Lightly Cooked Broccoli Is as Effective as Raw Broccoli in Mitigating Dextran Sulfate Sodium-Induced Colitis in Mice


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Preparation
2.2. Animal Use and Experimental Design
2.3. Diet Analysis
2.4. Tissue Collection
2.5. Disease Activity Index
2.6. Plasma LPS Determination
2.7. Urinary Sucralose Determination
2.8. mRNA Expression of Tight Junction Proteins and Pro-Inflammatory Cytokines
2.9. Protein Expression of Tight Junctions
2.10. Histology
2.11. Statistical Analysis
3. Results
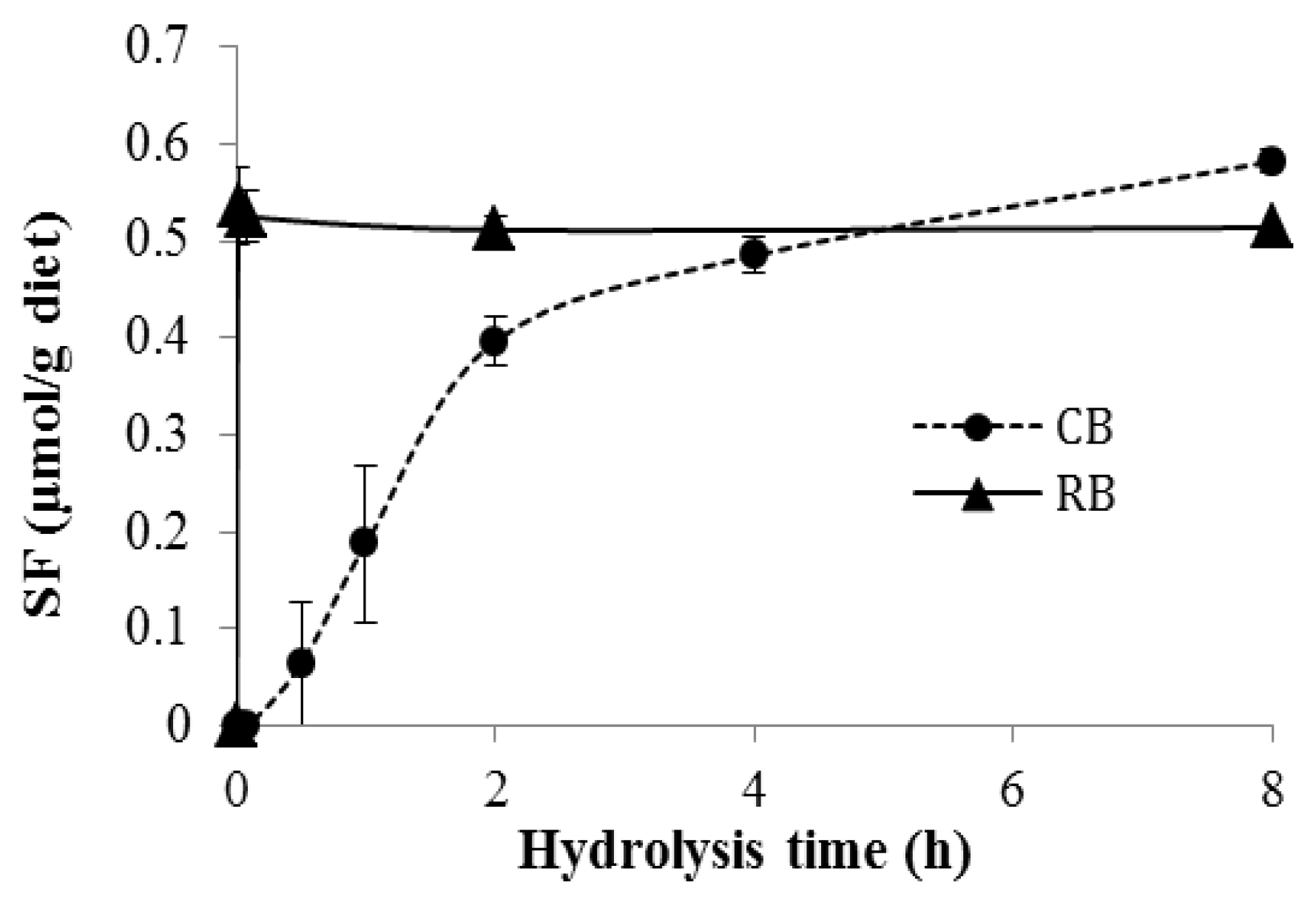
3.1. Formation of SF during Diet Hydrolysis In Vitro
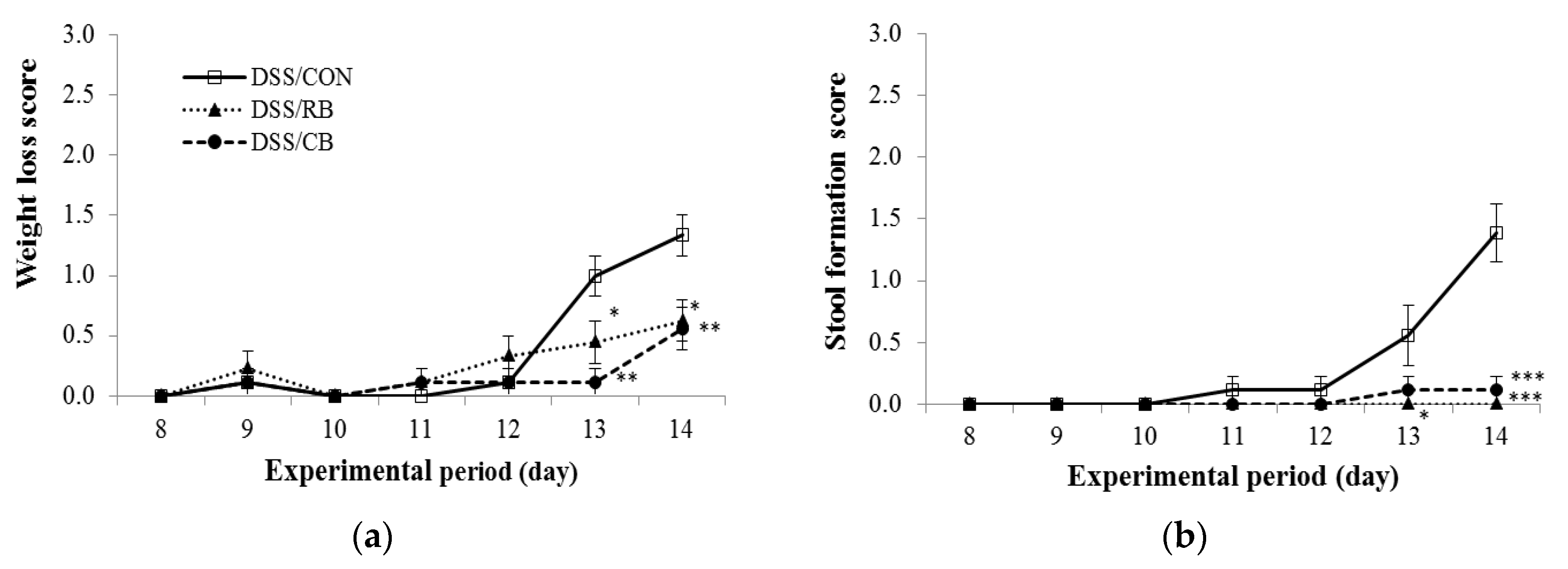
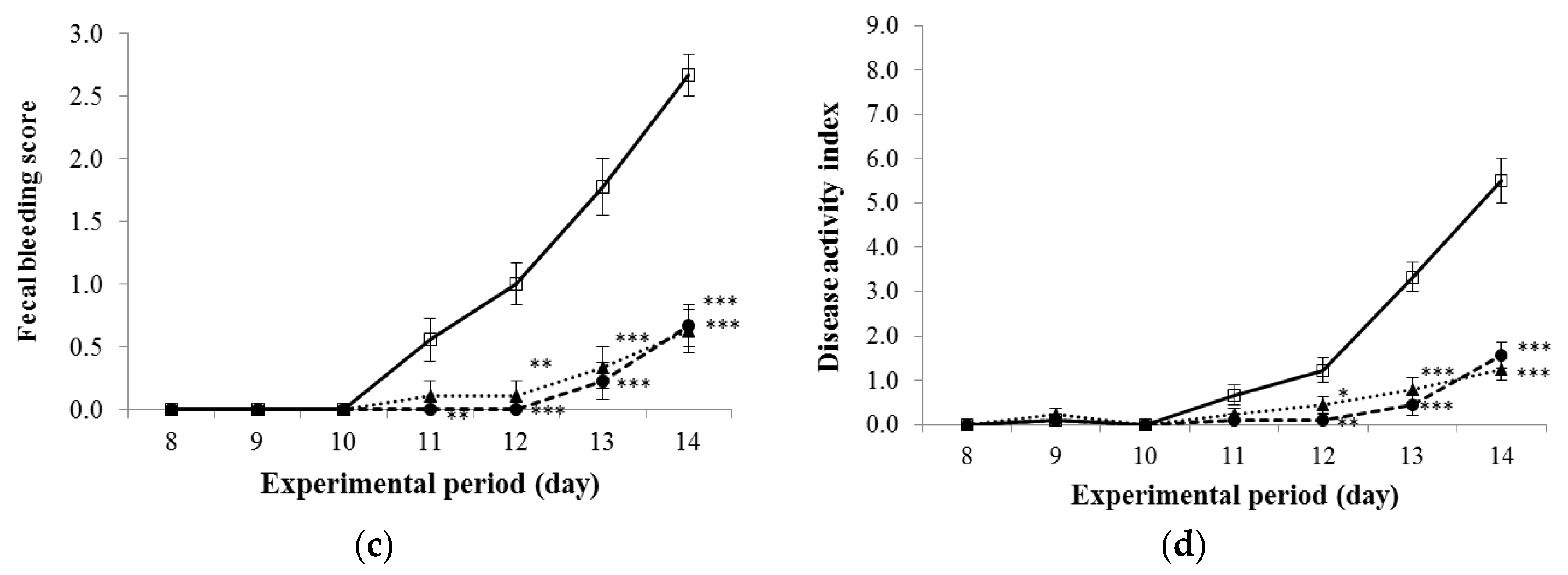
3.2. Disease Activity Index
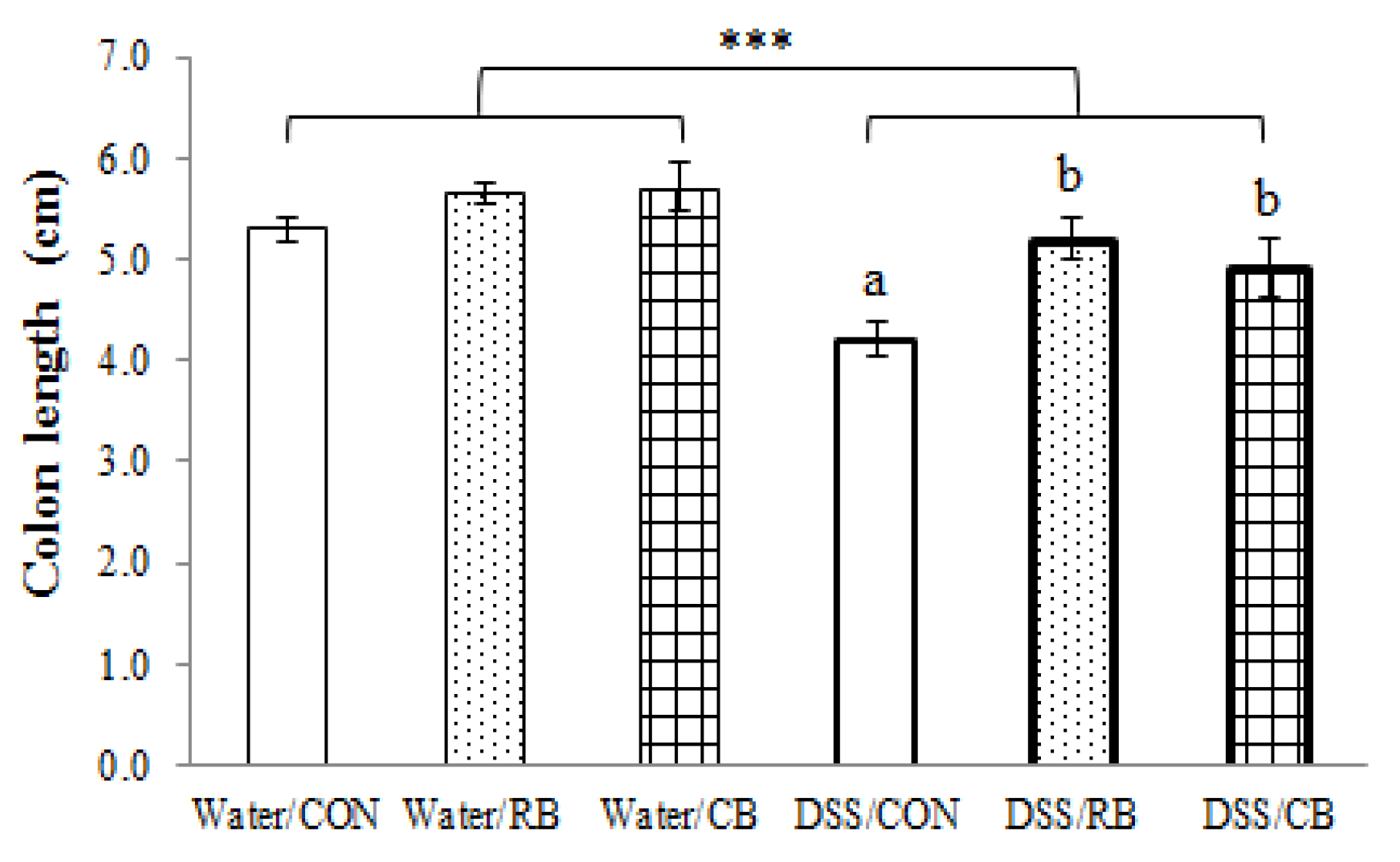
3.3. Colon Length
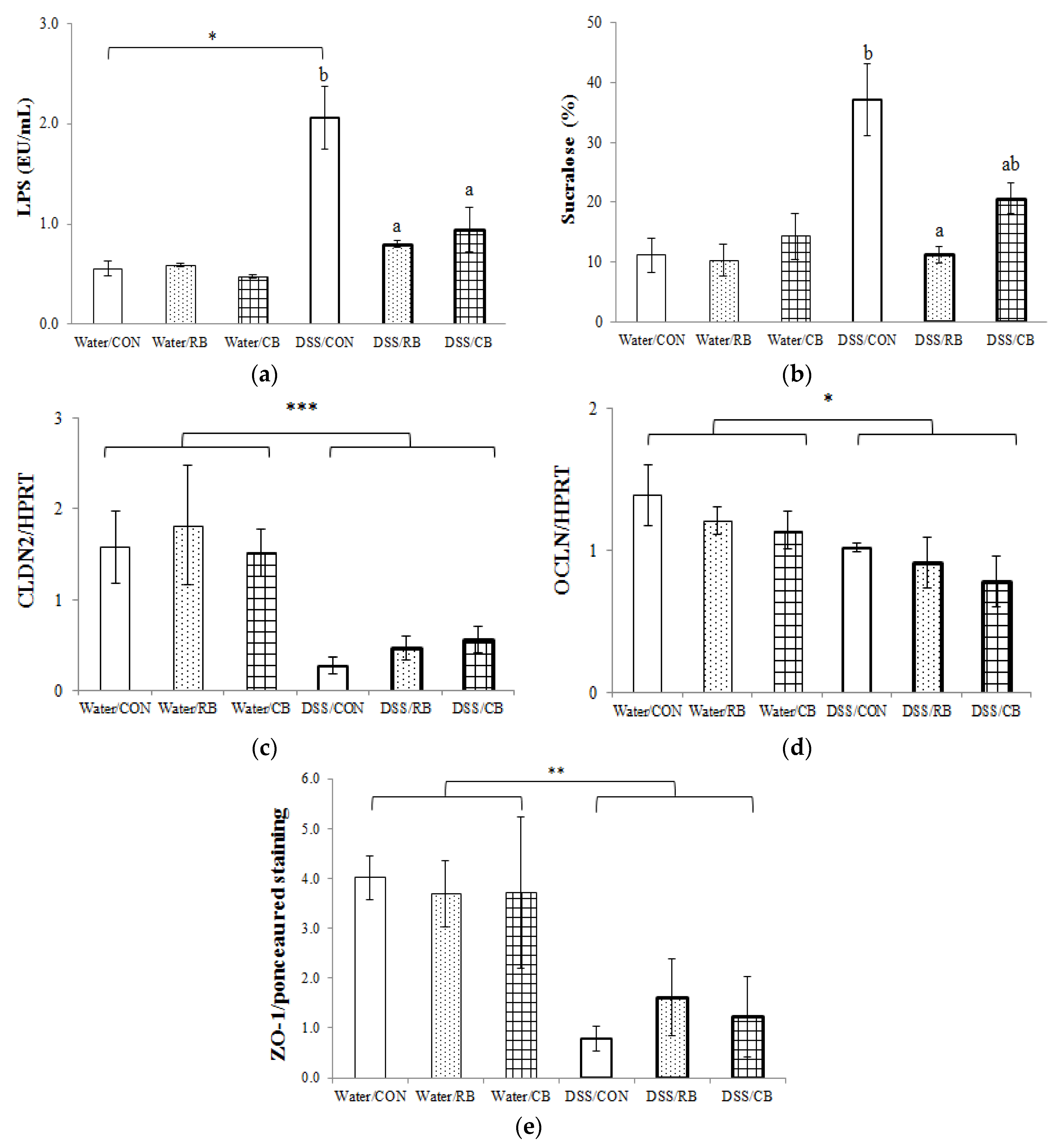
3.4. Gut Barrier Permeability
3.5. Tight Junction Expression
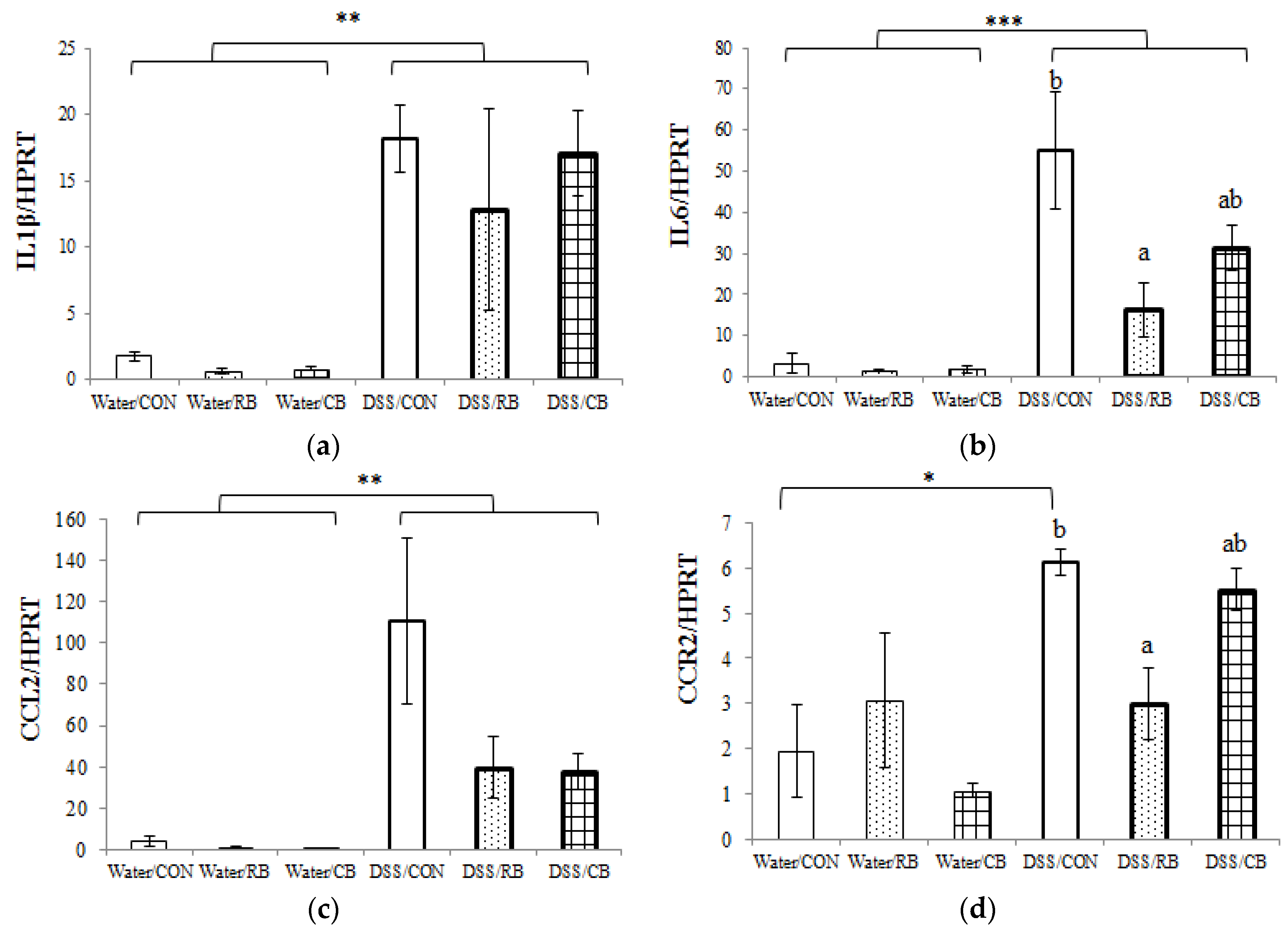
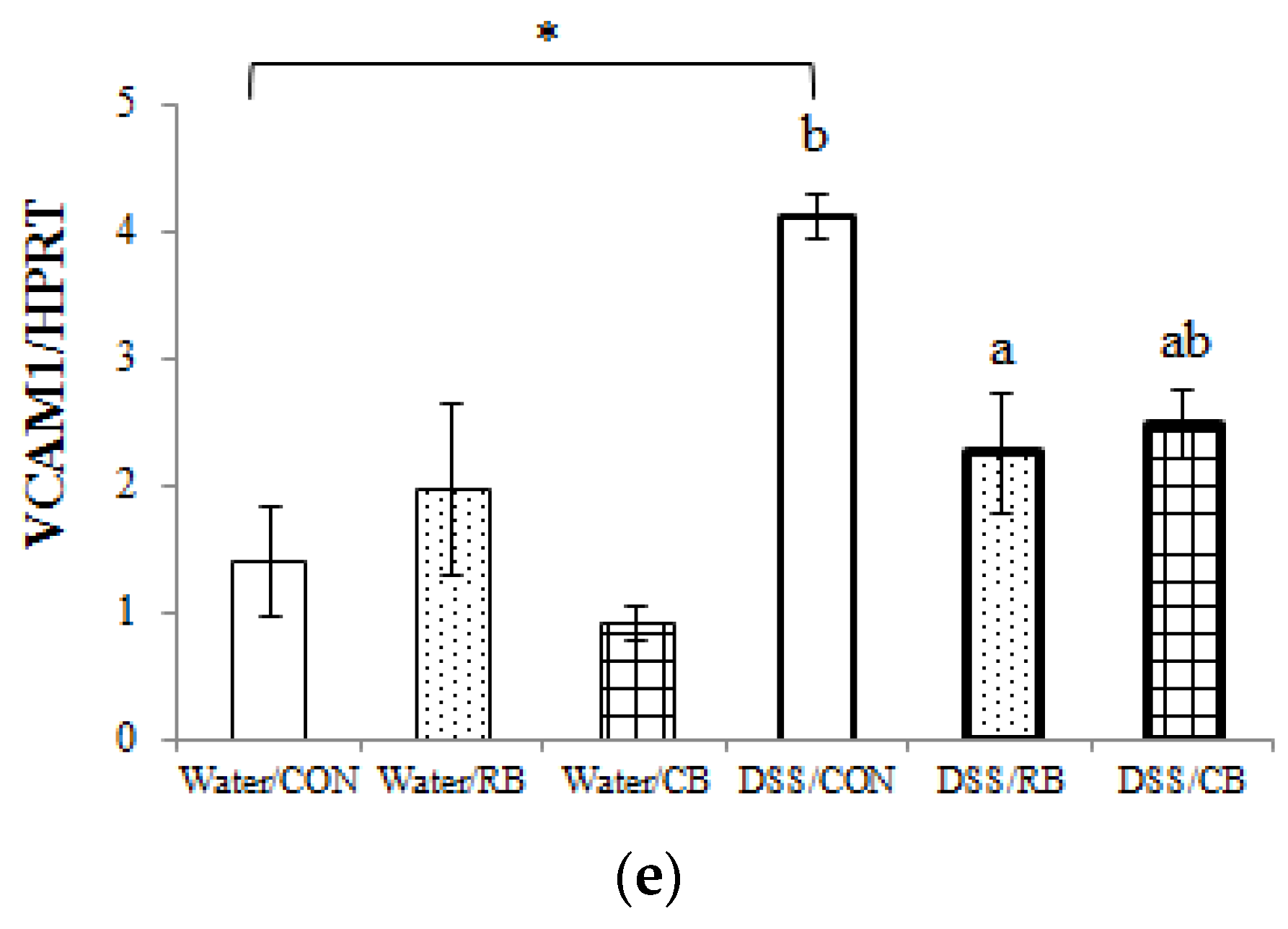
3.6. mRNA Expression of Pro-Inflammatory Cytokines
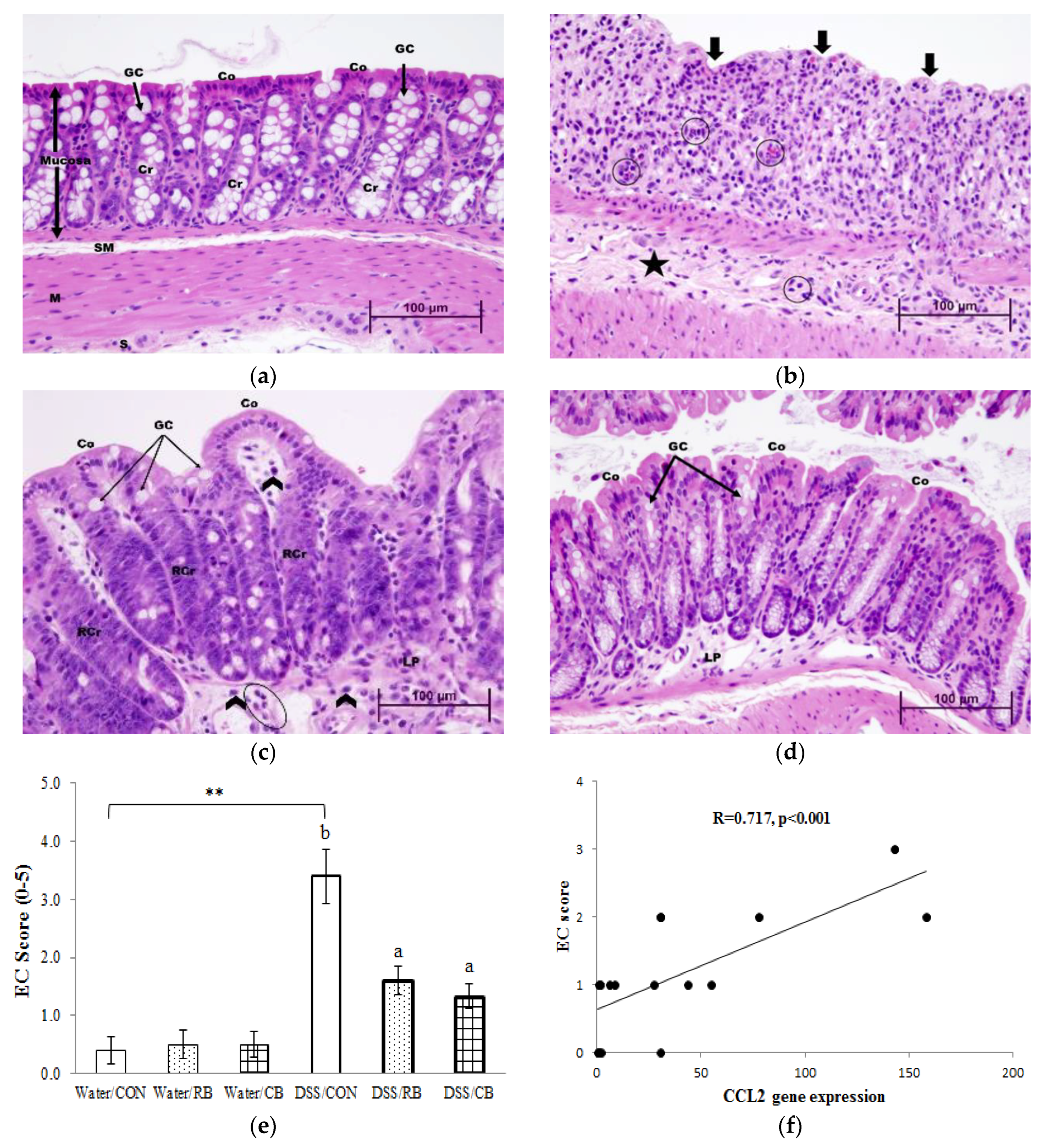
3.7. Histology
4. Discussion
4.1. Site of SF Absorption
4.2. Protection from DSS Colitis: Comparing CB and RB
4.3. Gut Barrier Integrity
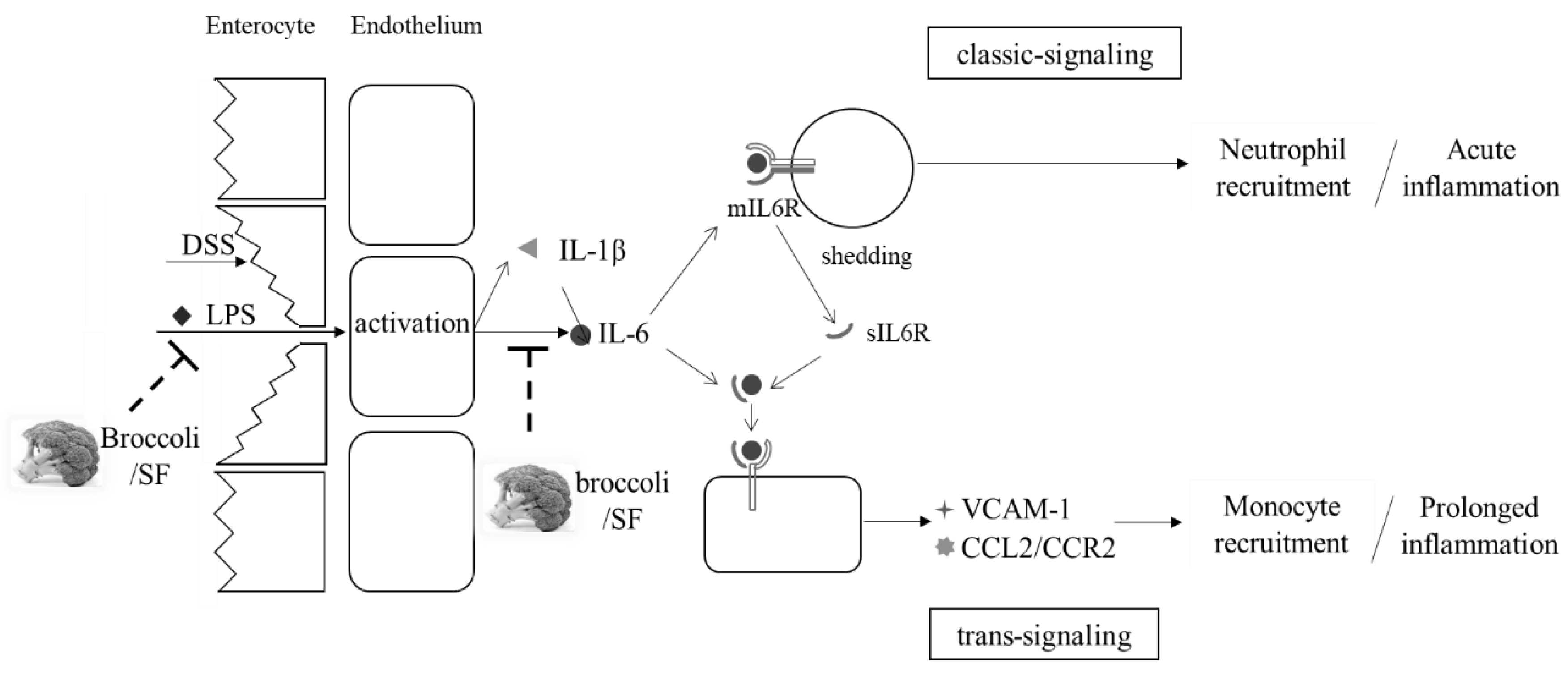
4.4. Inflammation
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Dahlhamer, J.M.; Zammitti, E.P.; Ward, B.W.; Wheaton, A.G.; Croft, J.B. Prevalence of Inflammatory Bowel Disease Among Adults Aged ≥18 Years—United States, 2015. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 1166–1169. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.E.; Will, O.; Sturm, C.; Lipinski, S.; Rosenstiel, P.; Rimbach, G. DSS-induced acute colitis in C57BL/6 mice is mitigated by sulforaphane pre-treatment. J. Nutr. Biochem. 2013, 24, 2085–2091. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.-O.; Kim, M.-O.; Kang, S.-H.; Choi, Y.H.; Kim, G.-Y. Sulforaphane suppresses TNF-α-mediated activation of NF-κB and induces apoptosis through activation of reactive oxygen species-dependent caspase-3. Cancer Lett. 2009, 274, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, H.; Soroka, Y.; Zlotkin-Frušić, M.; Gilhar, A.; Milner, Y.; Tamir, S. Isothiocyanates inhibit psoriasis-related proinflammatory factors in human skin. Inflamm. Res. 2012, 61, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, E.H.; Araya, M. Physiological effects of broccoli consumption. Phytochem. Rev. 2009, 8, 283–298. [Google Scholar] [CrossRef]
- Paturi, G.; Mandimika, T.; Butts, C.A.; Zhu, S.; Roy, N.C.; McNabb, W.C.; Ansell, J. Influence of dietary blueberry and broccoli on cecal microbiota activity and colon morphology in mdr1a−/− mice, a model of inflammatory bowel diseases. Nutrition 2012, 28, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Lippmann, D.; Lehmann, C.; Florian, S.; Barknowitz, G.; Haack, M.; Mewis, I.; Wiesner, M.; Schreiner, M.; Glatt, H.; Brigelius-Flohé, R.; et al. Glucosinolates from pak choi and broccoli induce enzymes and inhibit inflammation and colon cancer differently. Food Funct. 2014, 5, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Lai, R.-H.; Miller, M.J.; Jeffery, E. Glucoraphanin hydrolysis by microbiota in the rat cecum results in sulforaphane absorption. Food Funct. 2010, 1, 161. [Google Scholar] [CrossRef] [PubMed]
- Rungapamestry, V.; Duncan, A.J.; Fuller, Z.; Ratcliffe, B. Effect of meal composition and cooking duration on the fate of sulforaphane following consumption of broccoli by healthy human subjects. Br. J. Nutr. 2007, 97, 644. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.C.; Farnham, M.; Jeffery, E.H. Impact of Thermal Processing on Sulforaphane Yield from Broccoli (Brassica oleracea L. ssp. italica). J. Agric. Food Chem. 2012, 60, 6743–6748. [Google Scholar] [CrossRef] [PubMed]
- Poritz, L.S.; Garver, K.I.; Green, C.; Fitzpatrick, L.; Ruggiero, F.; Koltun, W.A. Loss of the Tight Junction Protein ZO-1 in Dextran Sulfate Sodium Induced Colitis. J. Surg. Res. 2007, 140, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.V.; Gustafsson, J.K.; Holmén-Larsson, J.; Jabbar, K.S.; Xia, L.; Xu, H.; Ghishan, F.K.; Carvalho, F.A.; Gewirtz, A.T.; Sjövall, H.; et al. Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis. Gut 2014, 63, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Lambert, G.P. Stress-induced gastrointestinal barrier dysfunction and its inflammatory effects1. J. Anim. Sci. 2009, 87, E101–E108. [Google Scholar] [CrossRef] [PubMed]
- Dosz, E.B.; Ku, K.-M.; Juvik, J.A.; Jeffery, E.H. Total Myrosinase Activity Estimates in Brassica Vegetable Produce. J. Agric. Food Chem. 2014, 62, 8094–8100. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Shajib, M.S.; Manocha, M.M.; Khan, W.I. Investigating Intestinal Inflammation in DSS-induced Model of IBD. J. Vis. Exp. 2012. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, M.; Rajan, K.; Forsyth, C.B.; Voigt, R.M.; Keshavarzian, A. Simultaneous gas-chromatographic urinary measurement of sugar probes to assess intestinal permeability: Use of time course analysis to optimize its use to assess regional gut permeability. Clin. Chim. Acta 2015, 442, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Viennois, E.; Chen, F.; Laroui, H.; Baker, M.T.; Merlin, D. Dextran sodium sulfate inhibits the activities of both polymerase and reverse transcriptase: lithium chloride purification, a rapid and efficient technique to purify RNA. BMC Res. Notes 2013, 6, 360. [Google Scholar] [CrossRef] [PubMed]
- Romero-Calvo, I.; Ocón, B.; Martínez-Moya, P.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; de Medina, F.S. Reversible Ponceau staining as a loading control alternative to actin in Western blots. Anal. Biochem. 2010, 401, 318–320. [Google Scholar] [CrossRef] [PubMed]
- Sirois, I.; Raymond, M.-A.; Brassard, N.; Cailhier, J.-F.; Fedjaev, M.; Hamelin, K.; Londono, I.; Bendayan, M.; Pshezhetsky, A.V.; Hébert, M.-J. Caspase-3-dependent export of TCTP: A novel pathway for antiapoptotic intercellular communication. Cell Death Differ. 2011, 18, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Hussein, H.; Wang, H.; Rabbi, M.F.; Bernstein, C.N.; Ghia, J.-E. Stability of Reference Genes for Messenger RNA Quantification by Real-Time PCR in Mouse Dextran Sodium Sulfate Experimental Colitis. PLoS ONE 2016, 11, e0156289. [Google Scholar] [CrossRef] [PubMed]
- Dieleman, L.A.; Ridwan, B.U.; Tennyson, G.S.; Beagley, K.W.; Bucy, R.P.; Elson, C.O. Dextran sulfate sodium-induced colitis occurs in severe combined immunodeficient mice. Gastroenterology 1994, 107, 1643–1652. [Google Scholar] [CrossRef]
- Wang, Y.; Jeffery, E.H.; Miller, M.J.; Wallig, M.A.; Wu, Y. (University of Illinois, Urbana, IL, USA). Unpublished work. 2018. [Google Scholar]
- Matusheski, N.V.; Jeffery, E.H. Comparison of the Bioactivity of Two Glucoraphanin Hydrolysis Products Found in Broccoli, Sulforaphane and Sulforaphane Nitrile. J. Agric. Food Chem. 2001, 49, 5743–5749. [Google Scholar] [CrossRef] [PubMed]
- Rungapamestry, V.; Duncan, A.J.; Fuller, Z.; Ratcliffe, B. Changes in Glucosinolate Concentrations, Myrosinase Activity, and Production of Metabolites of Glucosinolates in Cabbage (Brassica oleracea Var. capitata ) Cooked for Different Durations. J. Agric. Food Chem. 2006, 54, 7628–7634. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, P.; Grosse, J.; Asad, A.B.M.A.; Radda, G.K.; Golay, X. Gastrointestinal transit measurements in mice with 99mTc-DTPA-labeled activated charcoal using NanoSPECT-CT. EJNMMI Res. 2013, 3, 60. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Hoeflinger, J.L.; Neme, B.P.; Jeffery, E.H.; Miller, M.J. Dietary Broccoli Alters Rat Cecal Microbiota to Improve Glucoraphanin Hydrolysis to Bioactive Isothiocyanates. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, S.R.; Britton, G.J.; Contijoch, E.J.; Vennaro, O.H.; Mortha, A.; Colombel, J.-F.; Grinspan, A.; Clemente, J.C.; Merad, M.; Faith, J.J. Interactions Between Diet and the Intestinal Microbiota Alter Intestinal Permeability and Colitis Severity in Mice. Gastroenterology 2018, 154, 1037–1046.e2. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Niioka, M.; Kobayashi, N.; Tanaka, M.; Watanabe, T. Butyrate-Producing Probiotics Reduce Nonalcoholic Fatty Liver Disease Progression in Rats: New Insight into the Probiotics for the Gut-Liver Axis. PLoS ONE 2013, 8, e63388. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Bibi, S.; Du, M.; Suzuki, T.; Zhu, M.-J. Regulation of the intestinal tight junction by natural polyphenols: A mechanistic perspective. Crit. Rev. Food Sci. Nutr. 2017, 57, 3830–3839. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-T.; Xu, Y.-F.; Huang, Y.-F.; Qu, C.; Xu, L.-Q.; Su, Z.-R.; Zeng, H.-F.; Zheng, L.; Yi, T.-G.; Li, H.-L.; et al. Berberrubine attenuates mucosal lesions and inflammation in dextran sodium sulfate-induced colitis in mice. PLoS ONE 2018, 13, e0194069. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8 (Suppl. 2), S3. [Google Scholar] [CrossRef] [PubMed]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta BBA—Mol. Cell Res. 2011, 1813, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. IL-6 Trans-Signaling via the Soluble IL-6 Receptor: Importance for the Pro-Inflammatory Activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Mudter, J.; Neurath, M.F. Il-6 signaling in inflammatory bowel disease: Pathophysiological role and clinical relevance. Inflamm. Bowel Dis. 2007, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Kaplanski, G. IL-6: A regulator of the transition from neutrophil to monocyte recruitment during inflammation. Trends Immunol. 2003, 24, 25–29. [Google Scholar] [CrossRef]
- Saji, H.; Koike, M.; Yamori, T.; Saji, S.; Seiki, M.; Matsushima, K.; Toi, M. Significant correlation of monocyte chemoattractant protein-1 expression with neovascularization and progression of breast carcinoma. Cancer 2001, 92, 1085–1091. [Google Scholar] [CrossRef]
- Fujimoto, H.; Sangai, T.; Ishii, G.; Ikehara, A.; Nagashima, T.; Miyazaki, M.; Ochiai, A. Stromal MCP-1 in mammary tumors induces tumor-associated macrophage infiltration and contributes to tumor progression. Int. J. Cancer 2009, 125, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/100 g Diet) | AIN-93M (CON) | 10% Raw Broccoli (RB) | 10% Lightly Cooked Broccoli (CB) |
|---|---|---|---|
| Freeze-dried raw broccoli powder | 0 | 10.0 | 0 |
| Freeze-dried lightly cooked broccoli powder | 0.0 | 0 | 10.0 |
| Casein | 14.0 | 11.4 | 11.4 |
| Cornstarch | 46.6 | 44.4 | 44.4 |
| Maltodextrin | 15.5 | 14.6 | 14.6 |
| Sucrose | 10.0 | 9.3 | 9.3 |
| Cellulose | 5.0 | 2.6 | 2.6 |
| Mineral mix | 3.5 | 2.7 | 2.7 |
| Vitamin mix | 1.0 | 1.0 | 1.0 |
| L-cysteine | 0.2 | 0.2 | 0.2 |
| Choline bitartrate | 0.3 | 0.3 | 0.3 |
| Soybean oil | 4.0 | 4.0 | 4.0 |
| Gene | NCBI Reference Sequence | Forward Primer | Reverse Primer |
|---|---|---|---|
| Tight Junction | |||
| CLDN1 | NM_016674.4 | TGTGGATGGCTGTCATTGGGG | ATTCATACCTGGCATTGATGGGGG |
| CLDN2 | NM_016675.4 | ACGGCTCCGTTTTCTAGATGCC | CGTTTGGCTGCTGCTCTTGC |
| CLDN3 | NM_009902.4 | AGCCCTCATCGTGGTGTCCA | GGCCGTCTCGTCTTGTACGC |
| CLDN4 | NM_009903.2 | GAGCCGTGTTCATCGTGGCA | CCCAGCCGACGTAAAGCGAG |
| CLDN5 | NM_013805.4 | GTGCGTGGTGCAGAGTACCG | GAGCGCCGGTCAAGGTAACA |
| CLDN8 | NM_018778.3 | ATGCACGGGGGACGATGAGA | TGAGCACAACCAAGCCGGTG |
| OCLN | NM_008756.2 | ACGGTCCTCCTGGCTCAGTT | GATAAGCGAACCTTGGCGGC |
| TJP1 | NM_001163574.1 | TGTTTATGCGGACGGTGGCG | TCCATTGCTGTGCTCTTAGCGG |
| Inflammation | |||
| CCL2 | NM_011333.3 | TTAAAAACCTGGATCGGAACCAA | GCATTAGCTTCAGATTTACGGGT |
| CCR2 | NM_009915.2 | ATAAAGGAGCCATACCTGTAAATGC | CATGTGGTGAATCCAATGCCCT |
| IL1B | NM_008361.4 | TGCCACCTTTTGACAGTGATGAGA | TGTTGATGTGCTGCTGCGAGA |
| IL6 | NM_031168.2 | TAGTCCTTCCTACCCCAATTTCC | TTGGTCCTTAGCCACTCCTTC |
| TLR2 | NM_011905.3 | AGGAGGTGCGGACTGTTTCCT | ATTTGACGCTTTGTCTGAGGTTTCG |
| TLR4 | NM_021297.3 | TCCCTGCATAGAGGTAGTTCCTA | TTCAAGGGGTTGAAGCTCAGA |
| TNF | NM_001278601.1 | TCGGTCCCCAAAGGGATGAGA | GGTGGTTTGTGAGTGTGAGGGT |
| VCAM1 | NM_011693.3 | ACGTGGACATCTACTCTTTCCCCA | CTTGACCGTGACCGGCTTCC |
| NFKB1 | NM_008689.2 | CTGCCATGTCTGCTGCTGCT | CGTGGGCATCACCCTCCAGA |
| Housekeeping | |||
| HPRT | NM_013556.2 | TCCCAGCGTCGTGATTAGCG | TCGAGCAAGTCTTTCAGTCCTGT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Jeffery, E.H.; Miller, M.J.; Wallig, M.A.; Wu, Y. Lightly Cooked Broccoli Is as Effective as Raw Broccoli in Mitigating Dextran Sulfate Sodium-Induced Colitis in Mice. Nutrients 2018, 10, 748. https://doi.org/10.3390/nu10060748
Wang Y, Jeffery EH, Miller MJ, Wallig MA, Wu Y. Lightly Cooked Broccoli Is as Effective as Raw Broccoli in Mitigating Dextran Sulfate Sodium-Induced Colitis in Mice. Nutrients. 2018; 10(6):748. https://doi.org/10.3390/nu10060748
Chicago/Turabian StyleWang, Yanling, Elizabeth H. Jeffery, Michael J. Miller, Matthew A. Wallig, and Yuanfeng Wu. 2018. "Lightly Cooked Broccoli Is as Effective as Raw Broccoli in Mitigating Dextran Sulfate Sodium-Induced Colitis in Mice" Nutrients 10, no. 6: 748. https://doi.org/10.3390/nu10060748
APA StyleWang, Y., Jeffery, E. H., Miller, M. J., Wallig, M. A., & Wu, Y. (2018). Lightly Cooked Broccoli Is as Effective as Raw Broccoli in Mitigating Dextran Sulfate Sodium-Induced Colitis in Mice. Nutrients, 10(6), 748. https://doi.org/10.3390/nu10060748