Metabolomics and Microbiomes as Potential Tools to Evaluate the Effects of the Mediterranean Diet

 and
and
Abstract
1. Introduction
2. Metabolomics and the Mediterranean Diet—The Present Status
2.1. Metabolomics Approach as an Assessment of Adherence to the Mediterranean Diet
2.2. Metabolomics, the Mediterranean Diet, and the Association with Health
3. Microbiome and the Mediterranean Diet—The Present Status
3.1. Mediterranean Diet Effects on General Microbiome Composition
3.2. Microbiome, the Mediterranean Diet, and the Association with Health
4. Conclusions, Implications, and Future Directions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mitrou, P.N.; Kipnis, V.; Thiébaut, A.C.M.; Reedy, J.; Subar, A.F.; Wirfält, E.; Flood, A.; Mouw, T.; Hollenbeck, A.R.; Leitzmann, M.F.; et al. Mediterranean dietary pattern and prediction of all-cause mortality in a US population: Results from the NIH-AARP Diet and Health Study. Arch. Intern. Med. 2007, 167, 2461–2468. [Google Scholar] [CrossRef]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; De, S.F.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics-2017 Update: A Report From the American Heart Association. Circ. Circ. 2017, 135, e146. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Marventano, S.; Yang, J.; Micek, A.; Pajak, A.; Scalfi, L.; Galvano, F.; Kales, S.N. A comprehensive meta-analysis on evidence of Mediterranean diet and cardiovascular disease: Are individual components equal? Crit. Rev. Food Sci. Nutr. 2017, 57, 3218–3232. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Missbach, B.; König, J.; Hoffmann, G. Adherence to a Mediterranean diet and risk of diabetes: A systematic review and meta-analysis. Public Health Nutr. 2015, 18, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Schwedhelm, C.; Galbete, C.; Hoffmann, G. Adherence to Mediterranean Diet and Risk of Cancer: An Updated Systematic Review and Meta-Analysis. Nutrients 2017, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Sofi, F.; Abbate, R.; Gensini, G.F.; Casini, A. Accruing evidence on benefits of adherence to the Mediterranean diet on health: An updated systematic review and meta-analysis. Am. J. Clin. Nutr. 2010, 92, 1189–1196. [Google Scholar] [CrossRef]
- USDA. Scientific Report of the 2015 Dietary Guidelines Advisory Committee; USDA: Washington, DC, USA, 2015.
- Shen, J.; Wilmot, K.A.; Ghasemzadeh, N.; Molloy, D.L.; Burkman, G.; Mekonnen, G.; Gongora, M.C.; Quyyumi, A.A.; Sperling, L.S. Mediterranean Dietary Patterns and Cardiovascular Health. Annu. Rev. Nutr. 2015, 35, 425–449. [Google Scholar] [CrossRef]
- Korre, M.; Sotos-Prieto, M.; Kales, S.N. Survival Mediterranean Style: Lifestyle Changes to Improve the Health of the US Fire Service. Front. Public Health 2017, 5, 331. [Google Scholar] [CrossRef]
- Praticò, G.; Gao, Q.; Scalbert, A.; Vergères, G.; Kolehmainen, M.; Manach, C.; Brennan, L.; Pedapati, S.H.; Afman, L.A.; Wishart, D.S.; et al. Guidelines for Biomarker of Food Intake Reviews (BFIRev): How to conduct an extensive literature search for biomarker of food intake discovery. Genes Nutr. 2018, 13, 3. [Google Scholar] [CrossRef]
- Thompson, F.E.; Subar, A.F.; Loria, C.M.; Reedy, J.L.; Baranowski, T. Need for technological innovation in dietary assessment. J. Am. Diet. Assoc. 2010, 110, 48–51. [Google Scholar] [CrossRef]
- Frobisher, C.; Maxwell, S.M. The estimation of food portion sizes: A comparison between using descriptions of portion sizes and a photographic food atlas by children and adults. J. Hum. Nutr. Diet. Off. J. Br. Diet. Assoc. 2003, 16, 181–188. [Google Scholar] [CrossRef]
- Jenab, M.; Slimani, N.; Bictash, M.; Ferrari, P.; Bingham, S.A. Biomarkers in nutritional epidemiology: Applications, needs and new horizons. Hum. Genet. 2009, 125, 507–525. [Google Scholar] [CrossRef]
- O’Sullivan, A.; Gibney, M.J.; Brennan, L. Dietary intake patterns are reflected in metabolomic profiles: Potential role in dietary assessment studies. Am. J. Clin. Nutr. 2011, 93, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Metabolomics: Applications to food science and nutrition research. Trends Food Sci. Technol. 2008, 19, 482–493. [Google Scholar] [CrossRef]
- Panek, M.; Paljetak, H.Č.; Barešić, A.; Perić, M.; Matijašić, M.; Lojkić, I.; Bender, D.V.; Krznarić, Ž.; Verbanac, D. Methodology challenges in studying human gut microbiota—Effects of collection, storage, DNA extraction and next generation sequencing technologies. Sci. Rep. 2018, 8, 5143. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, W.; Downer, P.; Lever, M.; Chambers, S.T.; George, P.M. Effects of orange juice and proline betaine on glycine betaine and homocysteine in healthy male subjects. Eur. J. Nutr. 2007, 46, 446–452. [Google Scholar] [CrossRef]
- Pujos-Guillot, E.; Hubert, J.; Martin, J.-F.; Lyan, B.; Quintana, M.; Claude, S.; Chabanas, B.; Rothwell, J.A.; Bennetau-Pelissero, C.; Scalbert, A.; et al. Mass spectrometry-based metabolomics for the discovery of biomarkers of fruit and vegetable intake: Citrus fruit as a case study. J. Proteome Res. 2013, 12, 1645–1659. [Google Scholar] [CrossRef]
- Heinzmann, S.S.; Brown, I.J.; Chan, Q.; Bictash, M.; Dumas, M.-E.; Kochhar, S.; Stamler, J.; Holmes, E.; Elliott, P.; Nicholson, J.K. Metabolic profiling strategy for discovery of nutritional biomarkers: Proline betaine as a marker of citrus consumption. Am. J. Clin. Nutr. 2010, 92, 436–443. [Google Scholar] [CrossRef]
- May, D.H.; Navarro, S.L.; Ruczinski, I.; Hogan, J.; Ogata, Y.; Schwarz, Y.; Levy, L.; Holzman, T.; McIntosh, M.W.; Lampe, J.W. Metabolomic Profiling of Urine: Response to a Randomized, Controlled Feeding Study of Select Fruits and Vegetables, and Application to an Observational Study. Br. J. Nutr. 2013, 110, 1760–1770. [Google Scholar] [CrossRef]
- Lloyd, A.J.; Beckmann, M.; Favé, G.; Mathers, J.C.; Draper, J. Proline betaine and its biotransformation products in fasting urine samples are potential biomarkers of habitual citrus fruit consumption. Br. J. Nutr. 2011, 106, 812–824. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Gibbons, H.; O’Gorman, A. An Overview of the Role of Metabolomics in the Identification of Dietary Biomarkers. Curr. Nutr. Rep. 2015, 4, 304–312. [Google Scholar] [CrossRef]
- Edmands, W.M.B.; Beckonert, O.P.; Stella, C.; Campbell, A.; Lake, B.G.; Lindon, J.C.; Holmes, E.; Gooderham, N.J. Identification of human urinary biomarkers of cruciferous vegetable consumption by metabonomic profiling. J. Proteome Res. 2011, 10, 4513–4521. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.-B.S.; Reinbach, H.C.; Rinnan, Å.; Barri, T.; Mithril, C.; Dragsted, L.O. Discovery of exposure markers in urine for Brassica-containing meals served with different protein sources by UPLC-qTOF-MS untargeted metabolomics. Metabolomics 2013, 9, 984–997. [Google Scholar] [CrossRef]
- Guertin, K.A.; Moore, S.C.; Sampson, J.N.; Huang, W.-Y.; Xiao, Q.; Stolzenberg-Solomon, R.Z.; Sinha, R.; Cross, A.J. Metabolomics in nutritional epidemiology: Identifying metabolites associated with diet and quantifying their potential to uncover diet-disease relations in populations. Am. J. Clin. Nutr. 2014, 100, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Aubertin-Leheudre, M.; Koskela, A.; Samaletdin, A.; Adlercreutz, H. Plasma alkylresorcinol metabolites as potential biomarkers of whole-grain wheat and rye cereal fibre intakes in women. Br. J. Nutr. 2010, 103, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Landberg, R.; Townsend, M.K.; Neelakantan, N.; Sun, Q.; Sampson, L.; Spiegelman, D.; van Dam, R.M. Alkylresorcinol metabolite concentrations in spot urine samples correlated with whole grain and cereal fiber intake but showed low to modest reproducibility over one to three years in U.S. women. J. Nutr. 2012, 142, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Guyman, L.A.; Adlercreutz, H.; Koskela, A.; Li, L.; Beresford, S.A.A.; Lampe, J.W. Urinary 3-(3,5-dihydroxyphenyl)-1-propanoic acid, an alkylresorcinol metabolite, is a potential biomarker of whole-grain intake in a U.S. population. J. Nutr. 2008, 138, 1957–1962. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Bhupathiraju, S.N.; Hu, F.B. Use of Metabolomics in Improving Assessment of Dietary Intake. Clin. Chem. 2018, 64, 82–98. [Google Scholar] [CrossRef]
- Slemc, L.; Kunej, T. Transcription factor HIF1A: Downstream targets, associated pathways, polymorphic hypoxia response element (HRE) sites, and initiative for standardization of reporting in scientific literature. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 14851–14861. [Google Scholar] [CrossRef]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104 (Suppl. S2), S1–S63. [Google Scholar] [CrossRef] [PubMed]
- Guarner, F.; Malagelada, J.-R. Gut flora in health and disease. Lancet Lond. Engl. 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Human Nutrition, the Gut Microbiome and the Immune System|Nature. Available online: https://www-nature-com.ezp-prod1.hul.harvard.edu/articles/nature10213 (accessed on 28 November 2018).
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Leffler, D.A.; Lamont, J.T. Clostridium difficile Infection. N. Engl. J. Med. 2015, 373, 287–288. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C. Thinking from the Gut. Nature 2015, 518, S12. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Tindall, A.M.; Petersen, K.S.; Kris-Etherton, P.M. Dietary Patterns Affect the Gut Microbiome-The Link to Risk of Cardiometabolic Diseases. J. Nutr. 2018, 148, 1402–1407. [Google Scholar] [CrossRef]
- Castillo-Peinado, L.S.; Luque de Castro, M.D. Present and foreseeable future of metabolomics in forensic analysis. Anal. Chim. Acta 2016, 925, 1–15. [Google Scholar] [CrossRef]
- Xia, J.; Broadhurst, D.I.; Wilson, M.; Wishart, D.S. Translational biomarker discovery in clinical metabolomics: An introductory tutorial. Metabolomics 2013, 9, 280–299. [Google Scholar] [CrossRef]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2008, 5, 3. [Google Scholar] [CrossRef]
- Dam, R.M.V.; Li, T.; Spiegelman, D.; Franco, O.H.; Hu, F.B. Combined impact of lifestyle factors on mortality: Prospective cohort study in US women. BMJ 2008, 337, a1440. [Google Scholar] [CrossRef] [PubMed]
- Fitó, M.; Konstantinidou, V. Nutritional Genomics and the Mediterranean Diet’s Effects on Human Cardiovascular Health. Nutrients 2016, 8, 218. [Google Scholar] [CrossRef] [PubMed]
- Del Chierico, F.; Vernocchi, P.; Dallapiccola, B.; Putignani, L. Mediterranean Diet and Health: Food Effects on Gut Microbiota and Disease Control. Int. J. Mol. Sci. 2014, 15, 11678–11699. [Google Scholar] [CrossRef] [PubMed]
- Rezzi, S.; Ramadan, Z.; Fay, L.B.; Kochhar, S. Nutritional metabonomics: Applications and perspectives. J. Proteome Res. 2007, 6, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Martínez-González, M.Á.; Ruiz-Canela, M.; Hruby, A.; Liang, L.; Trichopoulou, A.; Hu, F.B. Intervention Trials with the Mediterranean Diet in Cardiovascular Prevention: Understanding Potential Mechanisms through Metabolomic Profiling. J. Nutr. 2016, 146, 913S–919S. [Google Scholar] [CrossRef]
- Esko, T.; Hirschhorn, J.N.; Feldman, H.A.; Hsu, Y.-H.H.; Deik, A.A.; Clish, C.B.; Ebbeling, C.B.; Ludwig, D.S. Metabolomic profiles as reliable biomarkers of dietary composition. Am. J. Clin. Nutr. 2017, 105, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, H.; Carr, E.; McNulty, B.A.; Nugent, A.P.; Walton, J.; Flynn, A.; Gibney, M.J.; Brennan, L. Metabolomic-based identification of clusters that reflect dietary patterns. Mol. Nutr. Food Res. 2017, 61, 1601050. [Google Scholar] [CrossRef] [PubMed]
- Playdon, M.C.; Moore, S.C.; Derkach, A.; Reedy, J.; Subar, A.F.; Sampson, J.N.; Albanes, D.; Gu, F.; Kontto, J.; Lassale, C.; et al. Identifying biomarkers of dietary patterns by using metabolomics. Am. J. Clin. Nutr. 2017, 105, 450–465. [Google Scholar] [CrossRef] [PubMed]
- Bondia-Pons, I.; Martinez, J.A.; de la Iglesia, R.; Lopez-Legarrea, P.; Poutanen, K.; Hanhineva, K.; de los Ángeles Zulet, M. Effects of short- and long-term Mediterranean-based dietary treatment on plasma LC-QTOF/MS metabolic profiling of subjects with metabolic syndrome features: The Metabolic Syndrome Reduction in Navarra (RESMENA) randomized controlled trial. Mol. Nutr. Food Res. 2015, 59, 711–728. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Fresno, R.; Llorach, R.; Urpi-Sarda, M.; Lupianez-Barbero, A.; Estruch, R.; Corella, D.; Fitó, M.; Arós, F.; Ruiz-Canela, M.; Salas-Salvadó, J.; et al. Metabolomic Pattern Analysis after Mediterranean Diet Intervention in a Nondiabetic Population: A 1- and 3-Year Follow-up in the PREDIMED Study. J. Proteome Res. 2015, 14, 531–540. [Google Scholar] [CrossRef]
- Rebholz, C.M.; Lichtenstein, A.H.; Zheng, Z.; Appel, L.J.; Coresh, J. Serum untargeted metabolomic profile of the Dietary Approaches to Stop Hypertension (DASH) dietary pattern. Am. J. Clin. Nutr. 2018, 108, 243–255. [Google Scholar] [CrossRef] [PubMed]
- González-Guardia, L.; Yubero-Serrano, E.M.; Delgado-Lista, J.; Perez-Martinez, P.; Garcia-Rios, A.; Marin, C.; Camargo, A.; Delgado-Casado, N.; Roche, H.M.; Perez-Jimenez, F.; et al. Effects of the Mediterranean Diet Supplemented with Coenzyme Q10 on Metabolomic Profiles in Elderly Men and Women. J. Gerontol. Ser. A 2015, 70, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Kakkoura, M.G.; Sokratous, K.; Demetriou, C.A.; Loizidou, M.A.; Loucaides, G.; Kakouri, E.; Hadjisavvas, A.; Kyriacou, K. Mediterranean diet-gene interactions: A targeted metabolomics study in Greek-Cypriot women. Mol. Nutr. Food Res. 2017, 61, 1600558. [Google Scholar] [CrossRef]
- Kakkoura, M.G.; Demetriou, C.A.; Loizidou, M.A.; Loucaides, G.; Neophytou, I.; Marcou, Y.; Hadjisavvas, A.; Kyriacou, K. Single-nucleotide polymorphisms in one-carbon metabolism genes, Mediterranean diet and breast cancer risk: A case-control study in the Greek-Cypriot female population. Genes Nutr. 2015, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- Kakkoura, M.G.; Demetriou, C.A.; Loizidou, M.A.; Loucaides, G.; Neophytou, I.; Malas, S.; Kyriacou, K.; Hadjisavvas, A. MnSOD and CAT polymorphisms modulate the effect of the Mediterranean diet on breast cancer risk among Greek-Cypriot women. Eur. J. Nutr. 2016, 55, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Kakkoura, M.G.; Loizidou, M.A.; Demetriou, C.A.; Loucaides, G.; Daniel, M.; Kyriacou, K.; Hadjisavvas, A. The synergistic effect between the Mediterranean diet and GSTP1 or NAT2 SNPs decreases breast cancer risk in Greek-Cypriot women. Eur. J. Nutr. 2017, 56, 545–555. [Google Scholar] [CrossRef]
- Almanza-Aguilera, E.; Urpi-Sarda, M.; Llorach, R.; Vázquez-Fresno, R.; Garcia-Aloy, M.; Carmona, F.; Sanchez, A.; Madrid-Gambin, F.; Estruch, R.; Corella, D.; et al. Microbial metabolites are associated with a high adherence to a Mediterranean dietary pattern using a 1H-NMR-based untargeted metabolomics approach. J. Nutr. Biochem. 2017, 48, 36–43. [Google Scholar] [CrossRef]
- Hadjisavvas, A.; Loizidou, M.A.; Middleton, N.; Michael, T.; Papachristoforou, R.; Kakouri, E.; Daniel, M.; Papadopoulos, P.; Malas, S.; Marcou, Y.; et al. An investigation of breast cancer risk factors in Cyprus: A case control study. BMC Cancer 2010, 10, 447. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef]
- Ruiz-Canela, M.; Hruby, A.; Clish, C.B.; Liang, L.; Martínez-González, M.A.; Hu, F.B. Comprehensive Metabolomic Profiling and Incident Cardiovascular Disease: A Systematic Review. J. Am. Heart Assoc. Cardiovasc. Cerebrovasc. Dis. 2017, 6, e005705. [Google Scholar] [CrossRef] [PubMed]
- Loo, R.L.; Zou, X.; Appel, L.J.; Nicholson, J.K.; Holmes, E. Characterization of metabolic responses to healthy diets and association with blood pressure: Application to the Optimal Macronutrient Intake Trial for Heart Health (OmniHeart), a randomized controlled study. Am. J. Clin. Nutr. 2018, 107, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Khakimov, B.; Poulsen, S.K.; Savorani, F.; Acar, E.; Gürdeniz, G.; Larsen, T.M.; Astrup, A.; Dragsted, L.O.; Engelsen, S.B. New Nordic Diet versus Average Danish Diet: A Randomized Controlled Trial Revealed Healthy Long-Term Effects of the New Nordic Diet by GC-MS Blood Plasma Metabolomics. J. Proteome Res. 2016, 15, 1939–1954. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Zheng, Y.; Ruiz-Canela, M.; Hruby, A.; Martínez-González, M.A.; Clish, C.B.; Corella, D.; Estruch, R.; Ros, E.; Fitó, M. Plasma acylcarnitines and risk of cardiovascular disease: Effect of Mediterranean diet interventions–3. Am. J. Clin. Nutr. 2016, 103, 1408–1416. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Hu, F.B.; Ruiz-Canela, M.; Bulló, M.; Toledo, E.; Wang, D.D.; Corella, D.; Gómez-Gracia, E.; Fiol, M.; Estruch, R. Plasma Metabolites from Choline Pathway and Risk of Cardiovascular Disease in the PREDIMED (Prevention with Mediterranean Diet) Study. J. Am. Heart Assoc. 2017, 6, e006524. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.; Ruiz-Canela, M.; Guasch-Ferré, M.; Zheng, Y.; Toledo, E.; Clish, C.B.; Salas-Salvadó, J.; Liang, L.; Wang, D.D.; Corella, D.; et al. Increases in Plasma Tryptophan Are Inversely Associated with Incident Cardiovascular Disease in the Prevención con Dieta Mediterránea (PREDIMED) Study. J. Nutr. 2017, 147, 314–332. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Toledo, E.; Hruby, A.; Rosner, B.A.; Willett, W.C.; Sun, Q.; Razquin, C.; Zheng, Y.; Ruiz-Canela, M.; Guasch-Ferré, M.; et al. Plasma Ceramides, Mediterranean Diet, and Incident Cardiovascular Disease in the PREDIMED Trial (Prevención con Dieta Mediterránea) Clinical Perspective. Circulation 2017, 135, 2028–2040. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Hu, F.B.; Ruiz-Canela, M.; Clish, C.B.; Dennis, C.; Salas-Salvado, J.; Hruby, A.; Liang, L.; Toledo, E.; Corella, D.; et al. Metabolites of Glutamate Metabolism Are Associated with Incident Cardiovascular Events in the PREDIMED PREvención con DIeta MEDiterránea (PREDIMED) Trial. J. Am. Heart Assoc. Cardiovasc. Cerebrovasc. Dis. 2016, 5, e00375. [Google Scholar] [CrossRef]
- Razquin, C.; Liang, L.; Toledo, E.; Clish, C.B.; Ruiz-Canela, M.; Zheng, Y.; Wang, D.D.; Corella, D.; Castaner, O.; Ros, E.; et al. Plasma lipidome patterns associated with cardiovascular risk in the PREDIMED trial: A case-cohort study. Int. J. Cardiol. 2018, 253, 126–132. [Google Scholar] [CrossRef]
- Ruiz-Canela, M.; Guasch-Ferré, M.; Toledo, E.; Clish, C.B.; Razquin, C.; Liang, L.; Wang, D.D.; Corella, D.; Estruch, R.; Hernáez, Á.; et al. Plasma branched chain/aromatic amino acids, enriched Mediterranean diet and risk of type 2 diabetes: Case-cohort study within the PREDIMED Trial. Diabetologia 2018, 61, 1560–1571. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.; Ruiz-Canela, M.; Razquin, C.; Guasch-Ferre, M.; Toledo, E.; Wang, D.D.; Papandreou, C.; Dennis, C.; Clish, C.; Liang, L.; et al. Changes in Arginine are Inversely Associated with Type 2 Diabetes: A Case-Cohort Study in the PREDIMED Trial. Diabetes Obes. Metab. 2019, 21, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Razquin, C.; Toledo, E.; Clish, C.B.; Ruiz-Canela, M.; Dennis, C.; Corella, D.; Papandreou, C.; Ros, E.; Estruch, R.; Guasch-Ferré, M.; et al. Plasma Lipidomic Profiling and Risk of Type 2 Diabetes in the PREDIMED Trial. Diabetes Care 2018, 41, 2617–2624. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.; Papandreou, C.; Ruiz-Canela, M.; Guasch-Ferre, M.; Clish, C.B.; Dennis, C.; Liang, L.; Corella, D.; Fitó, M.; Razquin, C.; et al. Association of Tryptophan Metabolites with Incident Type 2 Diabetes in the PREDIMED Trial: A Case-Cohort Study. Clin. Chem. 2018, 64, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, C.; Bulló, M.; Zheng, Y.; Ruiz-Canela, M.; Yu, E.; Guasch-Ferré, M.; Toledo, E.; Clish, C.; Corella, D.; Estruch, R.; et al. Plasma trimethylamine-N-oxide and related metabolites are associated with type 2 diabetes risk in the Prevención con Dieta Mediterránea (PREDIMED) trial. Am. J. Clin. Nutr. 2018, 108, 163–173. [Google Scholar] [PubMed]
- Toledo, E.; Wang, D.D.; Ruiz-Canela, M.; Clish, C.B.; Razquin, C.; Zheng, Y.; Guasch-Ferré, M.; Hruby, A.; Corella, D.; Gómez-Gracia, E. Plasma lipidomic profiles and cardiovascular events in a randomized intervention trial with the Mediterranean diet. Am. J. Clin. Nutr. 2017, 106, 973–983. [Google Scholar] [PubMed]
- Ruiz-Canela, M.; Toledo, E.; Clish, C.B.; Hruby, A.; Liang, L.; Salas-Salvadó, J.; Razquin, C.; Corella, D.; Estruch, R.; Ros, E.; et al. Plasma Branched-Chain Amino Acids and Incident Cardiovascular Disease in the PREDIMED Trial. Clin. Chem. 2016, 62, 582–592. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Ruiz-Canela, M.; Li, J.; Zheng, Y.; Bulló, M.; Wang, D.D.; Toledo, E.; Clish, C.; Corella, D.; Estruch, R.; et al. Plasma acylcarnitines and risk of type 2 diabetes in a Mediterranean population at high cardiovascular risk. J. Clin. Endocrinol. Metab. 2018. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.; Knight, R.; Gordon, J.I. The human microbiome project: Exploring the microbial part of ourselves in a changing world. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Thomas, S.; Izard, J.; Walsh, E.; Batich, K.; Chongsathidkiet, P.; Clarke, G.; Sela, D.A.; Muller, A.J.; Mullin, J.M.; Albert, K.; et al. The Host Microbiome Regulates and Maintains Human Health: A Primer and Perspective for Non-Microbiologists. Cancer Res. 2017, 77, 1783–1812. [Google Scholar] [CrossRef]
- Lopez-Legarrea, P.; Fuller, N.R.; Zulet, M.A.; Martinez, J.A.; Caterson, I.D. The influence of Mediterranean, carbohydrate and high protein diets on gut microbiota composition in the treatment of obesity and associated inflammatory state. Asia Pac. J. Clin. Nutr. 2014, 23, 360–368. [Google Scholar]
- Marlow, G.; Ellett, S.; Ferguson, I.R.; Zhu, S.; Karunasinghe, N.; Jesuthasan, A.C.; Han, D.Y.; Fraser, A.G.; Ferguson, L.R. Transcriptomics to study the effect of a Mediterranean-inspired diet on inflammation in Crohn’s disease patients. Hum. Genom. 2013, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Del Chierico, F.; Gnani, D.; Vernocchi, P.; Petrucca, A.; Alisi, A.; Dallapiccola, B.; Nobili, V.; Lorenza, P. Meta-Omic Platforms to Assist in the Understanding of NAFLD Gut Microbiota Alterations: Tools and Applications. Int. J. Mol. Sci. 2014, 15, 684–711. [Google Scholar] [CrossRef] [PubMed]
- Tosti, V.; Bertozzi, B.; Fontana, L. Health Benefits of the Mediterranean Diet: Metabolic and Molecular Mechanisms. J. Gerontol. Ser. A 2017, 73, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Bajzer, M.; Seeley, R.J. Physiology: Obesity and gut flora. Nature 2006, 444, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Caesar, R.; Fåk, F.; Bäckhed, F. Effects of gut microbiota on obesity and atherosclerosis via modulation of inflammation and lipid metabolism. J. Intern. Med. 2010, 268, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Diamant, M.; Blaak, E.E.; de Vos, W.M. Do nutrient-gut-microbiota interactions play a role in human obesity, insulin resistance and type 2 diabetes? Obes. Rev. 2011, 12, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.; Fairlie, D.P.; Prins, J.B.; Hammock, B.D.; Brown, L. Inflammatory lipid mediators in adipocyte function and obesity. Nat. Rev. Endocrinol. 2010, 6, 71–82. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, M.; Wang, S.; Han, R.; Cao, Y.; Hua, W.; Mao, Y.; Zhang, X.; Pang, X.; Wei, C.; et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010, 4, 232–241. [Google Scholar] [CrossRef]
- Gutierrez-Diaz, I.; Fernandez-Navarro, T.; Sanchez, B.; Margolles, A.; Gonzalez, S. Mediterranean diet and faecal microbiota: A transversal study. Food Funct. 2016, 7, 2347–2356. [Google Scholar] [CrossRef]
- Fitó, M.; Guxens, M.; Corella, D.; Sáez, G.; Estruch, R.; de la Torre, R.; Francés, F.; Cabezas, C.; López-Sabater, M.D.C.; Marrugat, J.; et al. Effect of a traditional Mediterranean diet on lipoprotein oxidation: A randomized controlled trial. Arch. Intern. Med. 2007, 167, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Cardona Diaz, F.; Andrés-Lacueva, C.; Tinahones, F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, S.M.; Meydani, M.; Barnett, J.B.; Goldin, B.; Kane, A.; Rasmussen, H.; Brown, C.; Vangay, P.; Knights, D.; Jonnalagadda, S.; et al. Substituting whole grains for refined grains in a 6-wk randomized trial has a modest effect on gut microbiota and immune and inflammatory markers of healthy adults. Am. J. Clin. Nutr. 2017, 105, 635–650. [Google Scholar] [CrossRef] [PubMed]
- Shankar, V.; Gouda, M.; Moncivaiz, J.; Gordon, A.; Reo, N.V.; Hussein, L.; Paliy, O. Differences in Gut Metabolites and Microbial Composition and Functions between Egyptian and U.S. Children Are Consistent with Their Diets. mSystems 2017, 2, e00169-16. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- Bohn, T.; Desmarchelier, C.; Dragsted, L.O.; Nielsen, C.S.; Stahl, W.; Rühl, R.; Keijer, J.; Borel, P. Host-related factors explaining interindividual variability of carotenoid bioavailability and tissue concentrations in humans. Mol. Nutr. Food Res. 2017, 61, 1600685. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Ren, J.; Ruffin, M.T.; Turgeon, D.K.; Brenner, D.E.; Sidahmed, E.; Rapai, M.E.; Cornellier, M.L.; Djuric, Z. Relationships between Serum and Colon Concentrations of Carotenoids and Fatty Acids in Randomized Dietary Intervention Trial. Cancer Prev. Res. 2013, 6, 558. [Google Scholar] [CrossRef]
- Djuric, Z.; Bassis, C.M.; Plegue, M.A.; Ren, J.; Chan, R.; Sidahmed, E.; Turgeon, D.K.; Ruffin, M.T., IV; Kato, I.; Sen, A. Colonic Mucosal Bacteria Are Associated with Inter-Individual Variability in Serum Carotenoid Concentrations. J. Acad. Nutr. Diet. 2018, 118, 606–616. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812. [Google Scholar] [CrossRef]
- Gutiérrez-Díaz, I.; Fernández-Navarro, T.; Salazar, N.; Bartolomé, B.; Moreno-Arribas, M.V.; de Andres-Galiana, E.J.; Fernández-Martínez, J.L.; de los Reyes-Gavilán, C.G.; Gueimonde, M.; González, S. Adherence to a Mediterranean Diet Influences the Fecal Metabolic Profile of Microbial-Derived Phenolics in a Spanish Cohort of Middle-Age and Older People. J. Agric. Food Chem. 2017, 65, 586–595. [Google Scholar] [CrossRef]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Front. Microbiol. 2018, 9, 890. [Google Scholar] [CrossRef]
- Haro, C.; Montes-Borrego, M.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Tinahones, F.J.; Landa, B.B.; et al. Two Healthy Diets Modulate Gut Microbial Community Improving Insulin Sensitivity in a Human Obese Population. J. Clin. Endocrinol. Metab. 2016, 101, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55. [Google Scholar] [CrossRef] [PubMed]
- Haro, C.; García-Carpintero, S.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Landa, B.B.; Clemente, J.C.; Pérez-Martínez, P.; López-Miranda, J.; Pérez-Jiménez, F.; Camargo, A. Consumption of Two Healthy Dietary Patterns Restored Microbiota Dysbiosis in Obese Patients with Metabolic Dysfunctio. Mol. Nutr. Food Res. 2017, 61, 1700300. [Google Scholar] [CrossRef] [PubMed]
- Mitsou, E.K.; Kakali, A.; Antonopoulou, S.; Mountzouris, K.C.; Yannakoulia, M.; Panagiotakos, D.B.; Kyriacou, A. Adherence to the Mediterranean diet is associated with the gut microbiota pattern and gastrointestinal characteristics in an adult population. Br. J. Nutr. 2017, 117, 1645–1655. [Google Scholar] [CrossRef]
- Haro, C.; Garcia-Carpintero, S.; Alcala-Diaz, J.F.; Gomez-Delgado, F.; Delgado-Lista, J.; Perez-Martinez, P.; Rangel Zuñiga, O.A.; Quintana-Navarro, G.M.; Landa, B.B.; Clemente, J.C.; et al. The gut microbial community in metabolic syndrome patients is modified by diet. J. Nutr. Biochem. 2016, 27, 27–31. [Google Scholar] [CrossRef]
- Cani, P.D. Human gut microbiome: Hopes, threats and promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef]
- UNC. Gut Microbiome, Diet and Health. Available online: https://www.uncnri.org/index.php/gut-microbiome-diet-and-health/ (accessed on 20 January 2019).
- Zmora, N.; Zeevi, D.; Korem, T.; Segal, E.; Elinav, E. Taking it Personally: Personalized Utilization of the Human Microbiome in Health and Disease. Cell Host Microbe 2016, 19, 12–20. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef]
- Ottosson, F.; Brunkwall, L.; Ericson, U.; Nilsson, P.M.; Almgren, P.; Fernandez, C.; Melander, O.; Orho-Melander, M. Connection Between BMI-Related Plasma Metabolite Profile and Gut Microbiota. J. Clin. Endocrinol. Metab. 2018, 103, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Hadrich, D. Microbiome Research Is Becoming the Key to Better Understanding Health and Nutrition. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [PubMed]



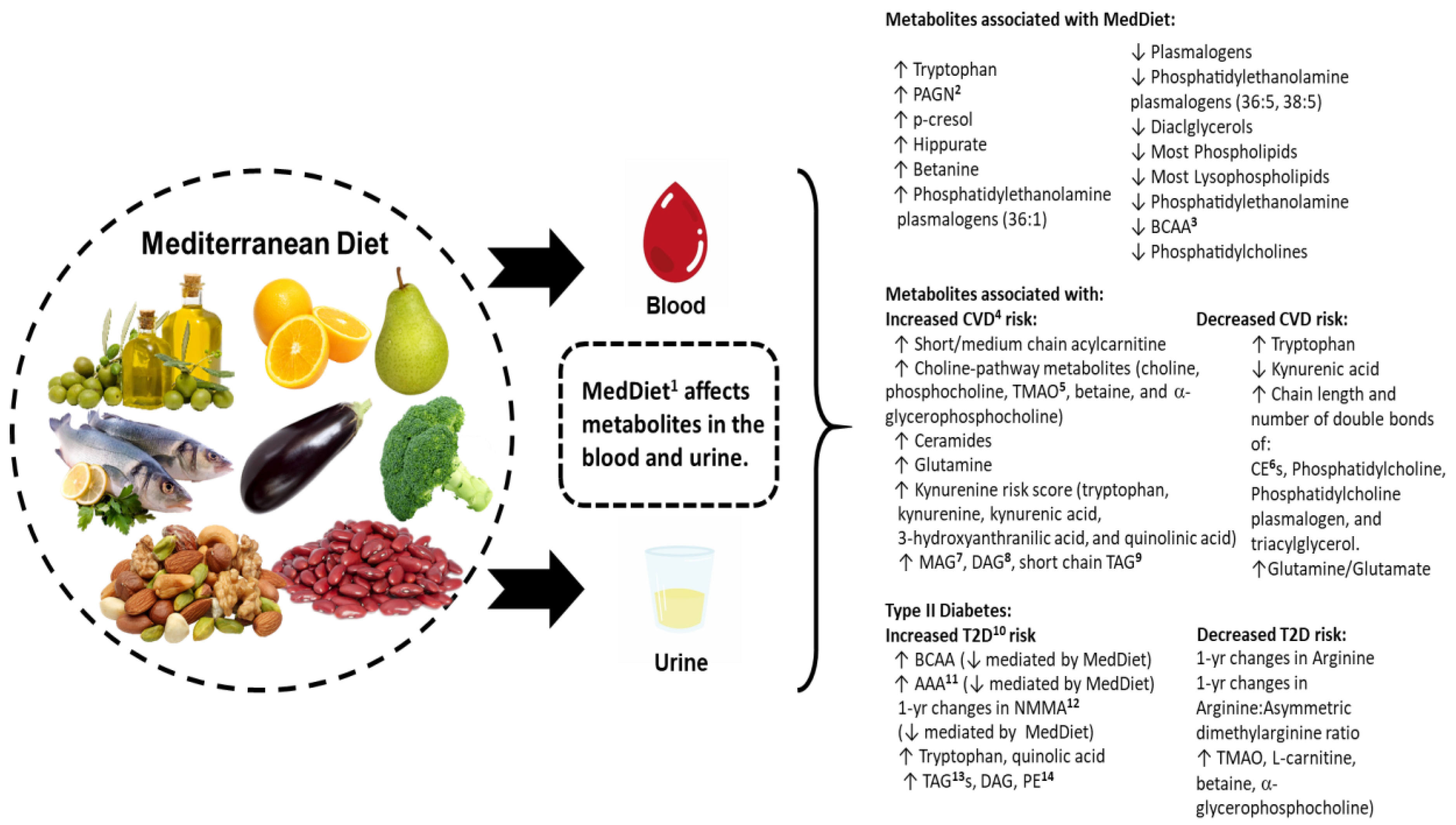{kind=link}
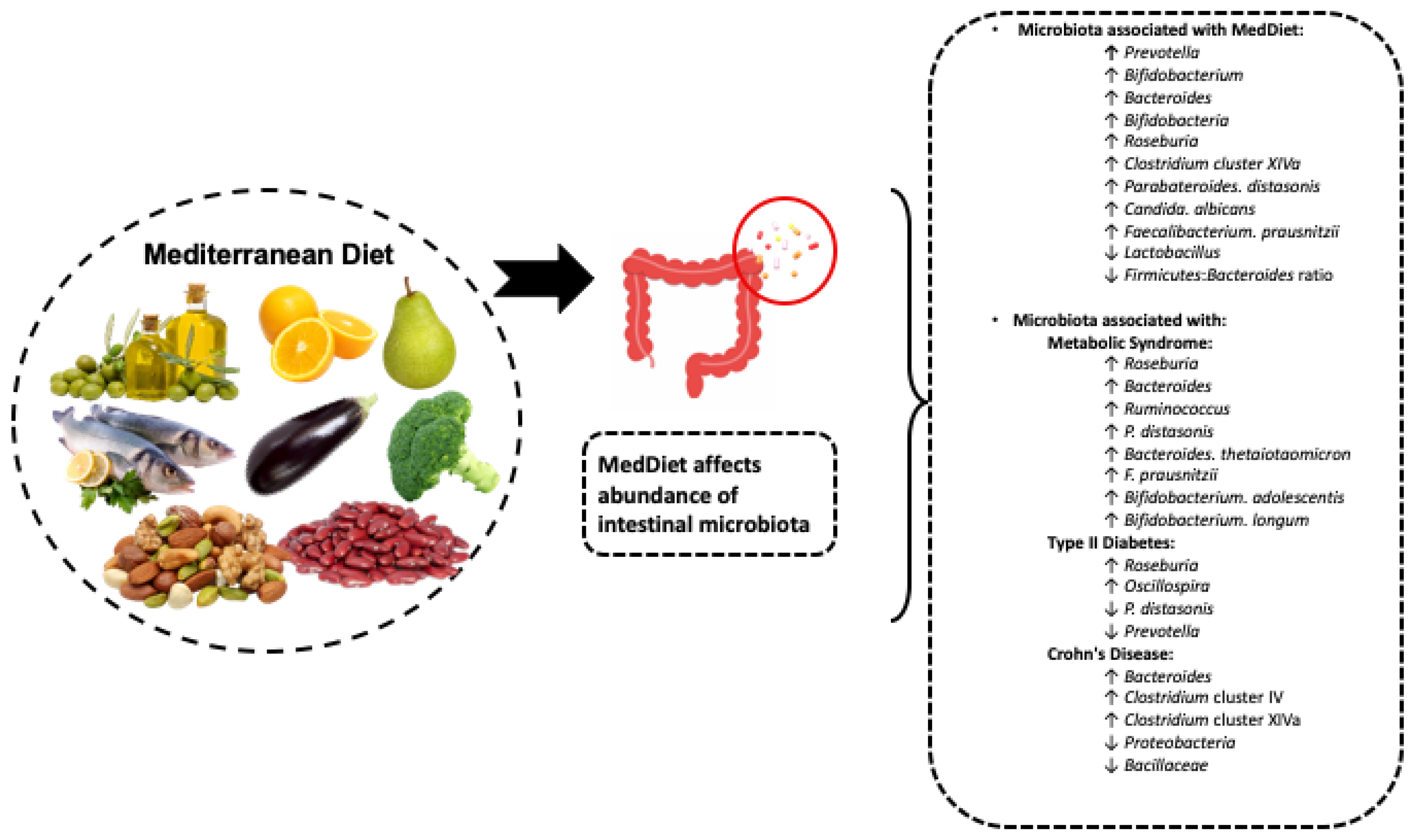{kind=link}
| Study Design | Participants | Dietary Pattern/Intervention | Follow-Up | Biological Sample/Metabolomics Approach/Technique | Biomarkers Identified | Main Conclusion | |
|---|---|---|---|---|---|---|---|
| Playdon et al., 2017 [51] | 5 nested case-control studies (within the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study) | Male Finnish smokers n = 1336, aged 50–69 years | HEI 1 2010, aMED 2, HDI 3, and BSD 4 | 3 years | Serum Untargeted MS | HEI 2010, HDI and BSD: associated with 17, 11, and 10 identified metabolites, respectively. aMED: associated with 21 identifiable metabolites: 4 aminoacids (indolebutyrate, tryptophan betaine, N-methylproline, 3-Hydroxy-2-ethylpropionate); 1 carbohydrate (threitol); 2 co-factors (threonate, γ-CEHC 5); 3 xenobiotics (stachydrine, Phytanate, ergothionein) 11 lipids (1-myristoleoylglycerophosphocholine (14:1), Scyllo-inositol, Mead acid (20:3n -9), g-CEHC, cis-4-Decenoyl carnitine, 3-Carboxy-4-methyl-5-propyl-2-furanpropanoate, linoleate (18:2n-6), linolenate (α or γ; 18:3n-3 or 18:3n-6), chiro-inositol, 1-linoleoylglycerol, DHA, methyl palmitate | The HEI-2010, aMED, HDI, and BSD were associated with metabolites correlated with foods that are used to evaluate adherence to each score. |
| Vázquez-Fresno et al., 2015 [53] | Parallel-group, single-blind, multicenter, randomized, controlled feeding trial. A follow-up in the PREDIMED 6 study. | Clinically identified non-diabetic participants at high CVD 7 risk n = 98 aged 55–80 years | MedDiet 8 + EVOO 9 (n = 41) (MedDiet + nuts (n = 27) LFD 10 (n = 30) | 3 years | Urine (baseline, 1 year, and 3 year of the intervention) Untargeted NMR | MedDiet: carbohydrates (3-HB 11, citrate, and cisaconitate), creatine, creatinine, amino acids (proline, N-acetylglutamine, glycine, branched-chain amino acids, and derived metabolites), lipids (oleic and suberic acids), and microbial cometabolites (PAGN 12 and p-cresol) LFD: hippurate, TMAO 13, anserine, histidine and derivates (3-MH 14, 1-MH, carnosine, anserine), and xanthosine. | The MedDiet groups had distinct metabolic profiles compared to the baseline and control group related to carbohydrate and lipid metabolism, amino acids, and microbial cometabolites (PAGN and p-cresol) |
| Bondia-Pons et al., 2015 [52] | Randomized controlled dietary intervention | Individuals with high BMI and at least two features of metabolic syndrome. N = 72 | RESMENA 15 diet (n = 47) (based on MedDiet). 7 meals/day. (40% CHO 16, 30%protein, 30% lipid). Control diet (n = 45) (American Heart Association guidelines). 5 meals/day (55% CHO 55%, 15% protein, 30% lipid) | 6-month (2-month nutritional learning followed by a 4-month self-control period) | Plasma Non-targeted MS-Liquid | Lipids, mainly phospholipids and lysophospholipids. lactic acid, L-isoleucine, alloisoleucine, hydroxyvaleric acid, hypaphorine, paraxanthine, hippuric acid, furancarboxylic acid, LysoPC 17 (14:0), LysoPC (20:5), LysoPC (16:1), LysoPC (22:6), LysoPE (20:4), LysoPE 18 (18:2), LysoPC (16:0), Linoleamide, LysoPC (20:3), LysoPE (18:1), LysoPC (18:1), LysoPC (20:4), Eicosapentaenoic acid, LysoPC (15:0), Lithocholic acid, Oleamide, 1-Monopalmitin, LysoPC (18:0), GPL 19 containing (18:2), Palmitic acid, PC 20, and PE 21 | The major discriminative markers between the two groups were the plasmalogen PC (P 22-18:1/20:3) after 2 months and palmitic acid after 6 months. |
| González-Guardia et al., 2015 [55] | Randomized, crossover | Men (n = 5) and women (n = 5) aged 65 years or older. n = 10 | MedDiet+ 200mg/d CoQ 23; MedDiet without CoQ Western diet rich in SFA 24; Low-fat, high-carbohydrate diet enriched in n-3 PUFA25. | 4 weeks | Urine and plasma Targeted NMR | CoQ and β-carotene plasma levels and isoprostanes urinary levels were determined. Higher levels of hippurate and lower levels of phenylacetylglycine were found when comparing the MedDiet + CoQ and the SFA. | The MedDiet supplemented with CoQ is associated with increased levels of excreted hippurate and decreased levels of phenylacetylglycine compared with a SFA-rich diet. |
| Kakkoura et al., 2017. [56] | Cross-sectional study. | Greek-Cypriot control women who have previously participated in the population-based case- control study of BC, MASTOS [61]. n = 564 | MedDiet (highest and lowest adherence to MedDiet) | N/A | Serum Targeted. UPLC-MS/MS | 5-MTHF 26, riboflavin, FMN 27, PA, methionine, methionine sulfoxide, SAM 28, SAH 29, total HCY 30, cystathionine, total cysteine, γ-glu-cys, total GSH 31, and α-hydroxybutyrate. | Higher adherence to the MedDiet was associated with an increase in antioxidant-related metabolites, 5-MTHF. |
| Author | Study Design | Study Population | Dietary Pattern/Intervention | Follow-Up | Biological Sample/Metabolomics Approach | Metabolites Examined | Main Conclusion |
|---|---|---|---|---|---|---|---|
| Guasch-Ferré et al., 2016 [66] | Case–cohort study within the PREDIMED 1 study | Participants aged 55–80 years at high risk of CVD 2. n = 980 (229 cases CVD cases). | MedDiet 3 groups: (MedDiet + EEVO 4 and MedDiet + mixed nuts. Control group: low-fat diet. | 4.8 years | Plasma. (baseline and after 1 year) Targeted | 28 acylcarnitines: short-chain acylcarnitines (C2–C7), medium-chain acylcarnitines (C8–C14), and long-chain acylcarnitines (C16–C26). | An increased level of acylcarnitines metabolic profiles is independently associated with total CVD risk and risk of stroke. MedDiet interventions may attenuate the association between acylcarnitines and CVD risk. |
| Guasch-Ferré et al., 2017 [67] | Case–cohort study within the PREDIMED study | Participants aged 55–80 years at high risk of CVD. n = 980 (229 CVD cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet. | 4.8 years | Plasma (baseline and after 1 year) Targeted | Metabolites of the choline pathway: TMAO 5, betaine, choline, phosphocholine, and a-glycerophosphocholine. A choline metabolite score was created. | The baseline choline metabolite score was associated with increased risk of CVD. The one-year changes in plasma metabolites were not significantly associated with CVD. The participants in the highest metabolite score quartile and assigned to low-fat diets had higher risk of CVD than those in the lowest metabolite quartile and in the MedDiet group. No significant interaction was found between the continuous choline score, the betaine/choline ratio, and the intervention group and CVD |
| Yu et al., 2017 [68] | Case–cohort study within the PREDIMED study, controlled trial | Participants aged 55–80 at high risk of CVD. n = 985 (231 CVD cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet. | 4.7 year | Plasma (baseline and after 1 year) Targeted | Tryptophan, kynurenine, kynurenic acid, 3-hydroxyanthranilic acid, and quinolinic acid concentrations. A KRS 6 was created. | The positive association between the KRS and CVD risk is stronger in the control group, indicating that the MedDiet may attenuate the effect of a high KRS score. |
| Toledo et al., 2017 [77] | Case–cohort study within the PREDIMED study | Participants aged 55–80 years at high risk of CVD. n = 983 (230 CVD cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet. | 4.8 years | Plasma (baseline and after 1 year) Untargeted lipidome | 202 lipid species | The baseline concentrations of cholesterol esters (CEs) were inversely associated with CVD. The MedDiet interventions resulted in changes in the lipidome at 1 year; however, they were not found to be associated with subsequent CVD risk. Lipid metabolites with a longer acyl chain and a higher number of double bonds at the baseline were significantly and inversely associated with the risk of CVD. |
| Wang et al., 2017 [69] | Case–cohort study within the PREDIMED study | Participants aged 55–80 years at high risk of CVD. n = 980 (230 CVD cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet. | ≤7.4 years | Plasma (baseline and after 1 year) Targeted | 4 different ceramides: ceramide (d 7 18:1/16:0), ceramide (d18:1/22:0), ceramide (d18:1/24:0), and ceramide (d18:1/24:1). A ceramide score was calculated. | The ceramide score was positively associated with the risk of CVD. The MedDiet may alleviate the potential negative effects of increased plasma ceramide levels on CVD. |
| Zheng et al., 2017 [70] | Case–cohort study within the PREDIMED study | Participants aged 55–80 years at high risk of CVD. n = 980 (788 subcohort, 192 incident external cases) | MedDiet groups (intervention diets): (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet. | 4.8 years | Plasma (baseline and after 1 year) Targeted | Glu14 8 Gln15 9, Glu/Gln ratio No significant effect of the intervention on one-year changes in the metabolites. No effect of the changes themselves on the CVD risk was apparent. | A positive association between Glu levels and CVD risk (43% increased risk) and a negative association between Gln/Glu and risk of CVD (25% decreased risk) were found. The interventions effectively lowered CVD risk for the participants with high baseline Glu, while no effects were found among the participants with low baseline Glu. |
| Razquin et al., 2018 [71] | Unstratified case–cohort design within the PREDIMED study. | n = 983 participants (233 CVD cases). | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet. | 4.8 years | Plasma (baseline and after 1 year) | Lipid group A: PC 10 (PCs, LysoPC11s and PC-plasmalogens with ≥5 double bonds); CE 12 with N3 double bonds; and TAG 13 with ≥52 carbon atoms containing ≥6 double bonds. Lipid group B: MAG 14; DAG 15; short-chain, TAGs containing ≤4 double bonds; PEs 16 except those with saturated fatty acids; hydroxyPC. PC, CE, long-chain TAG, MAG and DAG, short-chain TAG, PE, and Hpc 17 scores were calculated. | The metabolites from lipid group A were inversely associated with CVD; the metabolites from lipid group B were directly associated with CVD. The baseline phosphatidylethanolamines (PEs) and their one-year changes tended to be associated with higher CVD risk. No significant effect of the MedDiet intervention was found on the metabolite scores. |
| Ruiz-Canela et al., 2016 [78] | Case–cohort study within the PREDIMED study | n = 970 (226 CVD cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet. | 4.8 years | Plasma (baseline and after 1 year) | BCAAs | Higher concentrations of baseline BCAAs were associated with increased risk of CVD. No significant effect of the intervention on one-year changes in BCAAs or any association between one-year changes in BCAAs and CVD were observed. |
| Yu et al., 2017 [73] | Case–cohort study within the PREDIMED study | n = 984 (231 CVD cases) | MedDiet groups: (MedDiet +EEVO and MedDiet + mixed nuts. Control group: low-fat diet. | 4.7 years | Plasma (baseline and after 1 year) | arginine, ornithine, citrulline, ADMA 18, symmetric dimethylarginine (SDMA 19), and NG-monomethylarginine (NMMA 20) | A higher baseline arginine/asymmetric dimethylarginine ratio was associated with lower CVD incidence. No significant modification by the MedDiet after one-year intervention was observed. |
| Guasch-Ferre et al., 2018 [79] | Case–cohort study in the PREDIMED study | n = 892 participants (251 T2D cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet | 3.8 years | Plasma (baseline and after 1 year) | Short-chain acylcarnitines (C2–C7), medium-chain acylcarnitines (C8–C14), and long-chain acylcarnitines (C16–C26). | The acylcarnitines profile, specifically short- and long- chain acylcarnitines, was significantly associated with a higher risk of T2D. |
| Yu et al., 2018 [73] | Case–cohort study in the PREDIMED study | n = 892 participants (251 T2D cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet | 1 year | Plasma | Arginine, citrulline, ornithine, ADMA, SDMA, and NMMA | The one-year changes in arginine and the arginine/ADMA ratio were negatively associated with the risk of T2D 21. Positive changes in ornithine and citrulline and negative changes in SDMA and GABR were inversely associated with concurrent changes in HOMA-IR 22 The MedDiet significantly modified the association between one-year changes in NMMA and T2D risk. |
| Ruiz-Canela et al., 2018 [72] | Case–cohort study in the PREDIMED study | n = 945 participants (251 T2D cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet | 3.8 years | Plasma (baseline and after 1 year) | The baseline BCAA 23 (leucine, isoleucine and valine) and AAA 24 (phenylalanine and tyrosine) scores were associated with a higher risk of T2D. Increases in the BCAA score after one year were associated with higher T2D risk only in the control group. | The MedDiet rich in EVOO significantly reduced the levels of BCAA and attenuated the positive association between plasma BCAA levels and T2D incidence. |
| Papandreou et al. 2018 [76] | Case–cohort study in the PREDIMED study | n = 945 participants (251 T2D cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet | 3.8 years | Plasma (baseline and after 1 year) | TMAO, L-carnitine, betaine, LPC and LPE species, phosphocholine, α-glycerophosphocholine, and choline. Higher baseline concentrations of TMAO, L-carnitine, betaine, α-glycerophosphocholine, and several LPC 25 and LPE 26 species were associated with a lower risk of T2D development. | There was no significant difference in the association of most of the one-year changes in the metabolites with T2D risk in the MedDiet intervention and control groups. The intervention diets did not appear to significantly change the study metabolite levels during the intervention. |
| Razquin et al. (2018) [74] | Case–cohort study in the PREDIMED study | n = 942 participants (250 T2D cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet | 3.8 years | Plasma (baseline and after 1 year) | The baseline TAGs, DAGs, and PEs were positively associated with T2D risk. TAGs with odd-chain fatty acids showed inverse associations with T2D after adjusting for total TAGs. | The one-year changes in the baseline metabolites associated with T2D were not significant. The changes in LP 27, PC-PL 28, SM 29, and CE scores showed no apparent mediating effects. |
| Yu et al., 2018 [75] | Case–cohort study in the PREDIMED study | n = 892 participants (251 T2D cases) | MedDiet groups: (MedDiet + EEVO and MedDiet + mixed nuts. Control group: low-fat diet | 3.8 years | Plasma (baseline and after 1 year) Targeted | Tryptophan, kynurenine, kynurenic acid, 3-hydroxyanthranilic acid, and quinolinic acid concentrations. A KRS score was created. | The baseline tryptophan and one-year increases in quinolinic acid were positively associated with incident T2D. No effect of the MedDiet was observed. |
| Author | Study Design | Study Population | Dietary Pattern/Intervention | Follow-up | Sample | Microbiota Observed | Results/Conclusion |
|---|---|---|---|---|---|---|---|
| Gutierrez-Diaz et al., 2016 [92] | Cross-sectional | Adults with a non-declared pathology; n = 31 (23 females, 8 males, mean age of 42.1 years | MedDiet 1 score (0–8 points; > 4 = High adherence) | N/A | Stool | Bifidobacterium, Faecalibacterium, Tenericutes, Dorea, Rikenellaceae, Alistipes, Ruminococcus (Lechnospiraceae family), Coprococcu, Bacteroidetes, Prevotel-lacceae, Prevotella, and Firmicutes | The MedDiet score was associated with a higher abundance of Bacteroidetes, Prevotel-laceae, and Prevotella and a lower concentration of Firmicutes and Lachnospiraceae. |
| Gutierrez-Diaz et al., 2017 [102] | Cross-sectional | Healthy men (n = 20) and women (n = 54) older than 50 years of age | MedDiet | N/A | Stool | Akkermansia, Bacteroides-Prevotella-Porphiromonas, Bifidobacterium, Clostridium cluster XIVa, Lactobacillus group, and F. prausnitzii | Higher levels of Clostridium cluster XIVA and F. prausnitzii were found in subjects with MDS 2 scores ≥4 and were positively correlated with fecal concentrations of benzoic and 3-hydroxyphenylacetic acids and the intake of polyphenols and fibers. |
| De Filippis et al., 2016 [101] | Cross-sectional study | Healthy Volunteers n = 153 | MedDiet Vegan Vegetarian Omnivore | N/A | Stool, urine | Lachnospira, Prevotella, Roseburia, and Ruminococcus | Plant-based diets appear to increase fecal SCFAs, while Prevotella specifically was associated with fiber-degrading Firmicutes. Higher urinary trimethylamine oxide levels were found to be higher in those with lower MedDiet adherence. Beneficial microbiome-related metabolic profiles were associated with the increased consumption of plant-based foods, consistent with a MedDiet. |
| Garcia-Mantrana et al., 2018 [103] | Cross-sectional study | Healthy individuals n = 27, mean age 39.5 years | MedDiet | N/A | Stool samples | Enterobacteriaceae family, Bifidobacterium group, Bacteroides-Prevotella-Porphyromonas group, Bacteroides fragilis group, Blautia coccoides group, Methanobrevibacter smithii, and Faecalibacterium prausnitzii | A higher ratio of Firmicutes–Bacteroidetes was related to lower adherence to the MedDiet, and greater presence of Bacteroidetes was associated with lower animal protein intake. Better adherence to the MedDiet was associated with significantly higher levels of total SCFA 3. |
| Shankar et al., 2017 [96] | Comparative cross-sectional stud | Healthy Egyptian male teenagers n = 28, mean age 13.9 years Healthy American male teenagers n = 14, mean age 12.9 years | MedDiet Western diet | N/A | Stool | Egyptian: Gammaproteobacteria, Methanobacteria, Prevotella, Megasphaera, Eubacterium, Mitsuokella, Catenibacterium U.S.: Clostridia, Verrucomicrobia, Bacteroides, Ruminococcus, Coprococcus, Blautia, Bilophila, Akkermansia, and Faecalibacterium, | Egyptian gut microbial communities belonged to Prevotella in all the subjects with increased polysaccharide-degrading microbes and end products of polysaccharide fermentation. United States (US) gut microbial communities mostly belonged to Bacteroides with increased proteolytic microbes and end products of protein and fat metabolism. |
| Djuric et al., 2018 [100] | Randomized control trial | n = 88 baseline samples n = 82 post-intervention (men and women) Mean age 53 years | MedDiet (30% kcals form fat, PUFA/SAT/MUFA 4 ratios of 1:2:5, foods high in n-3 fatty acids 2x/week, 3 servings/day whole grains, 7–9.5 cup s/day F 5+V 6) including at least one cup dark green or orange F or V). Healthy Eating diet (5.5 cup servings/day F + V, 3 s/day whole grains, <10% kcals from sat. fat. | 6 months | Blood, colon biopsy | Firmicutes, Proteobacteria, Lachnospiraceae, Blautia, Roseburia, Prevotella, and Bacteroides, | A total of 11 operational taxonomic units were significantly associated with increased serum carotenoid levels. The Bacteria in the colonic mucosa was resistant to change after both diet interventions The intestinal microbiota did not show significant changes after 6 months of diet intervention; however, an abundance of specific OTUs 7 was significantly associated with serum carotenoid concentrations at the baseline, suggesting that long-term dietary exposures may have more of an influence on bacteria in the colonic mucosa. |
| Design | Participant Characteristics | Dietary Pattern/Treatment | Length | Sample | Microbiota Observed | Results/Conclusion | |
|---|---|---|---|---|---|---|---|
| Haro et al., 2016 [109] | Randomized control trial | n = 138 with metabolic syndrome (MetS) and n = 101 without MetS; male and female patients within the CORDIOPREV study with CHD 1, who had their last coronary event over 6 months before enrolling, in addition to conventional treatment for CHD, | MedDiet group:35% fat (22% MUFA, 6% PUFA, 7% SAT). Low-fat high-complex carb (LFHCC) diet group: 28% fat (12% MUFA, 8% PUFA, 8% SAT). | 2 years | Stool, blood | Bacteroides, Eubacterium, Lactobacillus, Bacteroides fragilis group, Parabacteroides distasonis, Bacteroides thetaiotaomicron, Faecalibacterium prausnitzii, Fusobacter- ium nucleatum, Bifidobacterium longum, Bifidobacterium adolescentis, Ruminococcus flavefaciens subgroup, and Eubacterium rectale | The long-term consumption of the Mediterranean diet partially restores the population of P. distasonis, B. thetaiotaomicron, F. prausnitzii, B. adolescentis and B. longum in MetS patients although MetS persists. |
| Haro et al., 2016 [104] | Randomized control trial | n= 20, 40 total samples collected (20 at the baseline, 20 post-intervention) from obese men with CHD within the CORDIOPREV study | MedDiet group: 35% fat (22% MUFA, 6% PUFA, 7% SAT). Low-fat high-complex carb (LFHCC) diet group: 28% fat (12% MUFA, 8% PUFA, 8% SAT). | 1 year | Blood, stool | Bacteroides, Prevotella, unknown Lachnospiraceae, Faecalibacterium, unknown Clostridiales, unknown Ruminococcaceae, Oscillospira, Parabacteroides, and unknown Bacteroidales | Both diet changes increased insulin sensitivity and appeared to exert protective effects on the development of T2DM 2 based off of specific changes in gut microbiota. Changes in feces include mostly amino acids, peptides, and shingolipid metabolism, which may be linked to changes occurring in the gut microbiota. |
| Haro et al., 2017 [107] | Randomized control trial | n = 33 obese patients with severe MetS–OB 3, n = 32 obese patients without non-MetS–OB, and 41 non-obese subjects (non-MetS–non-OB). | MedDiet group: 35% fat (22% MUFA, 6% PUFA, <10% SAT). Low-fat diet group: <30% total fat (<10% SAT., 12–14% MUFA, 6–8% PUFA). | 2 years | Stool | Actinobacteria, Bacteroidetes, Firmicutes, Bacteroides, Prevotella, Roseburia, Faecalibacterium, Ruminococcus, Streptococcus, Clostridium, P. distasonis, and F. prausnitzii | Both diets were associated with partially restored gut microbiome dysbiosis, converting MetS-OB microbiota patterns to microbiota patterns similar to those found in (metabolically) healthy people, after 2 years of nutrition intervention in participants with coronary heart disease. The degree of participants’ metabolic dysfunction may alter the effectiveness of nutrition therapy. |
| Marlow et al., 2014 [83] | Non-randomized trial | n = 8 Crohn’s patients with no history of bowel surgery who were not taking prednisone or similar anti-inflammatory medication and had no changes in medication over the last 3 months. | MedDiet | 6 weeks | Stool, blood | Firmicutes, Bacteroidetes, Actinobacteria, Proteo- bacteria, Fusobacteria, and Verrucomicrobia | The Mediterranean-inspired diet appeared to benefit the health of people with Crohn’s disease. The participants showed a trend for reduced markers of inflammation and normalization of the microbiota; however, the changes were not significant. |
| Mitsu et al., 2017 [108] | Cross-sectional | n = 120 Men and women, age 18–65 years | MedDiet Tertiles of adherence based on MedDiet score: Low tertile score = 19–30 (n = 31) Medium tertile score = 31–33 (n = 29) High tertile score = 34–41 (n = 40) | N/A | Stool | E. coli, bifidobacteria, and Candida albicans | The findings support a link between MedDiet adherence and the gut microbiota profile. Those with high adherence had lower E. coli counts, a higher bifidobacterial/E. coli ratio, and increased levels and prevalence of Candida albicans when compared to those with low adherence |
| Garcia-Mantrana et al., 2018 [103] | Cross-sectional study | Healthy individuals n = 27, mean age 39.5 years | MedDiet | Stool samples | Enterobacteriaceae family, Bifidobacterium group, Bacteroides-Prevotella-Porphyromonas group, Bacteroides fragilis group, Blautia coccoides group, Methanobrevibacter smithii, and Faecalibacterium prausnitzii | Butyricimonas, Desulfovibrio, and Oscillospira genera were associated with a BMI of <25 and the genus Catenibacterium was associated with a higher adherence to the MedDiet. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Q.; Black, A.; Kales, S.N.; Vattem, D.; Ruiz-Canela, M.; Sotos-Prieto, M. Metabolomics and Microbiomes as Potential Tools to Evaluate the Effects of the Mediterranean Diet. Nutrients 2019, 11, 207. https://doi.org/10.3390/nu11010207
Jin Q, Black A, Kales SN, Vattem D, Ruiz-Canela M, Sotos-Prieto M. Metabolomics and Microbiomes as Potential Tools to Evaluate the Effects of the Mediterranean Diet. Nutrients. 2019; 11(1):207. https://doi.org/10.3390/nu11010207
Chicago/Turabian StyleJin, Qi, Alicen Black, Stefanos N. Kales, Dhiraj Vattem, Miguel Ruiz-Canela, and Mercedes Sotos-Prieto. 2019. "Metabolomics and Microbiomes as Potential Tools to Evaluate the Effects of the Mediterranean Diet" Nutrients 11, no. 1: 207. https://doi.org/10.3390/nu11010207
APA StyleJin, Q., Black, A., Kales, S. N., Vattem, D., Ruiz-Canela, M., & Sotos-Prieto, M. (2019). Metabolomics and Microbiomes as Potential Tools to Evaluate the Effects of the Mediterranean Diet. Nutrients, 11(1), 207. https://doi.org/10.3390/nu11010207