Intermittent Fasting with or without Exercise Prevents Weight Gain and Improves Lipids in Diet-Induced Obese Mice

 and
and 
Abstract
1. Introduction
2. Materials and Methods
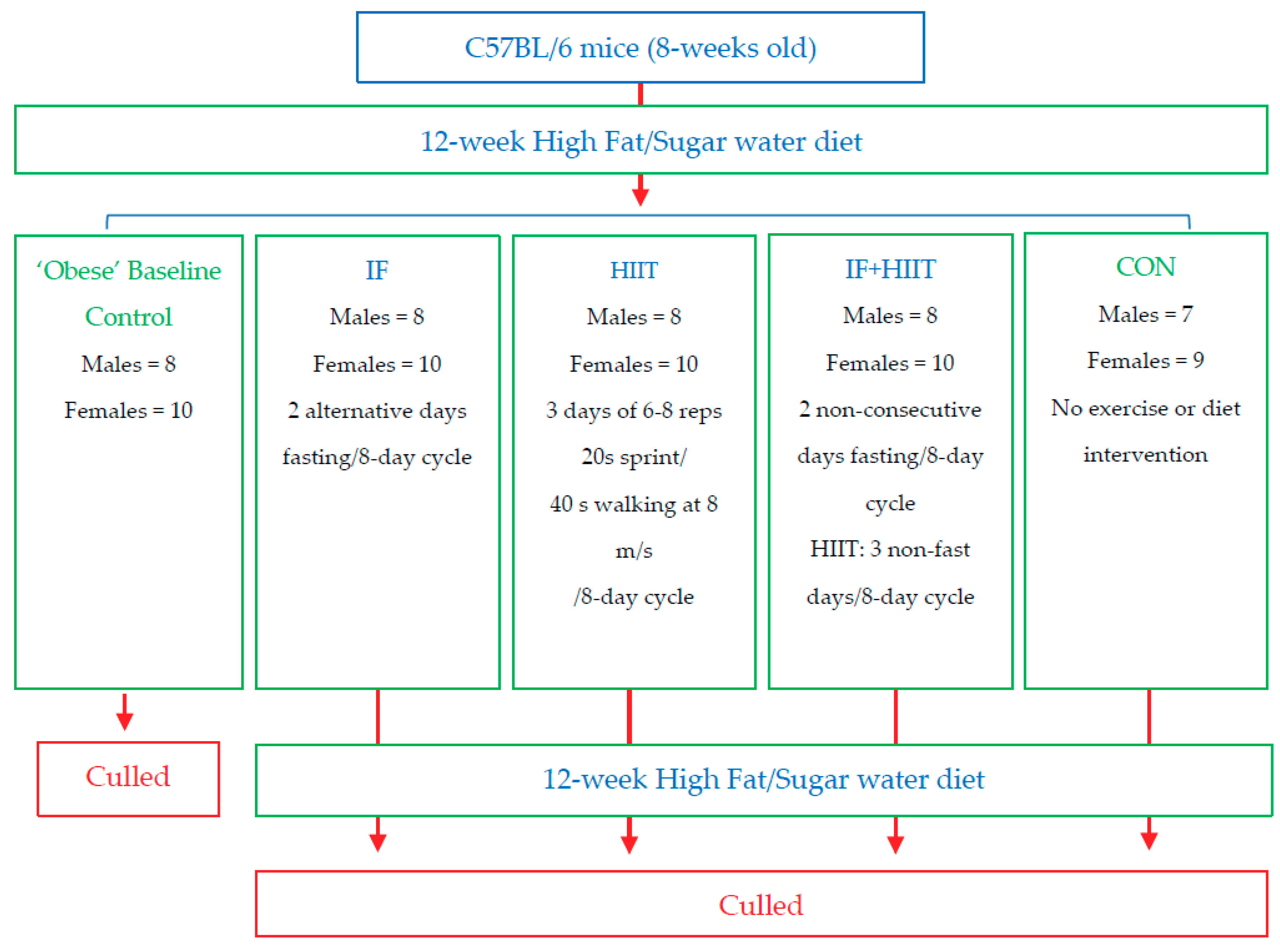
2.1. Animals
2.2. Intermittent Fasting Protocol
2.3. Top Speed Test and High Intensity Interval Training Protocol
2.4. Body Weight and Composition Assessment
2.5. Assessment of Fasting Blood Glucose and Glucose Tolerance (AUC)
2.6. Assessment of Plasma Insulin, Glucose and Triacylglycerol (TAG), Low-Density Lipoprotein (LDL) and High-Density Lipoprotein (HDL) Levels
2.7. Assessment of Homeostatic Model Assessment of Insulin Resistance (HOMA-IR)
2.8. Assessment of Muscle Strength
2.9. Assessment of Caloric Intake
2.10. Statistical Analysis
3. Results
3.1. Effects of 12-Weeks of the HF/S Diet on Body Weight of All Animals
3.2. Effects of IF on Caloric Intake
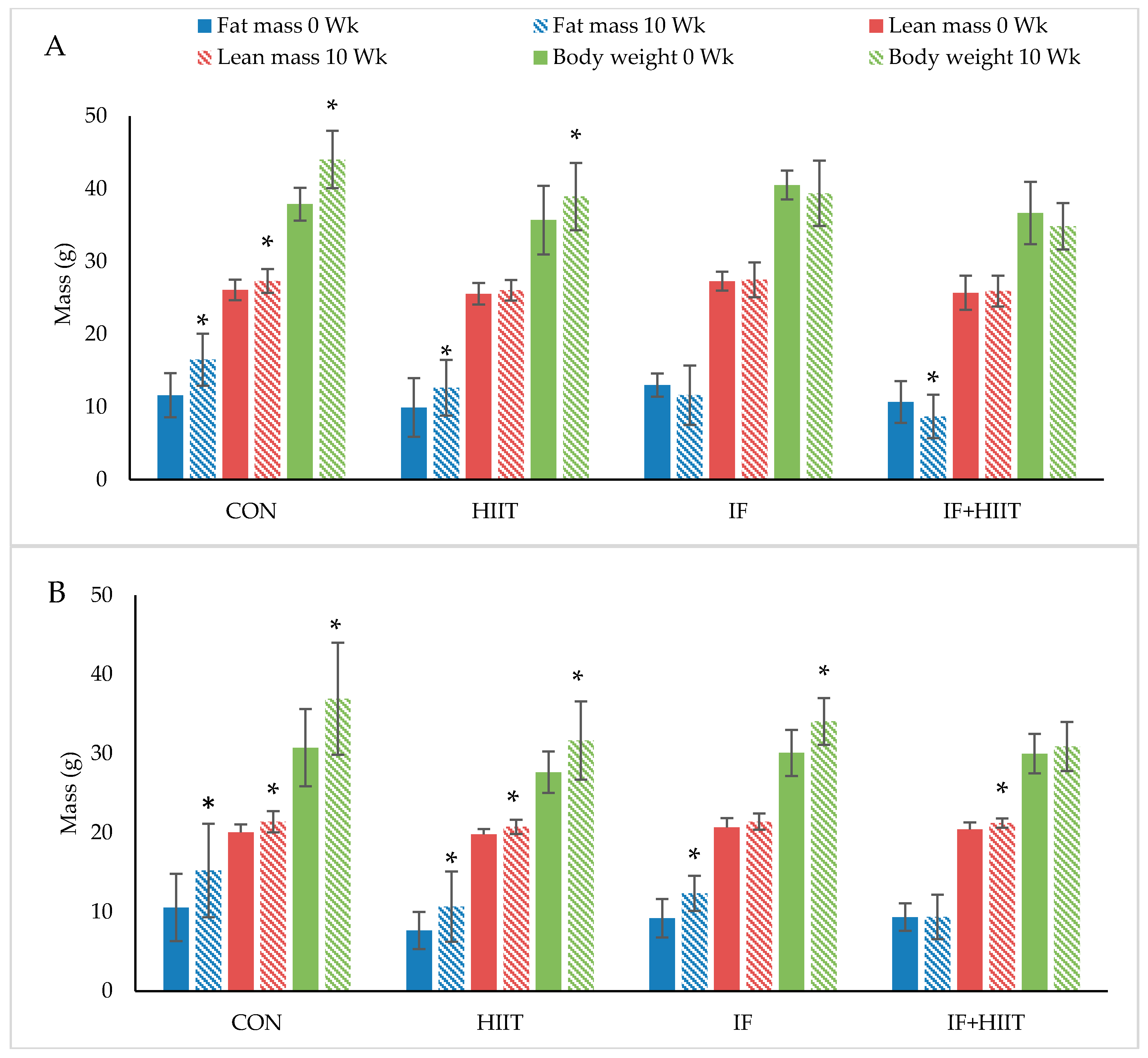
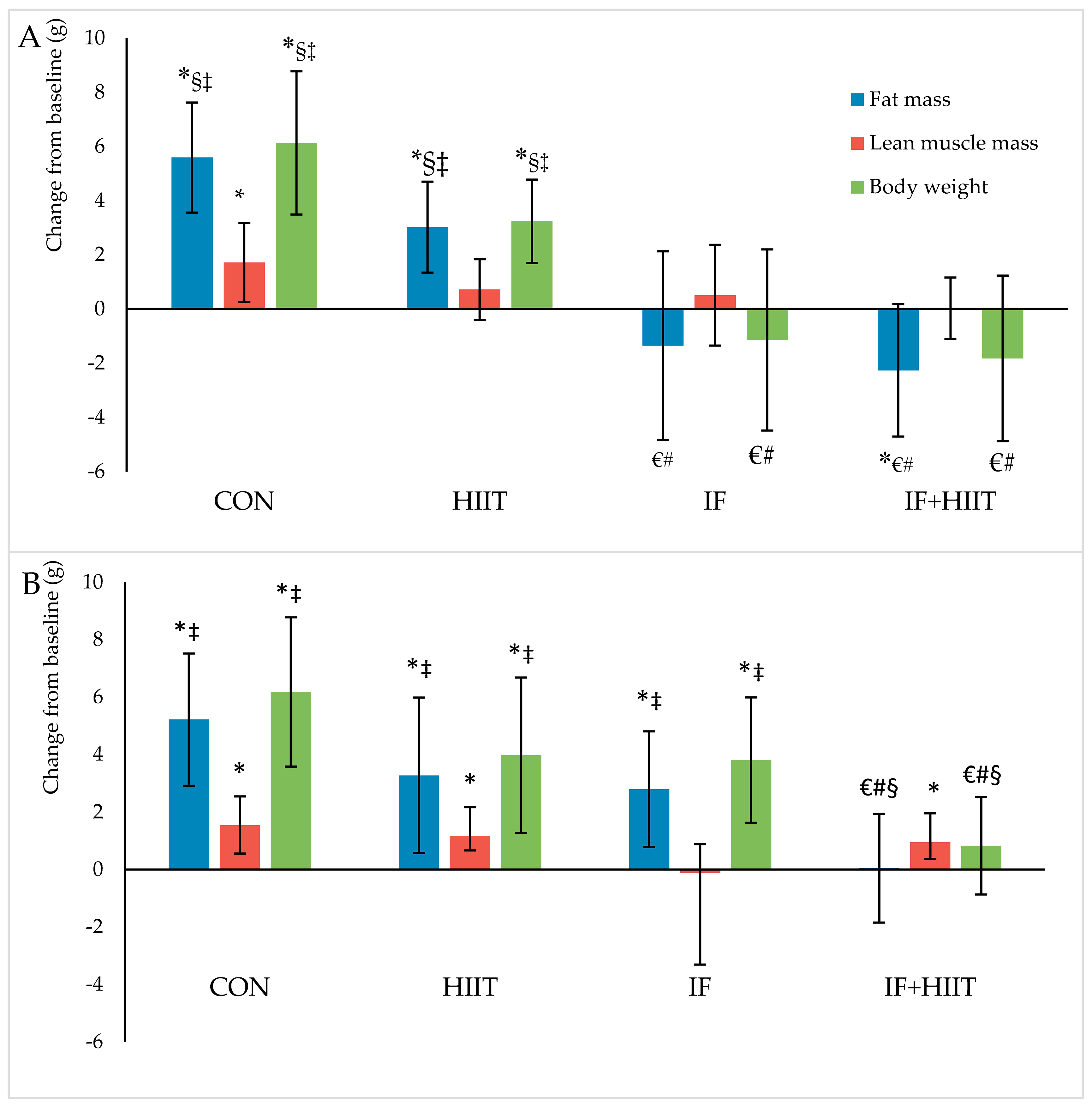
3.3. Effects of IF and/or HIIT on Body Weight and Composition
3.3.1. Body Weight and Fat Mass
3.3.2. Lean Mass
3.4. Effect of IF and/or HIIT on Blood Glucose, Plasma Insulin, HOMA-IR and AUC
3.4.1. Blood Glucose
3.4.2. Plasma Insulin
3.4.3. Insulin Resistance (HOMA-IR) and Glucose Tolerance (AUC)
3.5. Effect of IF and/or HIIT on Lipid Profiles
3.6. Effect of IF and/or HIIT on Muscle Strength
4. Discussion
5. Conclusions
6. Limitations
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Van Vliet-Ostaptchouk, J.V.; Snieder, H.; Lagou, V. Gene–lifestyle interactions in obesity. Curr. Nutr. Rep. 2012, 1, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Romieu, I.; Dossus, L.; Barquera, S.; Blottiere, H.M.; Franks, P.W.; Gunter, M.; Hwalla, N.; Hursting, S.D.; Leitzmann, M.; Margetts, B.; et al. Energy balance and obesity: What are the main drivers? Cancer Causes Control 2017, 28, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Spreitzer, A.; Haber, P. Fat loss depends on energy deficit only, independently of the method for weight loss. Ann. Nutr. Metabol. 2007, 51, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Türk, Y.; Theel, W.; Kasteleyn, M.J.; Franssen, F.M.E.; Hiemstra, P.S.; Rudolphus, A.; Taube, C.; Braunstahl, G.J. High intensity training in obesity: A meta-analysis. Obes. Sci. Pract. 2017, 3, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Tinsley, G.M.; La Bounty, P.M. Effects of intermittent fasting on body composition and clinical health markers in humans. Nutr. Rev. 2015, 73, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Barnosky, A.R.; Hoddy, K.K.; Unterman, T.G.; Varady, K.A. Intermittent fasting vs daily calorie restriction for type 2 diabetes prevention: A review of human findings. Transl. Res. J. Lab. Clin. Med. 2014, 164, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Heilbronn, L.K.; Smith, S.R.; Martin, C.K.; Anton, S.D.; Ravussin, E. Alternate-day fasting in nonobese subjects: Effects on body weight, body composition, and energy metabolism. Am. J. Clin. Nutr. 2005, 81, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, F.R.D.; Ikeoka, D.; Caramelli, B. Effects of intermittent fasting on metabolism in men. Rev. Assoc. Med. Bras. 2013, 59, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Seimon, R.V.; Roekenes, J.A.; Zibellini, J.; Zhu, B.; Gibson, A.A.; Hills, A.P.; Wood, R.E.; King, N.A.; Byrne, N.M.; Sainsbury, A. Do intermittent diets provide physiological benefits over continuous diets for weight loss? A systematic review of clinical trials. Mol. Cell. Endocrinol. 2015, 418 Pt 2, 153–172. [Google Scholar] [CrossRef] [PubMed]
- Gibala, M.J.; Little, J.P.; Macdonald, M.J.; Hawley, J.A. Physiological adaptations to low-volume, high-intensity interval training in health and disease. J. Physiol. 2012, 590, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Trapp, E.G.; Chisholm, D.J.; Freund, J.; Boutcher, S.H. The effects of high-intensity intermittent exercise training on fat loss and fasting insulin levels of young women. Int. J. Obes. 2008, 32, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Gerber, T.; Borg, M.L.; Hayes, A.; Stathis, C.G. High-intensity intermittent cycling increases purine loss compared with workload-matched continuous moderate intensity cycling. Eur. J. Appl. Physiol. 2014, 114, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Racil, G.; Ben Ounis, O.; Hammouda, O.; Kallel, A.; Zouhal, H.; Chamari, K.; Amri, M. Effects of high vs. Moderate exercise intensity during interval training on lipids and adiponectin levels in obese young females. Eur. J. Appl. Physiol. 2013, 113, 2531–2540. [Google Scholar] [CrossRef] [PubMed]
- Earnest, C.P.; Lupo, M.; Thibodaux, J.; Hollier, C.; Butitta, B.; Lejeune, E.; Johannsen, N.M.; Gibala, M.J.; Church, T.S. Interval training in men at risk for insulin resistance. Int. J. Sports Med. 2013, 34, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Gillen, J.B.; Percival, M.E.; Ludzki, A.; Tarnopolsky, M.A.; Gibala, M.J. Interval training in the fed or fasted state improves body composition and muscle oxidative capacity in overweight women. Obesity (Silver Spring Md.) 2013, 21, 2249–2255. [Google Scholar] [CrossRef] [PubMed]
- Sartor, F.; de Morree, H.M.; Matschke, V.; Marcora, S.M.; Milousis, A.; Thom, J.M.; Kubis, H.P. High-intensity exercise and carbohydrate-reduced energy-restricted diet in obese individuals. Eur. J. Appl. Physiol. 2010, 110, 893–903. [Google Scholar] [CrossRef] [PubMed]
- La Fleur, S.E.; Luijendijk, M.C.; van Rozen, A.J.; Kalsbeek, A.; Adan, R.A. A free-choice high-fat high-sugar diet induces glucose intolerance and insulin unresponsiveness to a glucose load not explained by obesity. Int. J. Obes. (Lond.) 2011, 35, 595–604. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Joslin, P.M.N.; Bell, R.K.; Swoap, S.J. Obese mice on a high-fat alternate-day fasting regimen lose weight and improve glucose tolerance. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Castello, L.; Froio, T.; Maina, M.; Cavallini, G.; Biasi, F.; Leonarduzzi, G.; Donati, A.; Bergamini, E.; Poli, G.; Chiarpotto, E. Alternate-day fasting protects the rat heart against age-induced inflammation and fibrosis by inhibiting oxidative damage and NF-κB activation. Free Radic. Biol. Med. 2010, 48, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Varady, K.A.; Roohk, D.J.; Loe, Y.C.; McEvoy-Hein, B.K.; Hellerstein, M.K. Effects of modified alternate-day fasting regimens on adipocyte size, triglyceride metabolism, and plasma adiponectin levels in mice. J. Lipid Res. 2007, 48, 2212–2219. [Google Scholar] [CrossRef] [PubMed]
- Tuazon, M.A.; McConnell, T.R.; Wilson, G.J.; Anthony, T.G.; Henderson, G.C. Intensity-dependent and sex-specific alterations in hepatic triglyceride metabolism in mice following acute exercise. J. Appl. Physiol. (Bethesda Md. 1985) 2015, 118, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Lund, J.; Hafstad, A.D.; Boardman, N.T.; Rossvoll, L.; Rolim, N.P.; Ahmed, M.S.; Florholmen, G.; Attramadal, H.; Wisloff, U.; Larsen, T.S.; et al. Exercise training promotes cardioprotection through oxygen-sparing action in high-fat fed mice. Am. J. Physiol. Heart Circ. Physiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Andrikopoulos, S.; Blair, A.R.; Deluca, N.; Fam, B.C.; Proietto, J. Evaluating the glucose tolerance test in mice. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1323–E1332. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.J. Measuring the strength of mice. J. Vis. Exp. JoVE 2013. [Google Scholar] [CrossRef] [PubMed]
- Higashida, K.; Fujimoto, E.; Higuchi, M.; Terada, S. Effects of alternate-day fasting on high-fat diet-induced insulin resistance in rat skeletal muscle. Life Sci. 2013, 93, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Gotthardt, J.D.; Verpeut, J.L.; Yeomans, B.L.; Yang, J.A.; Yasrebi, A.; Roepke, T.A.; Bello, N.T. Intermittent fasting promotes fat loss with lean mass retention, increased hypothalamic norepinephrine content, and increased neuropeptide y gene expression in diet-induced obese male mice. Endocrinology 2016, 157, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.A.H.; Halbrooks, J.E.; Watkins, E.E.; Fisher, G.; Hunter, G.R.; Nagy, T.R.; Plaisance, E.P. High-intensity interval training and calorie restriction promote remodeling of glucose and lipid metabolism in diet-induced obesity. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E243–E256. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Li, S.; Shao, M.; Qi, Q.; Zhao, F.; You, J.; Mao, T.; Li, W.; Yan, Z.; Liu, Y. Calorie restriction and endurance exercise share potent anti-inflammatory function in adipose tissues in ameliorating diet-induced obesity and insulin resistance in mice. Nutr. Metab. 2010, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Huffman, D.M.; Moellering, D.R.; Grizzle, W.E.; Stockard, C.R.; Johnson, M.S.; Nagy, T.R. Effect of exercise and calorie restriction on biomarkers of aging in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1618–R1627. [Google Scholar] [CrossRef] [PubMed]
- Karbowska, J.; Kochan, Z. Intermittent fasting up-regulates Fsp27/Cidec gene expression in white adipose tissue. Nutrition 2012, 28, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Byrne, N.M.; Sainsbury, A.; King, N.A.; Hills, A.P.; Wood, R.E. Intermittent energy restriction improves weight loss efficiency in obese men: The MATADOR study. Int. J. Obes. (Lond.) 2018, 42, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Keating, S.E.; Johnson, N.A.; Mielke, G.I.; Coombes, J.S. A systematic review and meta-analysis of interval training versus moderate-intensity continuous training on body adiposity. Obes. Rev. 2017, 18, 943–964. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Kim, S.; Lee, S.; Kang, H. Effect of training intensity on nonalcoholic fatty liver disease. Med. Sci. Sports Exerc. 2015, 47, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Hallahan, N.L.; Brown, S.H.; Liu, M.; Mitchell, T.W.; Cooney, G.J.; Turner, N. Mouse strain-dependent variation in obesity and glucose homeostasis in response to high-fat feeding. Diabetologia 2013, 56, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, F.M.; da Cunha, F.M.; Caldeira da Silva, C.C.; Chausse, B.; Romano, R.L.; Garcia, C.C.; Colepicolo, P.; Medeiros, M.H.; Kowaltowski, A.J. Long-term intermittent feeding, but not caloric restriction, leads to redox imbalance, insulin receptor nitration, and glucose intolerance. Free Radic. Biol. Med. 2011, 51, 1454–1460. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V. Rapamycin-induced glucose intolerance: Hunger or starvation diabetes. Cell. Cycle 2011, 10, 4217–4224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-N.; Mitchell, S.E.; Hambly, C.; Morgan, D.G.; Clapham, J.C.; Speakman, J.R. Physiological and behavioral responses to intermittent starvation in C57BL/6J mice. Physiol. Behav. 2012, 105, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Trepanowski, J.F.; Kroeger, C.M.; Barnosky, A.; Klempel, M.C.; Bhutani, S.; Hoddy, K.K.; Gabel, K.; Freels, S.; Rigdon, J.; Rood, J.; et al. Effect of alternate-day fasting on weight loss, weight maintenance, and cardioprotection among metabolically healthy obese adults: A randomized clinical trial. JAMA Intern. Med. 2017, 177, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Wing, R.R.; Lang, W.; Wadden, T.A.; Safford, M.; Knowler, W.C.; Bertoni, A.G.; Hill, J.O.; Brancati, F.L.; Peters, A.; Wagenknecht, L.; et al. Benefits of modest weight loss in improving cardiovascular risk factors in overweight and obese individuals with type 2 diabetes. Diabetes Care 2011, 34, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Burcelin, R.; Crivelli, V.; Dacosta, A.; Roy-Tirelli, A.; Thorens, B. Heterogeneous metabolic adaptation of C57BL/6J mice to high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E834–E842. [Google Scholar] [CrossRef] [PubMed]
- Muhlhausler, B.S.; Bloomfield, F.H.; Gillman, M.W. Whole animal experiments should be more like human randomized controlled trials. PLoS Biol. 2013, 11, e1001481. [Google Scholar] [CrossRef] [PubMed]
- Gillen, J.B.; Martin, B.J.; MacInnis, M.J.; Skelly, L.E.; Tarnopolsky, M.A.; Gibala, M.J. Twelve weeks of sprint interval training improves indices of cardiometabolic health similar to traditional endurance training despite a five-fold lower exercise volume and time commitment. PLoS ONE 2016, 11, e0154075. [Google Scholar] [CrossRef] [PubMed]
- Varady, K.A.; Bhutani, S.; Church, E.C.; Klempel, M.C. Short-term modified alternate-day fasting: A novel dietary strategy for weight loss and cardioprotection in obese adults. Am. J. Clin. Nutr. 2009, 90, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Varady, K.A.; Bhutani, S.; Klempel, M.C.; Kroeger, C.M.; Trepanowski, J.F.; Haus, J.M.; Hoddy, K.K.; Calvo, Y. Alternate day fasting for weight loss in normal weight and overweight subjects: A randomized controlled trial. Nutr. J. 2013, 12, 146. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Gender | CON | HIIT | IF | IF + HIIT | ||||
|---|---|---|---|---|---|---|---|---|---|
| 0 Week | 10 Week | 0 Week | 10 Week | 0 Week | 10 Week | 0 Week | 10 Week | ||
| Fasting blood glucose (mmol/L) | Males | 7.2 ± 0.9 | 8.8 ± 1.7 * | 7.3 ± 1.4 | 7.8 ± 1.1 | 7.6 ± 0.9 | 9.3 ± 2.1 * | 7.3 ± 0.8 | 7.3 ± 1.3 |
| Females | 6.9 ± 1.6 | 8.1 ± 2.3 | 6.8 ± 0.6 | 6.5 ± 1.2 | 7.3 ± 1.0 | 10.6± 0.8 *,€,#,‡ | 6.6 ± 0.7 | 6.4 ± 1.0 | |
| Glucose AUC | Males | 2772 ± 492 | 2785 ± 446 | 2490 ± 377 | 2608 ± 309 | 2308 ± 231 | 2519 ± 512 | 2048 ± 325 | 1917 ± 219 |
| Females | 1756 ± 361 | 2079 ± 553 | 1666 ± 391 | 1851 ± 546 | 1648 ± 267 | 2270 ± 96 * | 1387 ± 190 | 1882 ± 349 * | |
| Variables | Males | Females | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| OBC | CON | HIIT | IF | IF + HIIT | OBC | CON | HIIT | IF | IF + HIIT | |
| TAG (mg/dL) | 60.1 ± 14.6 | 58.9 ± 10.7 | 72.1 ± 16.5 | 62.7 ± 10.6 | 57.0 ± 27.7 | 65.9 ± 20.5 | 70.9 ± 13.1 | 75.6 ± 14.8 ‡ | 66.5 ± 15.7 | 51.3 ± 24.7 # |
| HDL (mg/dL) | 69.8 ± 12.2 | 76.2 ± 11.6 | 68.7 ± 11.3 | 82.1 ± 32.1 | 72.1 ± 17.3 | 51.1 ± 9.0 | 65.1 ± 23.0 | 61.1 ± 9.5 * | 58.6 ± 9.1 * | 50.8 ± 7.7 |
| LDL (mg/dL) | 40.5 ± 17.7 | 82.3 ± 36.0 *,‡,§ | 74.8 ± 15.6 *,‡,§ | 40.4 ± 16.9 €,# | 29.9 ± 8.9 *,€,# | 40.5 ± 17.7 | 24.8 ± 11.8 * | 17.8 ± 4.8 * | 19.5 ± 6.0 * | 27.0 ± 8.9 * |
| Variables | Gender | CON | HIIT | IF | IF + HIIT | ||||
|---|---|---|---|---|---|---|---|---|---|
| 0 Week | 10 Week | 0 Week | 10 Week | 0 Week | 10 Week | 0 Week | 10 Week | ||
| Absolute muscle strength (Score) # | Males | 9.4 ± 1.8 | 10.5 ± 1.7 * | 10.1 ± 1.2 | 10.6 ± 1.4 | 10.7 ± 1.6 | 11.2 ± 0.9 | 10.5 ± 1.1 | 11.1 ± 1.3 |
| Females | 9.7 ± 2.7 | 9.8 ± 2.6 | 10.8 ± 1.9 | 11.3 ± 2.3 | 12.1 ± 1.9 | 9.4 ± 1.6 * | 11.8 ± 1.9 | 12.2 ± 1.6 | |
| Relative muscle strength (Score/LM) $ | Males | 0.33 ± 0.05 | 0.35 ± 0.05 | 0.37 ± 0.04 | 0.38 ± 0.05 | 0.37 ± 0.05 | 0.39 ± 0.04 | 0.37 ± 0.05 | 0.39 ± 0.05 |
| Females | 0.44 ± 0.12 | 0.42 ± 0.10 | 0.50 ± 0.10 | 0.50 ± 0.11 | 0.52 ± 0.11 | 0.40 ± 0.09 * | 0.54 ± 0.09 | 0.53 ± 0.08 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, R.A.; Deasy, W.; Stathis, C.G.; Hayes, A.; Cooke, M.B. Intermittent Fasting with or without Exercise Prevents Weight Gain and Improves Lipids in Diet-Induced Obese Mice. Nutrients 2018, 10, 346. https://doi.org/10.3390/nu10030346
Wilson RA, Deasy W, Stathis CG, Hayes A, Cooke MB. Intermittent Fasting with or without Exercise Prevents Weight Gain and Improves Lipids in Diet-Induced Obese Mice. Nutrients. 2018; 10(3):346. https://doi.org/10.3390/nu10030346
Chicago/Turabian StyleWilson, Robin A., William Deasy, Christos G. Stathis, Alan Hayes, and Matthew B. Cooke. 2018. "Intermittent Fasting with or without Exercise Prevents Weight Gain and Improves Lipids in Diet-Induced Obese Mice" Nutrients 10, no. 3: 346. https://doi.org/10.3390/nu10030346
APA StyleWilson, R. A., Deasy, W., Stathis, C. G., Hayes, A., & Cooke, M. B. (2018). Intermittent Fasting with or without Exercise Prevents Weight Gain and Improves Lipids in Diet-Induced Obese Mice. Nutrients, 10(3), 346. https://doi.org/10.3390/nu10030346