Neuroprotective Effects of Taraxacum officinale Wigg. Extract on Glutamate-Induced Oxidative Stress in HT22 Cells via HO-1/Nrf2 Pathways


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Extract
2.3. Characterization of ETOW
2.4. Cell Culture
2.5. Cell Viability Assays
2.6. Measurement of Reactive Oxygen Species Assay
2.7. Preparation of Cytosolic and Nuclear Fractions
2.8. Western Blot
2.9. RNA Extraction and Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
2.10. Immunofluorescence
2.11. Statistical Analysis
3. Results
3.1. Characterization of ETOW by LC/MS
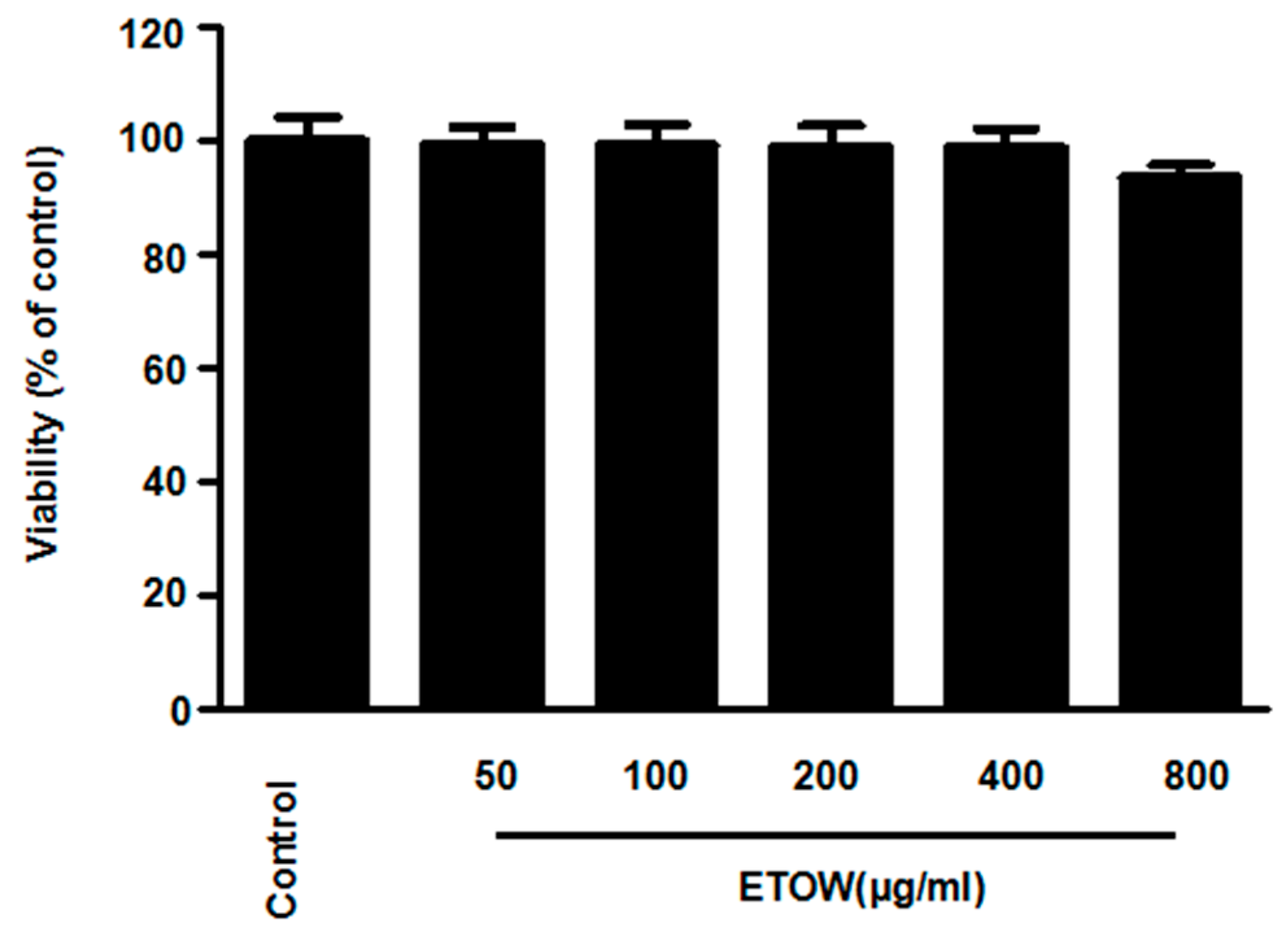
3.2. Effects of ETOW on Cell Viability
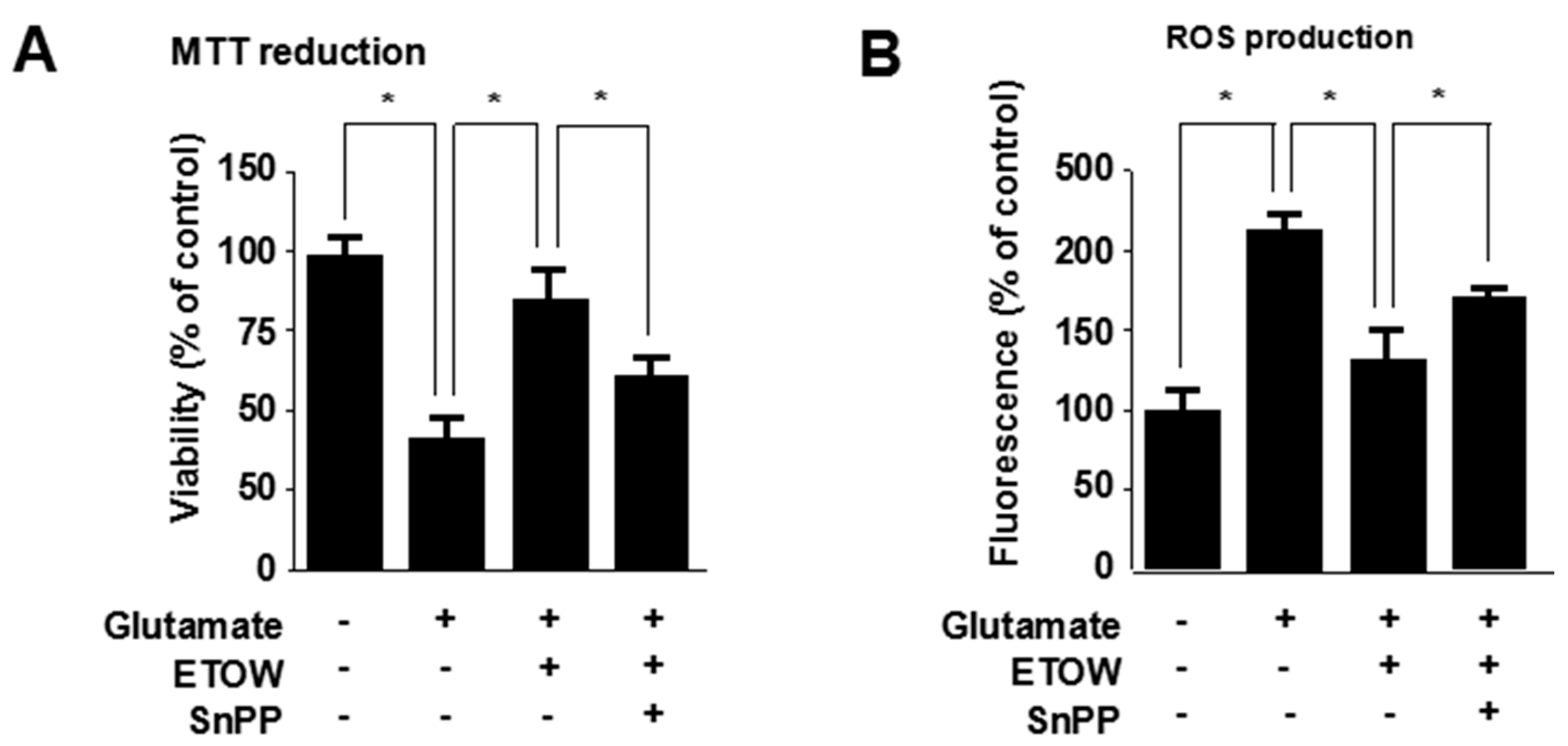
3.3. Effects of ETOW on Glutamate-Induced Cytotoxicity and Reactive Oxygen Species Production in HT22 Cells
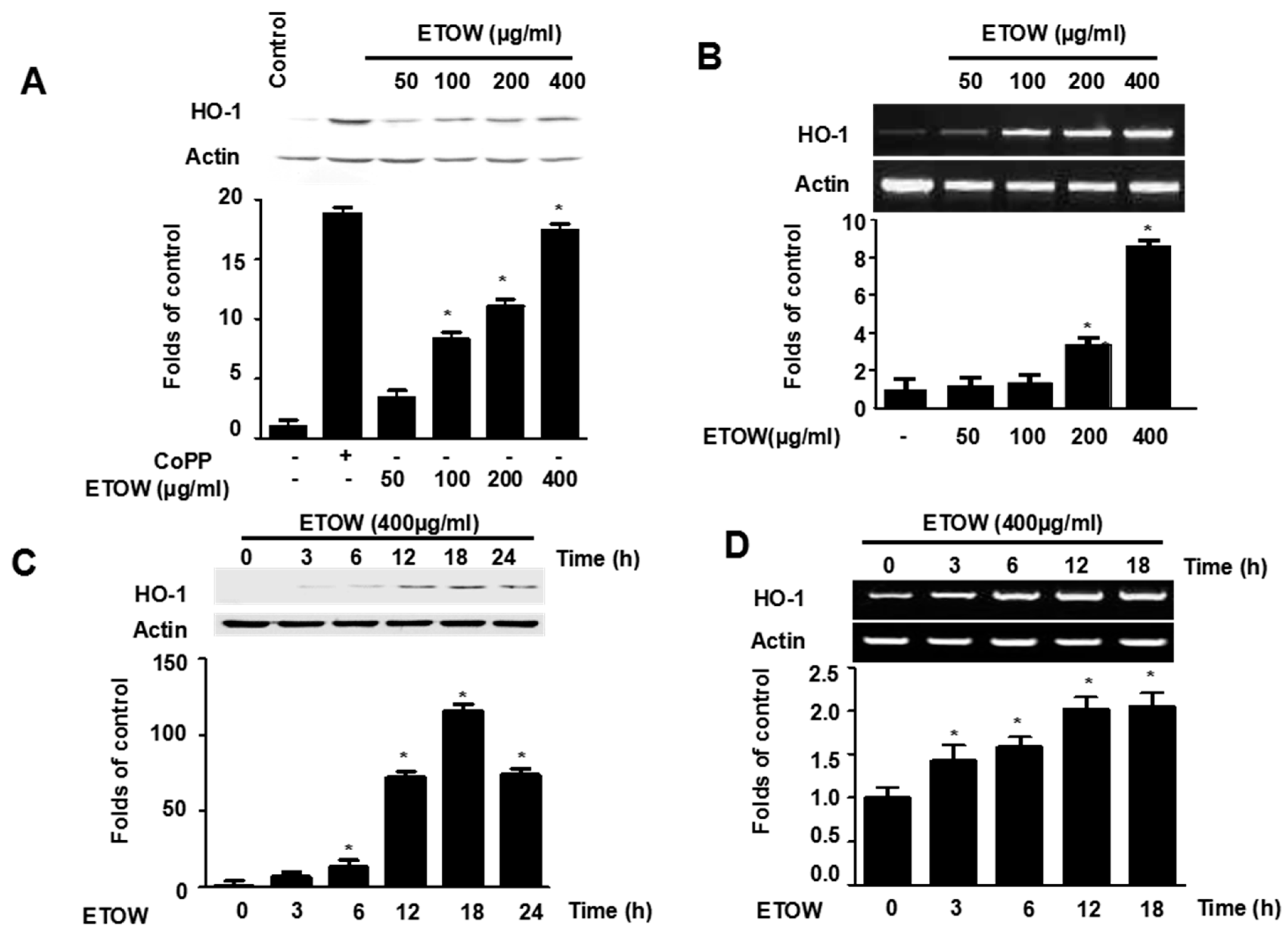
3.4. Effects of ETOW on Expression of HO-1 in HT22 Cells
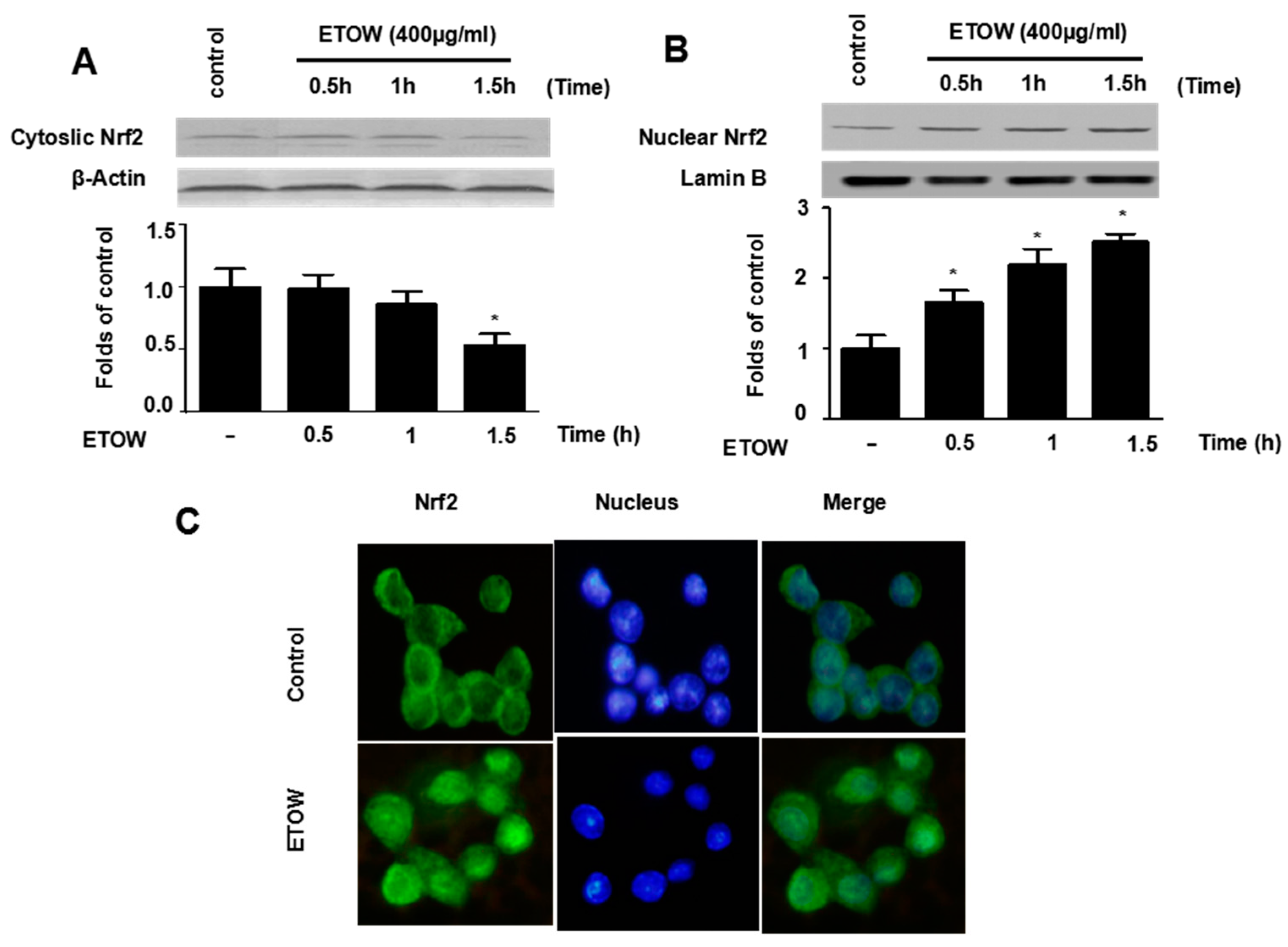
3.5. Effects of ETOW on Nrf2 Translocation in HT22 Cells
3.6. Effects of ETOW-Induced HO-1 Expression Pathway in Glutamate-Induced HT22 Cells
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ienco, E.C.; Logerfo, A.; Carlesi, C.; Orsucci, D.; Ricci, G.; Mancuso, M.; Siciliano, G. Oxidative stress treatment for clinical trials in neurodegenerative diseases. J. Alzheimers Dis. 2011, 24, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Barnham, K.J.; Masters, C.L.; Bush, A.I. Neurodegenerative diseases and oxidative stress. Biomed. Pharmacother. 2004, 58, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, M.; Giacomazza, D.; Picone, P.; Nuzzo, D.; San Biagio, P.L. Are oxidative stress and mitochondrial dysfunction the key players in the neurodegenerative diseases? Free Radic. Res. 2012, 46, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Guerraaraiza, C.; Álvarezmejía, A.L.; Sáncheztorres, S.; Farfangarcía, E.; Mondragónlozano, R.; Pintoalmazán, R.; Salgadoceballos, H. Effect of natural exogenous antioxidants on aging and on neurodegenerative diseases. Free Radic. Res. 2013, 47, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wuliji, O.; Li, W.; Jiang, Z.G.; Ghanbari, H.A. Oxidative Stress and Neurodegenerative Disorders. Int. J. Mol. Sci. 2013, 14, 24438–24475. [Google Scholar] [CrossRef] [PubMed]
- Solanki, I.; Parihar, P.; Mansuri, M.L.; Parihar, M.S. Flavonoid-Based Therapies in the Early Management of Neurodegenerative Diseases. Adv. Nutr. 2015, 6, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Peden, A.H.; Ironside, J.W. Molecular pathology in neurodegenerative diseases. Curr. Drug Targets 2012, 13, 1548–1559. [Google Scholar] [CrossRef] [PubMed]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative Stress in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [PubMed]
- Schipper, H.M. Heme oxygenase expression in human central nervous system disorders. Free Radic. Biol. Med. 2004, 37, 1995–2011. [Google Scholar] [CrossRef] [PubMed]
- Schipper Hyman, M.; Song, W.; Zukor, H.; Hascalovici Jacob, R.; Zeligman, D. Heme oxygenase-1 and neurodegeneration: Expanding frontiers of engagement. J. Neurochem. 2009, 110, 469–485. [Google Scholar] [CrossRef] [PubMed]
- Schipper, H.M. Heme oxygenase-1 in Alzheimer disease: A tribute to Moussa Youdim. J. Neural Transm. 2011, 118, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Schipper Hyman, M. Heme Oxygenase-1: Transducer of Pathological Brain Iron Sequestration under Oxidative Stress. Ann. N. Y. Acad. Sci. 2006, 1012, 84–93. [Google Scholar] [CrossRef]
- Chao, X.J.; Chen, Z.W.; Liu, A.M.; He, X.X.; Wang, S.G.; Wang, Y.T.; Liu, P.Q.; Ramassamy, C.; Mak, S.H.; Cui, W.; et al. Effect of Tacrine-3-caffeic Acid, A Novel Multifunctional Anti-Alzheimer’s Dimer, Against Oxidative-Stress-Induced Cell Death in HT22 Hippocampal Neurons: Involvement of Nrf2/HO-1 Pathway. CNS Neurosci. Ther. 2014, 20, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Barone, E.; Di Domenico, F.; Mancuso, C.; Butterfield, D.A. The Janus Face of the Heme Oxygenase/Biliverdin Reductase System in Alzheimer Disease: It’s Time for Reconciliation. Neurobiol. Dis. 2014, 62, 144–159. [Google Scholar] [CrossRef] [PubMed]
- Akanda, M.R.; Kim, M.-J.; Kim, I.-S.; Ahn, D.; Tae, H.-J.; Rahman, M.M.; Park, Y.-G.; Seol, J.-W.; Nam, H.-H.; Choo, B.-K.; et al. Neuroprotective Effects of Sigesbeckia pubescens Extract on Glutamate-Induced Oxidative Stress in HT22 Cells via Downregulation of MAPK/caspase-3 Pathways. Cell. Mol. Neurobiol. 2018, 38, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.; Carle, R.; Schieber, A. Taraxacum—A review on its phytochemical and pharmacological profile. J. Ethnopharmacol. 2006, 107, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Esatbeyoglu, T.; Obermair, B.; Dorn, T.; Siems, K.; Rimbach, G.; Birringer, M. Sesquiterpene Lactone Composition and Cellular Nrf2 Induction of Taraxacum officinale Leaves and Roots and Taraxinic Acid β-d-Glucopyranosyl Ester. J. Med. Food 2017, 20, 71–78. [Google Scholar] [CrossRef] [PubMed]
- González-Castejón, M.; García-Carrasco, B.; Fernández-Dacosta, R.; Dávalos, A.; Rodriguez-Casado, A. Reduction of Adipogenesis and Lipid Accumulation by Taraxacum officinale (Dandelion) Extracts in 3T3L1 Adipocytes: An in vitro Study. Phytother. Res. 2013, 28, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.; Petkova, N.; Tumbarski, J.; Dincheva, I.; Badjakov, I.; Denev, P.; Pavlov, A. GC-MS characterization of n-hexane soluble fraction from dandelion (Taraxacum officinale Weber ex F.H. Wigg.) aerial parts and its antioxidant and antimicrobial properties. Z. Naturforsch. C 2017, 73, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Davaatseren, M.; Hur, H.J.; Yang, H.J.; Hwang, J.-T.; Park, J.H.; Kim, H.-J.; Kim, M.J.; Kwon, D.Y.; Sung, M.J. Taraxacum official (dandelion) leaf extract alleviates high-fat diet-induced nonalcoholic fatty liver. Food Chem. Toxicol. 2013, 58, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.Y.; Cho, H.S.; Lee, J.J.; Lee, H.J.; Jun, S.Y.; Lee, J.H.; Song, H.H.; Choi, S.; Saloura, V.; Park, C.G. Novel TRAIL Sensitizer Taraxacum Officinale F.H. Wigg Enhances TRAIL-Induced Apoptosis in Huh7 Cells. Mol. Carcinog. 2016, 55, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Domitrović, R.; Jakovac, H.; Romić, Ž.; Rahelić, D.; Tadić, Ž. Antifibrotic activity of Taraxacum officinale root in carbon tetrachloride-induced liver damage in mice. J. Ethnopharmacol. 2010, 130, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.J.; Cha, D.S.; Ko, J.S.; Park, H.J.; Choi, H.D. Anti-inflammatory effect of Taraxacum officinale leaves on lipopolysaccharide-induced inflammatory responses in RAW 264.7 cells. J. Med. Food. 2010, 13, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kang, M.J.; Kim, M.J.; Kim, M.E.; Song, J.H.; Lee, Y.M.; Kim, J.I. Pancreatic lipase inhibitory activity of taraxacum officinale in vitro and in vivo. Nutr. Res. Pract. 2008, 2, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Shen, J.D.; Li, Y.Y.; Huang, Q. Antidepressant effects of the water extract from Taraxacum officinale leaves and roots in mice. Pharm. Biol. 2014, 52, 1028–1032. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Poirrier, P.; Chamy, R.; Prüfer, D.; Schulze-Gronover, C.; Jorquera, L.; Ruiz, G. Taraxacum officinale and related species—An ethnopharmacological review and its potential as a commercial medicinal plant. J. Ethnopharmacol. 2015, 169, 244–262. [Google Scholar] [CrossRef] [PubMed]
- Hfaiedh, M.; Brahmi, D.; Zourgui, L. Hepatoprotective effect of Taraxacum officinale leaf extract on sodium dichromate-induced liver injury in rats. Environ. Toxicol. 2014, 31, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Park, C.M.; Park, J.Y.; Noh, K.H.; Shin, J.H.; Song, Y.S. Taraxacum officinale Weber extracts inhibit LPS-induced oxidative stress and nitric oxide production via the NF-κB modulation in RAW 264.7 cells. J. Ethnopharmacol. 2011, 133, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Colle, D.; Arantes, L.P.; Gubert, P.; Da, L.S.; Athayde, M.L.; Teixeira Rocha, J.B.; Soares, F.A. Antioxidant properties of Taraxacum officinale leaf extract are involved in the protective effect against hepatoxicity induced by acetaminophen in mice. J. Med. Food 2012, 15, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Colle, D.; Arantes, L.P.; Rauber, R.; de Mattos, S.E.C.; Rocha, J.B.T.D.; Nogueira, C.W.; Soares, F.A.A. Antioxidant properties of Taraxacum officinale fruit extract are involved in the protective effect against cellular death induced by sodium nitroprusside in brain of rats. Pharm. Biol. 2012, 50, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Choi, U.-K.; Lee, O.-H.; Yim, J.H.; Cho, C.-W.; Rhee, Y.K.; Lim, S.-I.; Kim, Y.-C. Hypolipidemic and Antioxidant Effects of Dandelion (Taraxacum officinale) Root and Leaf on Cholesterol-Fed Rabbits. Int. J. Mol. Sci. 2010, 11, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.S.; Ko, W.; Lee, D.S.; Kim, D.C.; Kim, J.; Choi, M.; Beom, J.S.; An, R.B.; Oh, H.; Kim, Y.C. Taraxacum coreanum protects against glutamate-induced neurotoxicity through heme oxygenase-1 expression in mouse hippocampal HT22 cells. Mol. Med. Rep. 2017, 15, 2347–2352. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.; Kammerer, D.R.; Carle, R.; Schieber, A. Characterization of phenolic acids and flavonoids in dandelion (Taraxacum officinale WEB. ex WIGG.) root and herb by high-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Zhao, Y.; Zhou, H.; Zhang, Y.; Jiang, X.; Huang, K. Identification of antioxidants from Taraxacum mongolicum by high-performance liquid chromatography–diode array detection–radical-scavenging detection–electrospray ionization mass spectrometry and nuclear magnetic resonance experiments. J. Chromatogr. A 2008, 1209, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.Y.; Zhang, Y.P.; Zhou, H.H.; Huang, K.L.; Jiang, X.Y. Screening and identification of radical scavengers from Neo-Taraxacum siphonanthum by online rapid screening method and nuclear magnetic resonance experiments. J. Immunoass. Immunochem. 2010, 31, 233–249. [Google Scholar] [CrossRef] [PubMed]
- Mingarro, D.M.; Plaza, A.; Galán, A.; Vicente, J.A.; Martínez, M.P.; Acero, N. The effect of five Taraxacum species on in vitro and in vivo antioxidant and antiproliferative activity. Food Funct. 2015, 6, 2787–2793. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Jeong, G.S.; Kang, D.G.; Lee, H.S.; Kim, Y.C. Cytoprotective effects of lindenenyl acetate isolated from Lindera strychnifolia on mouse hippocampal HT22 cells. Eur. J. Pharmacol. 2009, 614, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hao, W.; Qin, Y.; Decker, Y.; Wang, X.; Burkart, M.; Schötz, K.; Menger, M.D.; Fassbender, K.; Liu, Y. Long-term treatment with Ginkgo biloba extract EGb 761 improves symptoms and pathology in a transgenic mouse model of Alzheimer’s disease. Brain Behav. Immun. 2015, 46, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.-H. Effects of Panax ginseng in Neurodegenerative Diseases. J. Ginseng Res. 2012, 36, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Z.; Li, W.P.; Zhang, W.; Yin, Y.Y.; Sun, X.X.; Zhou, S.S.; Xu, X.Q.; Tao, C.R. Protective Effect of Extract of Astragalus on Learning and Memory Impairments and Neurons Apoptosis Induced by Glucocorticoids in 12-Month Male Mice. Anat. Rec. 2011, 294, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. Glutamate: A truly functional amino acid. Amino Acids 2013, 45, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Danbolt, N.C. Glutamate as a neurotransmitter in the healthy brain. J. Neural Transm. 2014, 121, 799–817. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Jeong, G.S. Butein provides neuroprotective and anti-neuroinflammatory effects through Nrf2/ARE-dependent haem oxygenase 1 expression by activating the PI3K/Akt pathway. Br. J. Pharmacol. 2016, 173, 2894–2909. [Google Scholar] [CrossRef] [PubMed]
- Sukprasansap, M.; Chanvorachote, P.; Tencomnao, T. Cleistocalyx nervosum var. paniala berry fruit protects neurotoxicity against endoplasmic reticulum stress-induced apoptosis. Food Chem. Toxicol. 2017, 103, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.N. Simultaneous activation of Nrf2 and elevation of antioxidant compounds for reducing oxidative stress and chronic inflammation in human Alzheimer’s disease. Mech. Ageing Dev. 2016, 153, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications. Oxid. Med. Cell. Longev. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fujino, M.; Nishio, Y.; Ito, H.; Tanaka, T.; Li, X.-K. 5-Aminolevulinic acid regulates the inflammatory response and alloimmune reaction. Int. Immunopharmacol. 2016, 37, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed]
- Paine, A.; Eiz-Vesper, B.; Blasczyk, R.; Immenschuh, S. Signaling to heme oxygenase-1 and its anti-inflammatory therapeutic potential. Biochem. Pharmacol. 2010, 80, 1895–1903. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tentative Identification | M | tR (min) | m/z Traces(−) | m/z Traces(+) | MS2 |
|---|---|---|---|---|---|
| cis-Caftaric acid | 312 | 2.4–2.6 | 311.0283 | 335.0603 | MS2(−)[311.0285]: 149.0075, 179, 135.0441 |
| trans-Coutaric acid | 296 | 3.7–3.8 | 295.0349 | MS2(−)[295.0354]: 163.0370,119.0506, 113, 164 | |
| Ferulic acid | 194 | 4.26–4.3 | 217.1082 | MS2(+)[217.1082]: 217.1086, 144.0847 | |
| Esculetin | 178 | 4.45–4.50 | 177.0147 | ||
| 5-O-Caffeoylquinic acid | 354 | 4.55–4.57 | 353.0755 | MS2(−)[353.0750]: 191.051, 354.077, 179 | |
| Caffeic acid | 180 | 4.6 | 179.0313 | MS2(−)[179.0323]: 135 | |
| Chicoric acid | 474 | 6.4–6.49 | 473.0541 | MS2(−)[473.0563]: 311.029, 293, 179.033, 474.059, 149, 312, 294, 219.0252, 341.0544 | |
| Luteolin | 286 | 10.3–10.4 | 285.0308 | 287.0739 | |
| 11β,13-dihydro-taraxinic acid | 264 | 10.7 | 263.1202 | MS2(−)[263.1202]: 219.1327, 245.1085 | |
| Quercetin-3’,4’,7-trimethyl ether | 344 | 18.5–18.6 | 345 | MS2(+)[345.2637]: 345.2638 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Meng, N.; Liu, Z.; Guo, L.; Dong, L.; Li, B.; Ye, Q. Neuroprotective Effects of Taraxacum officinale Wigg. Extract on Glutamate-Induced Oxidative Stress in HT22 Cells via HO-1/Nrf2 Pathways. Nutrients 2018, 10, 926. https://doi.org/10.3390/nu10070926
Huang S, Meng N, Liu Z, Guo L, Dong L, Li B, Ye Q. Neuroprotective Effects of Taraxacum officinale Wigg. Extract on Glutamate-Induced Oxidative Stress in HT22 Cells via HO-1/Nrf2 Pathways. Nutrients. 2018; 10(7):926. https://doi.org/10.3390/nu10070926
Chicago/Turabian StyleHuang, Shan, Ning Meng, Zhiming Liu, Li Guo, Linsha Dong, Bin Li, and Qiang Ye. 2018. "Neuroprotective Effects of Taraxacum officinale Wigg. Extract on Glutamate-Induced Oxidative Stress in HT22 Cells via HO-1/Nrf2 Pathways" Nutrients 10, no. 7: 926. https://doi.org/10.3390/nu10070926
APA StyleHuang, S., Meng, N., Liu, Z., Guo, L., Dong, L., Li, B., & Ye, Q. (2018). Neuroprotective Effects of Taraxacum officinale Wigg. Extract on Glutamate-Induced Oxidative Stress in HT22 Cells via HO-1/Nrf2 Pathways. Nutrients, 10(7), 926. https://doi.org/10.3390/nu10070926