar] [CrossRef]
- Kawabata, F.; Neya, M.; Hamazaki, K.; Watanabe, Y.; Kobayashi, S.; Tsuji, T. Supplementation with eicosapentaenoic acid-rich fish oil improves exercise economy and reduces perceived exertion during submaximal steady-state exercise in normal healthy untrained men. Biosci. Biotechnol. Biochem. 2014, 78, 2081–2088. [Google Scholar] [CrossRef]
- Nieman, D.C.; Mitmesser, S.H. Potential impact of nutrition on immune system recovery from heavy exertion: A metabolomics perspective. Nutrients 2017, 9, 513. [Google Scholar] [CrossRef]
- Lemaitre, R.N.; King, I.B.; Mozaffarian, D.; Kuller, L.H. N-3 Polyunsaturated fatty acids, fatal ischemic heart disease, and non fatal myocardial infarction in older adults: The Cardiovascular Health Study. Am. J. Clin. Nutr. 2003, 77, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, M.; Origasa, H.; Matsuzaki, M.; Japan, M.Y. EPA lipid intervention study (JELIS) investigators. Effects of eicosapentaenoic acid on major coronary events in hypercholesterolaemic patients:a randomized open label, blinded endpoint analysis. Lancet 2007, 369, 1090–1098. [Google Scholar] [CrossRef]
- Albert, C.M.; Campos, H.; Stampfer, M.J.; Ridker, P.M. Blood levels of long-chain n-3 fatty acids and the risk of sudden death. N. Eng. J. Med. 2002, 346, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Favaloro, E.J.; Sanchis-Gomar, F. Sudden cardiac and noncardiac death in sports: Epidemiology, causes, pathogenesis, and prevention. Semin. Thromb. Hemost. 2018, 44, 780–786. [Google Scholar] [PubMed]
- Friedman, A.N.; Yu, Z.; Denski, C.; Tamez, H.; Wenger, J.; Thadhani, R.; Li, Y.; Watkins, B. Fatty acids and other risk factors for sudden cardiac death in patients starting hemodialysis. Am. J. Nephrol. 2013, 38, 12–18. [Google Scholar] [CrossRef]
- Soumia, P.; Sandeep, C.; Jubbin, J.J. A fish a day, keeps the cardiologist away! – A review of the effect of omega-3 fatty acids in the cardiovascular system. Indian J. Endocrinol. Metab. 2013, 17, 422–429. [Google Scholar]
- Billman, G.E.; Kang, J.X.; Leaf, A. Prevention of sudden cardiac death by dietary pure ω-3 polyunsaturated fatty acids in dogs. Circulation 1999, 99, 2452–2457. [Google Scholar] [CrossRef]
- McLennan, P.L.; Bridle, T.M.; Abeywardena, M.Y.; Charnock, J.S. Dietary lipid modulation of ventricular fibrillation threshold in the marmoset monkey. Am. Heart J. 1992, 123, 1555–1561. [Google Scholar] [CrossRef]
- Kang, J.X.; Leaf, A. Antiarrhythmic effects of polyunsaturated fatty acids: Recent studies. Circulation 1996, 94, 1774–1780. [Google Scholar] [CrossRef]
- Xiao, Y.F.; Wright, S.N.; Wang, G.K.; Morgan, J.P.; Leaf, A. Fatty acids suppress voltage-gated Na+ currents in HEK293t cells transfected with the alpha-subunit of the human cardiac Na+ channel. Proc. Natl. Acad. Sci. USA 1998, 95, 2680–2685. [Google Scholar] [CrossRef]
- Nair, S.S.D.; Leitch, J.W.; Falconer, J.; Garg, M.L. Prevention of cardiac arrhythmia by dietary (n-3) polyunsaturated fatty acids and their mechanism of action. J. Nutr. 1997, 127, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kang, J.X.; Leaf, A. Differential effects of various eicosanoids on the production or prevention of arrhythmias in cultured neonatal rat cardiac myocytes. Prostaglandins 1997, 54, 511–530. [Google Scholar] [CrossRef]
- Christensen, J.H.; Korup, E.; Aaroe, J.; Toft, E.; Møller, J.; Rasmussen, K.; Dyerberg, J.; Schmidt, E.B. Fish consumption, n-3 fatty acids in cell membranes, and heart rate variability in survivors of myocardial infarction with left ventricular dysfunction. Am. J. Cardiol. 1997, 79, 1670–1673. [Google Scholar] [CrossRef]
- Christensen, J.H.; Skou, H.A.; Fog, L.; Hansen, V.; Vesterlund, T.; Dyerberg, J.; Toft, E.; Schmidt, E.B. Marine n-3 fatty acids, wine intake, and heart rate variability in patients referred for coronary angiography. Circulation 2001, 103, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Kromhout, D.; Yasuda, S.; Geleijnse, J.M.; Shimokawa, H. Fish oil and omega-3 fatty acids in cardiovascular disease: Do they really work? Eur. Heart J. 2012, 33, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; Vicentini, A.; Riccioni, G.; DeGirolamo, M.; D’Aulerio, A.; D’Orazio. Case report: Food-related atrial fibrillation? The potential role of biogenic amines in “nutria-arrhythmias” genesis. Reports 2019, in press. [Google Scholar]
- Jouven, X.; Charles, M.A.; Desnos, M.; Ducimetiere, P. Circulating non-esterified fatty acid level as a predictive risk factor for sudden death in the population. Circulation 2001, 104, 756–761. [Google Scholar] [CrossRef]
- Calder, P.C. The role of marine omega-3 (n-3) fatty acids in inflammatory processes, atherosclerosis and plaque stability. Mol. Nutr. Food Res. 2012, 56, 1073–1080. [Google Scholar] [CrossRef]
- Raatz, S.K.; Silverstein, J.T.; Jahns, L.; Picklo, M.J. Issues of fish consumption for cardiovascular disease risk reduction. Nutrients 2013, 5, 1081–1097. [Google Scholar] [CrossRef]
- Cottin, S.; Sanders, T.; Hall, W. The differential effects of EPA and DHA on cardiovascular risk factors. Proc. Nutr. Soc. 2011, 70, 215–231. [Google Scholar] [CrossRef]
- Näsman, A.; Irewall, T.; Hållmarker, U.; Lindberg, A.; Stenfors, N. Asthma and Asthma medication are common among recreational athletes participating in endurance sport competitions. Can. Respir. J. 2018, 21, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.; Stang, J.; Horta, L. Two distinct phenotypes of asthma in elite athletes identified by latent class analysis. J. Asthma 2015, 52, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Norqvist, J.; Eriksson, L.; Söderström, L.; Lindberg, A.; Lindberg, A.; Stenfors, N. Self-reported physician-diagnosed asthma among Swedish adolescent, adult and former elite endurance athletes. J. Asthma 2015, 52, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, K.H.; Anderson, S.D.; Bjermer, L. Exercise-induced asthma, respiratory and allergic disorders in elite athletes: Epidemiology, mechanisms and diagnosis: Part I of the report from the Joint Task Force of the European Respiratory Society (ERS) and the European Academy of Allergy and Clinical Immunology (EAACI) in cooperation with GA2LEN. Allergy 2008, 63, 387–403. [Google Scholar] [PubMed]
- Bougault, V.; Boulet, L.P. Airway dysfunction in swimmers. Brit. J. Sports Med. 2012, 46, 402–406. [Google Scholar] [CrossRef]
- Couto, M.; Kurowski, M.; Moreira, A.; Bullens, D.M.A.; Carlsen, K.H.; Delgado, L.; Kowalski, M.L.; Seys, S.F. Mechanisms of exercise-induced bronchoconstriction in athletes: Current perspectives and future challenges. Allergy 2018, 73, 8–16. [Google Scholar] [CrossRef]
- Efthimiou, P.; Kukar, M. Complementary and alternative medicine use in rheumatoid arthritis: Proposed mechanism of action and efficacy of commonly used modalities. Rheumatol. Int. 2009, 30, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Horrobin, D.F. Low prevalences of coronary heart disease (CHD), psoriasis, asthma and rheumatoid arthritis in Eskimos: Are they caused by high dietary intake of eicosapentaenoic acid (EPA), a genetic variation of essential fatty acid (EFA) metabolism or a combination of both? Med. Hypotheses 1987, 22, 421–428. [Google Scholar]
- Schubert, R.; Kitz, R.; Beermann, C. Effect of n-3 polyunsaturated fatty acids in asthma after low-dose allergen challenge. Int. Arch. Allergy Immunol. 2009, 148, 321–329. [Google Scholar] [CrossRef]
- Tecklenburg-Lund, S.; Mickleborough, T.D.; Turner, L.A.; Fly, A.D.; Stager, J.M.; Montgomery, G.S. Randomized controlled trial of fish oil and montelukast and their combination on airway inflammation and hyperpnea-induced bronchoconstriction. PLoS ONE 2010, 5, e13487. [Google Scholar] [CrossRef]
- Biltagi, M.A.; Baset, A.A.; Bassiouny, M.; Kasrawi, M.A.; Attia, M. Omega-3 fatty acids, vitamin C and Zn supplementation in asthmatic children: A randomized self- controlled study. Acta Paediatrics 2009, 98, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.D.; Holzer, K. Exercise-induced asthma: Is it the right diagnosis in elite athletes? J. Allergy Clin. Immunol. 2000, 106, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Vannini, F.; Spalding, T.; Andriolo, L.; Berruto, M.; Denti, M.; Espregueira-Mendes, J.; Menetrey, J.; Peretti, G.M.; Seil, R.; Filardo, G. Sport and early osteoarthritis: The role of sport in aetiology, progression and treatment of knee osteoarthritis. Knee Surg Sports Traumatol. Arthrosc. 2016, 24, 1786–1796. [Google Scholar] [CrossRef]
- Otsuki, S.; Nakajima, M.; Okamoto, Y.; Oda, S.; Hoshiyama, Y.; Iida, G.; Neo, M. Correlation between varus knee malalignment and patellofemoral osteoarthritis. Knee Surg. Sports Traumatol. Arthrosc. 2016, 24, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Buckwalter, J.A. Sports, joint injury, and post-traumatic osteoarthritis. J. Orthop Sports Phys. Ther. 2003, 33, 578–588. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, N.; Gammone, M.A.; Gemello, E.; DeGirolamo, M.; Cusenza, S.; Riccioni, G. Marine bioactives: Pharmacological properties and potential applications against inflammatory diseases. Mar. Drugs 2012, 10, 812–833. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef]
- Leslie, C.A.; Gonnerman, W.A.; Ullman, M.D. Dietary fish oil modulates macrophage fatty acids and decreases arthritis susceptibility in mice. J. Exp. Med. 1985, 162, 1336–1339. [Google Scholar] [CrossRef]
- Zawadzki, M.; Janosch, C.; Szechinski, J. Perna canaliculus lipid complex PCSO-524™ demonstrated pain relief for osteoarthritis patients benchmarked against fish oil, a randomized trial, without placebo control. Mar. Drugs 2013, 11, 1920–1935. [Google Scholar] [CrossRef]
- Wensing, A.G.; Mensink, R.P.; Hornstra, G. Effects of dietary n-3 polyunsaturated fatty acids from plant and marine origin on platelet aggregation in healthy elderly subjects. Br. J. Nutr. 1999, 82, 183–191. [Google Scholar]
- Wander, R.C.; Hall, J.A.; Gradin, J.L. The ratio of dietary (n-6) to (n-3) fatty acids influences immune system function, eicosanoid metabolism, lipid peroxidation and vitamin E status in aged dogs. J. Nutr. 1997, 127, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; D’Orazio, N. Anti-obesity activity of the marine carotenoid fucoxanthin. Mar. Drugs 2015, 13, 2196–2214. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Carotenoids: Potential allies of cardiovascular health? Food Nutr. Res. 2015, 59, 26762. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; Riccioni, G.; Galvano, F.; D’Orazio, N. Novel Therapeutic Strategies Against Cancer: Marine-derived Drugs May Be the Answer? Anticancer Agents Med. Chem. 2016, 16, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.H.; Stein, E.A.; Bays, H.E. Efficacy and tolerability of adding prescription omega-3 fatty acids 4 g/d to simvastatin 40mg/d in hypertriglyceridemic patients: An 8-week, randomized, double-blind, placebo-controlled study. Clin. Ther. 2007, 29, 1354–1367. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Silveira, S.; Dujovne, C.A. The combined effects of N-3 fatty acids and aspirin on hemostatic parameters in man. Thromb. Res. 1990, 57, 517–526. [Google Scholar] [CrossRef]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Marine carotenoids against oxidative stress: Effects on human health. Mar. Drugs 2015, 13, 6226–6246. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, G.; Gammone, M.A.; Tettamanti, G.; Bergante, S.; Pluchinotta, F.R.; D’Orazio, N. Resveratrol and anti-atherogenic effects. Int. J. Food Sci. Nutr. 2015, 66, 603–610. [Google Scholar] [CrossRef]
- Gammone, M.A.; Efthymakis, K.; Pluchinotta, F.R.; Tettamanti, G.; Bergante, S.; Riccioni, G.; D’Orazio, N. Impact of chocolate on the cardiovascular health. Front. Biosci. (Landmark Ed.) 2018, 23, 852–864. [Google Scholar] [CrossRef]
- Gammone, M.A.; Tettamanti, G.; Bergante, S.; Pulchinotta, F.R.; D’Orazio, N. Prevention of cardiovascular diseases with carotenoids. Front. Biosci. (Schol. Ed.) 2017, 9, 165–171. [Google Scholar] [CrossRef]
- Riccioni, G.; Gammone, M.A.; Currenti, W.; D’Orazio, N. Effectiveness and safety of dietetic supplementation of a new nutraceutical on lipid profile and serum inflammation biomarkers in hypercholesterolemic patients. Molecules 2018, 23, 1168. [Google Scholar] [CrossRef] [PubMed]
- Shei, R.J.; Lindley, M.R.; Mickleborough, T.D. Omega-3 polyunsaturated fatty acids in the optimization of physical performance. Mil. Med. 2014, 179, 144–156. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Protocol | Key Results |
|---|---|
| 551 mg eicosapentaenoic acid (EPA) and 551 mg docosahexaenoic acid (DHA) twice daily, during five weeks of pre-season rugby training | Reduced fatigue in countermovement jump tests [20] |
| 24-h exposure with 100 microM EPA in human myotubes | Augmented adaptability and upregulation of specific genes implicated in fatty acid beta-oxidation with global improvement in muscle metabolic flexibility [21] |
| Four-week supplementation with n-3 PUFAs 1.1 g per day | Significant increase in maximal oxygen uptake (VO2-max) and in endothelial function [22] |
| 14-days diet enriched with 5% cod liver oil followed by 14 days immobilization | Reduced myosin heavy chain loss during 14 days of hind limb immobilization [23] |
| Six-months supplementation with 1.8 g EPA, 1.5 g DHA daily | Increased hand grip and muscle strength [24] |
| Three-week supplementation with 3.2 g of EPA and 2.0 g of DHA | Reduced eicosanoids and pro-inflammatory cytokines concentration in the sputum of asthmatic athletes [25] |
| Six-months supplementation with 3.36 g/day of n-3 PUFAs | Increased muscle mass and strength in older people [24] |
| Eight-weeks supplementation with 1.86 g EPA, 1.5 g DHA daily | Augmented muscle protein synthesis, enhanced rapamycin (mTOR)-ribosomal protein S6 kinase beta-1 (p70s6k1) signaling after hyperaminoacidemic-hyperinsulinemic clamp [26] |
| Supplementation with 0.4 g EPA, 0.3 g DHA (60 days pre-training and 90 days during training) | Potential training increase in peak torque and rate of torque development (Knee extensor, flexor, plantar, and dorsiflexor) [27] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gammone, M.A.; Riccioni, G.; Parrinello, G.; D’Orazio, N. Omega-3 Polyunsaturated Fatty Acids: Benefits and Endpoints in Sport. Nutrients 2019, 11, 46. https://doi.org/10.3390/nu11010046
Gammone MA, Riccioni G, Parrinello G, D’Orazio N. Omega-3 Polyunsaturated Fatty Acids: Benefits and Endpoints in Sport. Nutrients. 2019; 11(1):46. https://doi.org/10.3390/nu11010046
Chicago/Turabian StyleGammone, Maria Alessandra, Graziano Riccioni, Gaspare Parrinello, and Nicolantonio D’Orazio. 2019. "Omega-3 Polyunsaturated Fatty Acids: Benefits and Endpoints in Sport" Nutrients 11, no. 1: 46. https://doi.org/10.3390/nu11010046
APA StyleGammone, M. A., Riccioni, G., Parrinello, G., & D’Orazio, N. (2019). Omega-3 Polyunsaturated Fatty Acids: Benefits and Endpoints in Sport. Nutrients, 11(1), 46. https://doi.org/10.3390/nu11010046