Hepatic Mitochondrial Dysfunction and Immune Response in a Murine Model of Peanut Allergy




Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials
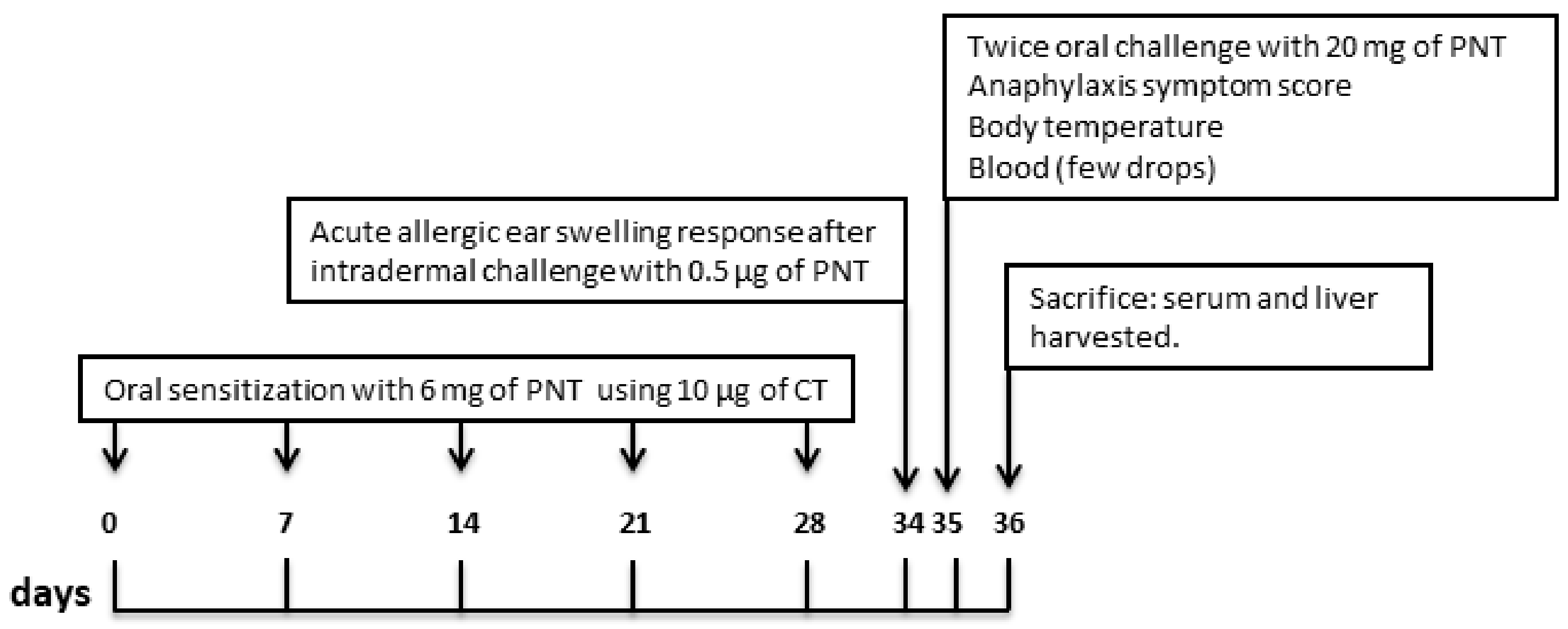
2.3. Sensitization Protocol
2.4. Acute Allergic Skin Response, Anaphylaxis Symptom Score, Body Temperature and mMCP-1 Serum Level
2.5. Serum Anti-Peanuts Immunoglobulin E
2.6. Cytokines Gene Expression Analysis by Quantitative Real Time PCR
2.7. Preparation of Isolated Mitochondria
2.8. Oxidative Capacity
2.9. Determination of Superoxide Dismutase (SOD) and Aconitase Enzymatic Activities and H2O2 Release
2.10. Statistical Analysis
3. Results
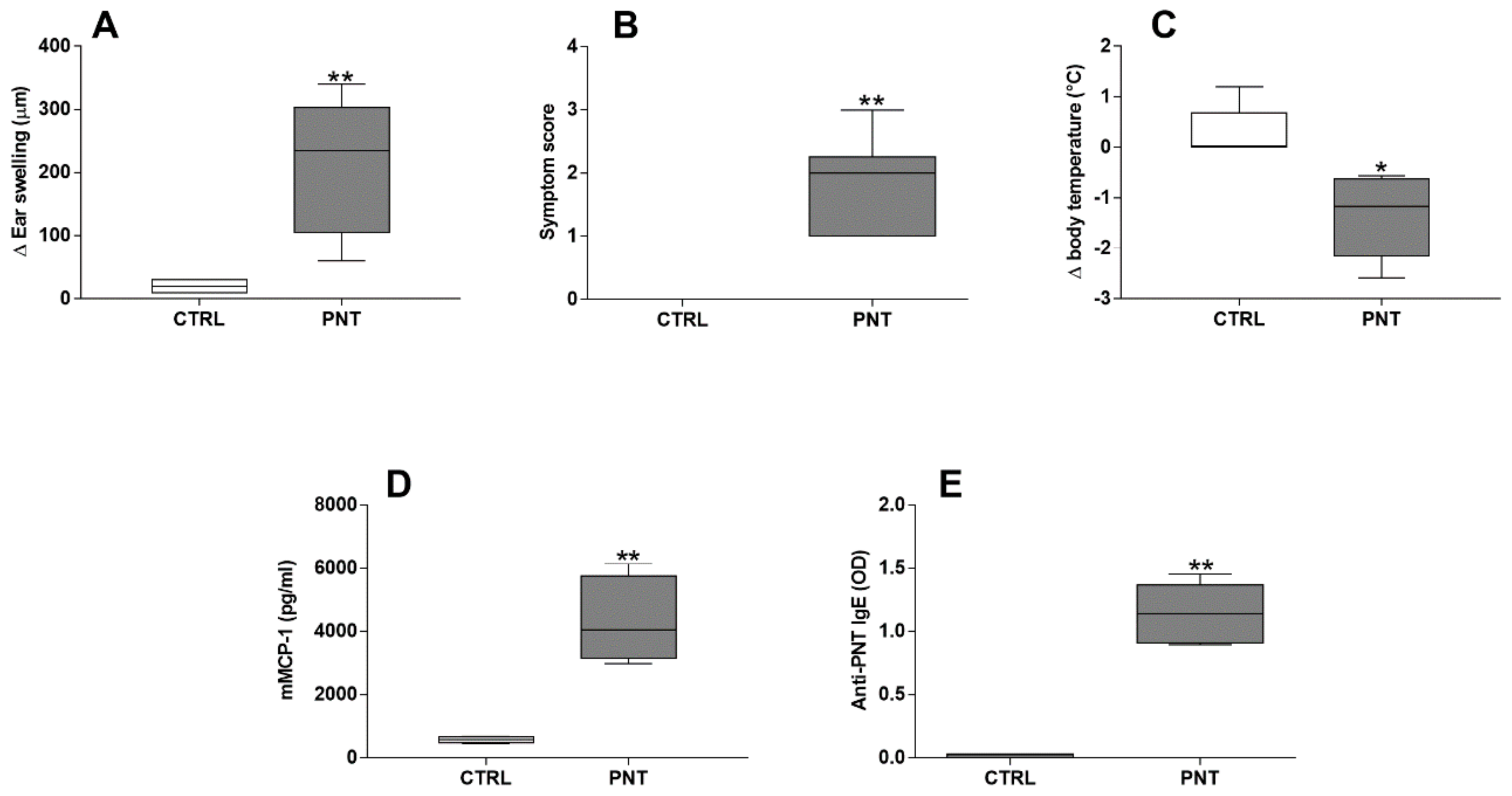
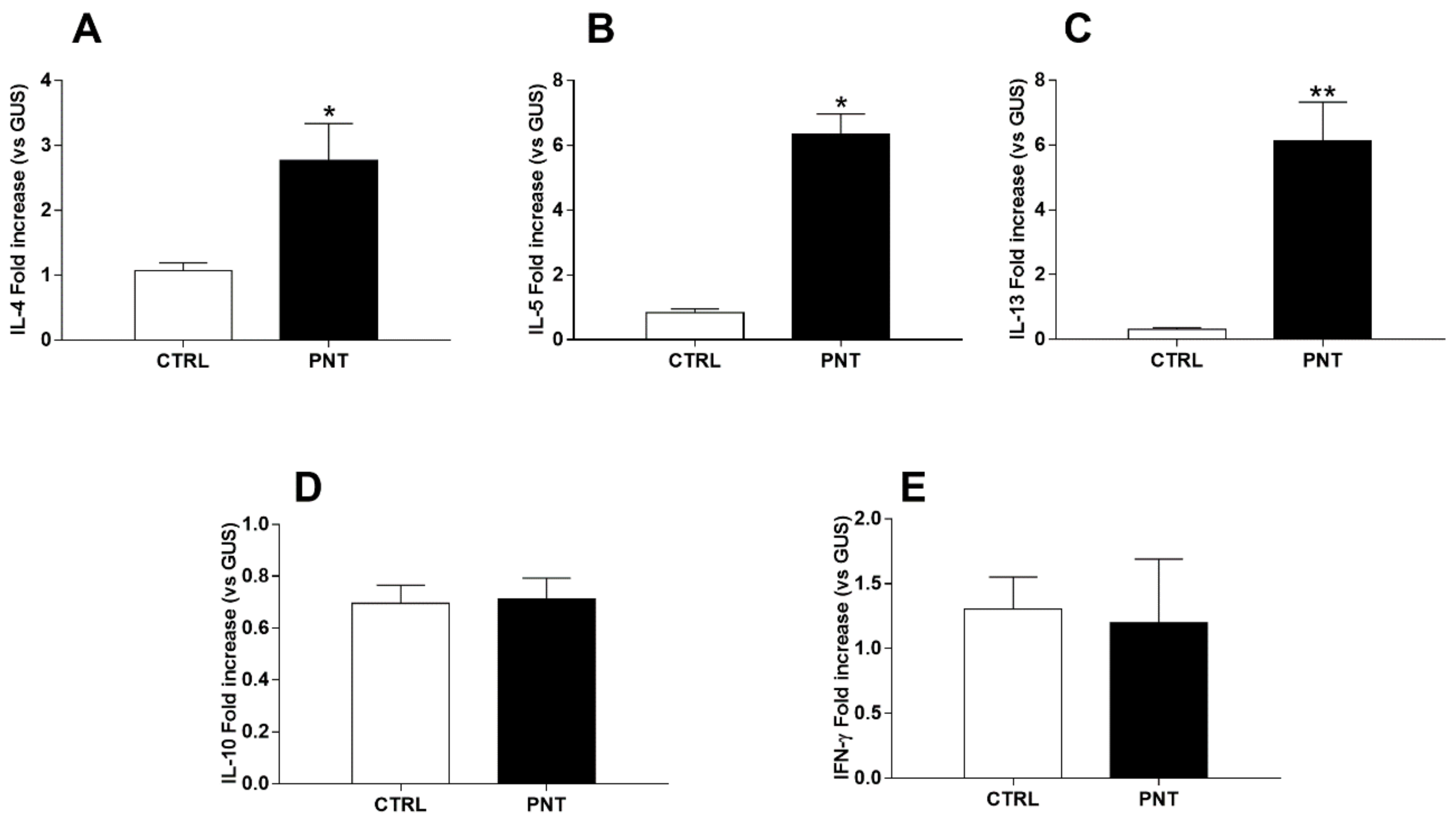
3.1. Allergic Response and Liver Involvement in Peanut Extract-Sensitized Mice
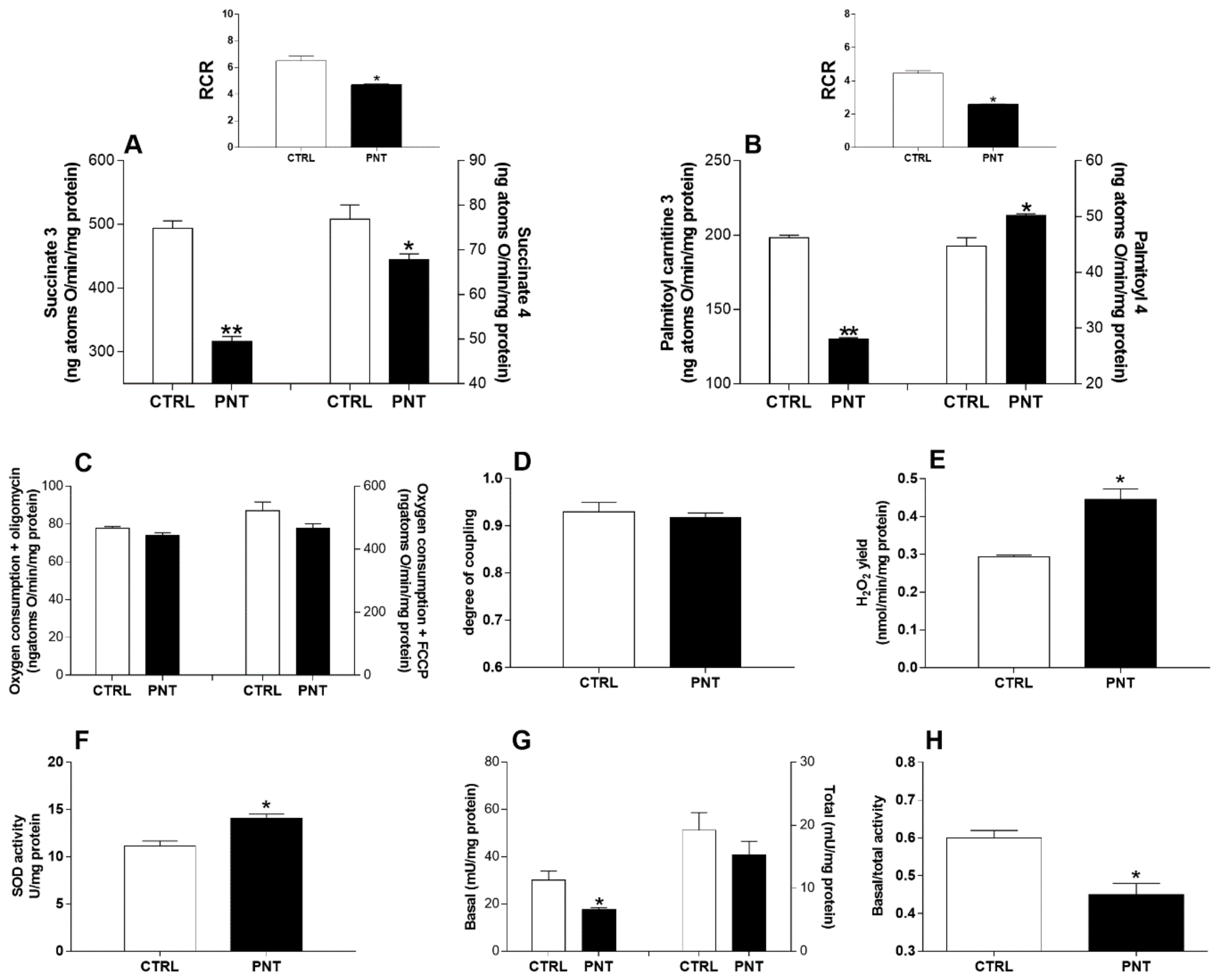
3.2. Hepatic Mitochondrial Oxidative Capacity
3.3. Hepatic Oxidative Stress
3.4. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Prescott, S.L.; Pawankar, R.; Allen, K.J.; Campbell, D.E.; Sinn, J.K.; Fiocchi, A.; Ebisawa, M.; Sampson, H.A.; Beyer, K.; Lee, B.-W. A global survey of changing patterns of food allergy burden in children. World Allergy Organ. J. 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Di Costanzo, M.; Paparo, L.; Cosenza, L.; Di Scala, C.; Nocerino, R.; Aitoro, R.; Berni Canani, R. Food Allergies: Novel Mechanisms and Therapeutic Perspectives. Methods Mol. Biol. 2016, 1371, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Roberts, G.; Sayre, P.H. Randomized Trial of Peanut Consumption in Infants at Risk for Peanut Allergy. N. Engl. J. Med. 2015, 372, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Scurlock, A.M.; Vickery, B.P.; Hourihane, J.O.B.; Burks, A.W. Pediatric food allergy and mucosal tolerance. Mucosal Immunol. 2010, 3, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.L.; Kelly, B.; O’Neill, L.A.J. Mitochondria are the powerhouses of immunity. Nat. Immunol. 2017, 18, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H. Mitochondrial Dysfunction and Oxidative Stress in Asthma: Implications for Mitochondria-Targeted Antioxidant Therapeutics. Pharmaceuticals 2011, 4, 429–456. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Mabalirajan, U. Rejuvenating cellular respiration for optimizing respiratory function: Targeting mitochondria. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L103–L113. [Google Scholar] [CrossRef] [PubMed]
- Mabalirajan, U.; Ghosh, B. Mitochondrial dysfunction in metabolic syndrome and asthma. J. Allergy 2013, 2013, 340476. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Aguirre, L.; Bacsi, A.; Saavedra-Molina, A.; Kurosky, A.; Sur, S.; Boldogh, I. Mitochondrial dysfunction increases allergic airway inflammation. J. Immunol. 2009, 183, 5379–5387. [Google Scholar] [CrossRef] [PubMed]
- Brookes, P.S.; Yoon, Y.; Pobotham, J.L.; Anders, M.W.; Sheu, S.S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am. J. Physiol. Cell Physiol. 2004, 287, C817–C833. [Google Scholar] [CrossRef] [PubMed]
- Cloonan, S.M.; Choi, A.M. Mitochondria in lung disease. J. Clin. Investig. 2016, 126. [Google Scholar] [CrossRef] [PubMed]
- Phaniendra, A.; Jestadi, B.D.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2014, 30, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Kim, D.I.; Kim, S.H.; Lee, H.; Lee, K.S.; Cho, S.H.; Lee, Y.C. NLRP3 inflammasome activation by mitochondrial ROS in bronchial epithelial cells is required for allergic inflammation. Cell Death Dis. 2014, 5, e1498. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.D.; Cui, J.M.; Wang, Y.; Yin, L.M.; Gao, C.K.; Liu, Y.Y.; Yang, Y.Q. The early asthmatic response is associated with glycolysis, calcium binding and mitochondria activity as revealed by proteomic analysis in rats. Respir. Res. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Iyer, D.; Mishra, N.; Agrawal, A. Mitochondrial Function in Allergic Disease. Curr. Allergy Asthma Rep. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Barnig, C.; Charle, A.-L.; Poirot, A.; Meyer, A.; Clere-Jehl, R.; de Blay, F.; Geny, B. Effect of nasal allergen challenge in allergic rhinitis on mitochondrial function of peripheral blood mononuclear cells. Ann. Allergy Asthma Immunol. 2017, 118, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, M.; Shoda, T.; Natsume, O.; Nomura, I.; Narita, M.; Fukuda, A.; Sakamoto, S.; Kasahara, M.; Ohya, Y. Factors Associated with Development of Food Allergy in Young Children after Liver Transplantation: A Retrospective Analysis of 10 Years’ Experience. J. Allergy Clin. Immunol. Pract. 2017, 5, 1698–1706. [Google Scholar] [CrossRef] [PubMed]
- Ozbek, O.Y.; Ozcay, F.; Avci, Z.; Haberal, A.; Haberal, M. Food allergy after liver transplantation in children: A prospective study. Pediatr. Allergy Immunol. 2009, 20. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Ochiai, H. Food allergy after cord blood stem cell transplantation with tacrolimus therapy in two patients who developed veno-occlusive disease. Allergol. Int. 2012, 61, 497–499. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Katsukura, H.; Shirai, Y.; Yamori, M.; Chiba, T.; Kita, T.; Wakatsuki, Y. Helper CD4+ T cells for IgE response to a dietary antigen develop in the liver. J. Allergy Clin. Immunol. 2003, 111, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Aitoro, R.; Simeoli, R.; Amoroso, A.; Paparo, L.; Nocerino, R.; Pirozzi, C.; di Costanzo, M.; Meli, R.; De Caro, C.; Picariello, G.; et al. Extensively hydrolyzed casein formula alone or with L. rhamnosus GG reduces β-lactoglobulin sensitization in mice. Pediatr. Allergy Immunol. 2017, 28, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Stefka, A.T.; Feehley, T.; Tripathi, P.; Qiu, J.; McCoy, K. Commensal bacteria protect against food allergen sensitization. Proc. Natl. Acad. Sci. USA 2014, 111, 13145–13150. [Google Scholar] [CrossRef] [PubMed]
- Berin, M.C.; Shreffler, W.G. T(H)2 adjuvants: Implications for food allergy. J. Allergy Clin. Immunol. 2008, 121, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Van Wijk, F.; Hoeks, S.; Nierkens, S.; Koppelman, S.J.; van Kooten, P.; Boon, L.; Knippels, L.M.J.; Pieters, R. CTLA-4 signaling regulates the intensity of hypersensitivity responses to food antigens, but is not decisive in the induction of sensitization. J. Immunol. 2005, 174, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Van Esch, B.C.; Schouten, B. Acute allergic skin response as a new tool to evaluate the allergenicity of whey hydrolysates in a mouse model of orally induced cow’s milk allergy. Pediatr. Allergy Immunol. 2010, 21, e780–e786. [Google Scholar] [CrossRef] [PubMed]
- Schouten, B.; van Esch, B.C.; Hofman, G.A.; van Doorn, S.A.; Knol, J.; Nauta, A.J.; Garssen, J.; Willemsen, L.E.; Knippels, L.M. Cow milk allergy symptoms are reduced in mice fed dietary synbiotics during oral sensitization with whey. J. Nutr. 2009, 139, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Sun, L.; Chen, B.; Han, Y.; Pang, J.; Wu, W.; Qi, R.; Zhang, T.M. Evaluation of candidate reference genes for RT-qPCR studies in three metabolism related tissues of mice after caloric restriction. Sci. Rep. 2016, 6, 38513. [Google Scholar] [CrossRef] [PubMed]
- Hartree, E.F. Determination of protein: A modification of the Lowry method that gives a linear photometric response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef]
- Mollica, M.P.; MattaceRaso, G.; Cavaliere, G.; Trinchese, G.; De Filippo, C.; Aceto, S.; Prisco, M.; Pirozzi, C.; Di Guida, F.; Lama, A.; et al. Butyrate Regulates Liver Mitochondrial Function, Efficiency, and Dynamics in Insulin-Resistant Obese Mice. Diabetes 2017, 66, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Estabrook, R.W. Mitochondrial respiratory control and the polarographic measurement of ADP:O ratios. Methods Enzymol. 1967, 10, 41–47. [Google Scholar]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, G.; Trinchese, G.; Bergamo, P.; De Filippo, C.; Mattace Raso, G.; Gifuni, G.; Putti, R.; Moni, B.H.; Canani, R.B.; Meli, R.; et al. Polyunsaturated Fatty Acids Attenuate Diet Induced Obesity and Insulin Resistance, Modulating Mitochondrial Respiratory Uncoupling in Rat Skeletal Muscle. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Cairns, C.B.; Walther, J.; Harken, A.H.; Banerjee, A. Mitochondrial oxidative phosphorylation thermodynamic efficiencies reflect physiological organ roles. Am. J. Physiol. 1998, 274, R1376–R1383. [Google Scholar] [CrossRef] [PubMed]
- Flohè, L.; Otting, F. Superoxide Dismutase Assay. Methods Enzymol. 1984, 105, 93–104. [Google Scholar] [PubMed]
- Hausladen, A.; Fridovich, I. Measuring nitric oxide and superoxide: Rate constants for aconitase reactivity. Methods Enzymol. 1996, 269, 37–41. [Google Scholar] [PubMed]
- Barja, G. Mitochondrial free radical production and aging in mammals and birds. Ann. N. Y. Acad. Sci. 1998, 854, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Shaughnessy, D.T.; McAllister, K.; Worth, L.; Haugen, A.C.; Meyer, J.N.; Domann, F.E.; Van Houten, B.; Mostoslavsky, R.; Bultman, S.J.; Baccarelli, A.A.; et al. Mitochondria, energetics, epigenetics, and cellular responses to stress. Environ. Health Perspect. 2014, 122, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Raby, B.A.; Klanderman, B.; Murphy, A.; Mazza, S.; Camargo, C.A., Jr.; Silverman, E.K.; Weiss, S.T. A common mitochondrial haplogroup is associated with elevated total serum IgE levels. J. Allergy Clin. Immunol. 2007, 120, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, A.; Zaccagnino, P.; Di Paola, M.; Saltarella, M.; Oliveros Celis, C.; Nico, B.; Santoro, G.; Lorusso, M. Role of mitochondria and reactive oxygen species in dendritic cell differentiation and functions. Free Radic. Biol. Med. 2008, 44, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.F.; Zhang, M.S.; Hu, A.H.; Zhu, Z.; Yin, K.S. Selective blockade of NF-kappa B by novel mutated IkappaBalpha suppressesCD3/CD28-induced activation of memory CD4+ T cells in asthma. Allergy 2008, 63, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Bulua, A.C.; Simon, A.; Maddipati, R.; Pelletier, M.; Park, H.; Kim, K.Y.; Sack, M.N.; Kastner, D.L.; Siegel, R.M. Mitochondrial reactive oxygen species promote production of proinflammatory cytokines and are elevated inTNFR1-associated periodic syndrome (TRAPS). J. Exp. Med. 2011, 208, 519–533. [Google Scholar] [CrossRef] [PubMed]
- Sebag, S.C.; Koval, O.M.; Paschke, J.D.; Winters, C.J.; Jaffer, O.A.; Dworski, R.; Sutterwala, F.S.; Anderson, M.E.; Grumbach, I.M. Mitochondrial CaMKII inhibition in airway epithelium protects against allergic asthma. JCI Insight 2017, 2, e88297. [Google Scholar] [CrossRef] [PubMed]
- Biltagi, M.A.; Baset, A.A.; Bassiouny, M.; Kasrawi, M.A.; Attia, M. Omega-3 fatty acids, vitamin C and Zn supplementation in asthmatic children: A randomized self-controlled study. Acta Paediatr. 2009, 98, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Leone, L.; D’Auria, E.; Riva, E.; Nocerino, R.; Ruotolo, S.; Terrin, G.; Cosenza, L.; Di Costanzo, M.; Passariello, A.; et al. The effects of dietary counseling on children with food allergy: A prospective, multicenter intervention study. J. Acad. Nutr. Diet. 2014, 114, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Aldámiz-Echevarría, L.; Bilbao, A.; Andrade, F.; Elorz, J.; Prieto, J.A.; Rodríguez-Soriano, J. Fatty acid deficiency profile in children with food allergy managed with elimination diets. Acta Paediatr. 2008, 97, 1572–1576. [Google Scholar] [CrossRef] [PubMed]
- Van den Elsen, L.; Garssen, J. Long chain N-3 polyunsaturated fatty acids in the prevention of allergic and cardiovascular disease. Curr. Pharm. Des. 2012, 18, 2375–2392. [Google Scholar] [CrossRef] [PubMed]
- Weise, C.; Hilt, K.; Milovanovic, M.; Ernst, D.; Rühl, R.; Worm, M. Inhibition of IgE production by docosahexaenoic acid is mediated by direct interference with STAT6 and NFκB pathway in human B cells. J. Nutr. Biochem. 2011, 22, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Trinchese, G.; Cavaliere, G.; Canani, R.B.; Matamoros, S.; Bergamo, P.; De Filippo, C.; Aceto, S.; Gaita, M.; Cerino, P.; Negri, R.; et al. Human, donkey and cow milk differently affects energy efficiency and inflammatory state by modulating mitochondrial function and gut microbiota. J. Nutr. Biochem. 2015, 26, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Mollica, M.P.; Trinchese, G.; Cavaliere, G.; De Filippo, C.; Cocca, E.; Gaita, M.; Della-Gatta, A.; Marano, A.; Mazzarella, G.; Bergamo, P. c9, t11-Conjugated linoleic acid ameliorates steatosis by modulating mitochondrial uncoupling and Nrf2 pathway. J. Lipid Res. 2014, 55, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, B.; Bodas, M.; Bhatraju, N.K.; Ahmad, T.; Pant, R.; Guleria, R.; Ghosh, B.; Agrawal, A. IL-4 promotes asymmetric dimethylarginine accumulation, oxo-nitrative stress, and hypoxic response-induced mitochondrial loss in airway epithelial cells. J. Allergy Clin. Immunol. 2016, 138, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Bogdanos, D.P.; Gao, B. Liver immunology. Compr. Physiol. 2013, 3, 567–598. [Google Scholar] [CrossRef] [PubMed]
- Knolle, P.A.; Wohlleber, D. Immunological functions of liver sinusoidal endothelial cells. Cell. Mol. Immunol. 2016, 13, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.W.; Knolle, P.A. Antigen-presenting cell function in the tolerogenic liver environment. Nat. Rev. Immunol. 2010, 10, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, J.P.; Knolle, P.A.; Stabenow, D. Mechanisms balancing tolerance and immunity in the liver. Dig. Dis. 2011, 29, 384–390. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | RefSeq | Exon Boundary | Assay Location | Amplicon Length |
|---|---|---|---|---|
| IL5 | NM_010558.1 | 2–3 | 219 | 62 |
| IL4 | NM_021283.2 | 2–3 | 241 | 79 |
| IL13 | NM_008355.3 | 1–2 | 211 | 56 |
| IL-10 | NM_010548.2 | 4–5 | 515 | 136 |
| IFN-γ | NM_008337.3 | 3–4 | 469 | 100 |
| GUSb | NM_010368.1 | 10–11 | 1657 | 71 |
| Palmitoyl-Carnitine 3 | H2O2 | Basal/Total Aconitase | Anti-PNT IgE | IL-4 | IL-5 | IL-13 | ||
|---|---|---|---|---|---|---|---|---|
| Succinate 3 | Pearson Correlation | 0.951 ** | −0.864 ** | 0.778 ** | −0.950 ** | −0.588 * | −0.854 ** | −0.742 ** |
| Sig. (2-tailed) | 0.000 | 0.000 | 0.003 | 0.000 | 0.044 | 0.000 | 0.006 | |
| Palmitoyl-carnitine 3 | Pearson Correlation | – | −0.873 ** | 0.786 ** | −0.966 ** | −0.683 * | −0.919 ** | −0.832 ** |
| Sig. (2-tailed) | 0.000 | 0.002 | 0.000 | 0.014 | 0.000 | 0.001 | ||
| H2O2 | Pearson Correlation | – | −0.848 ** | 0.0875 ** | 0.651 * | 0.750 ** | 0.802 ** | |
| Sig. (2-tailed) | 0 | 0.000 | 0.022 | 0.005 | 0.002 | |||
| Basal/total aconitase | Pearson Correlation | – | −0.875** | −0.376 | −0.768 ** | −0.642 * | ||
| Sig. (2-tailed) | 0.000 | 0.228 | 0.004 | 0.024 | ||||
| Anti-PNT IgE | Pearson Correlation | – | 0.525 | 0.931 ** | 0.757 ** | |||
| Sig. (2-tailed) | 0.079 | 0.000 | 0.004 | |||||
| IL-4 | Pearson Correlation | – | 0.600 * | 0.849 ** | ||||
| Sig. (2-tailed) | 0.039 | 0.000 | ||||||
| IL-5 | Pearson Correlation | – | 0.842 ** | |||||
| Sig. (2-tailed) | 0.001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trinchese, G.; Paparo, L.; Aitoro, R.; Fierro, C.; Varchetta, M.; Nocerino, R.; Mollica, M.P.; Berni Canani, R. Hepatic Mitochondrial Dysfunction and Immune Response in a Murine Model of Peanut Allergy. Nutrients 2018, 10, 744. https://doi.org/10.3390/nu10060744
Trinchese G, Paparo L, Aitoro R, Fierro C, Varchetta M, Nocerino R, Mollica MP, Berni Canani R. Hepatic Mitochondrial Dysfunction and Immune Response in a Murine Model of Peanut Allergy. Nutrients. 2018; 10(6):744. https://doi.org/10.3390/nu10060744
Chicago/Turabian StyleTrinchese, Giovanna, Lorella Paparo, Rosita Aitoro, Carmela Fierro, Michela Varchetta, Rita Nocerino, Maria Pina Mollica, and Roberto Berni Canani. 2018. "Hepatic Mitochondrial Dysfunction and Immune Response in a Murine Model of Peanut Allergy" Nutrients 10, no. 6: 744. https://doi.org/10.3390/nu10060744
APA StyleTrinchese, G., Paparo, L., Aitoro, R., Fierro, C., Varchetta, M., Nocerino, R., Mollica, M. P., & Berni Canani, R. (2018). Hepatic Mitochondrial Dysfunction and Immune Response in a Murine Model of Peanut Allergy. Nutrients, 10(6), 744. https://doi.org/10.3390/nu10060744